A 2D model for hydrodynamics and biology coupling applied to algae growth simulations
Texte intégral
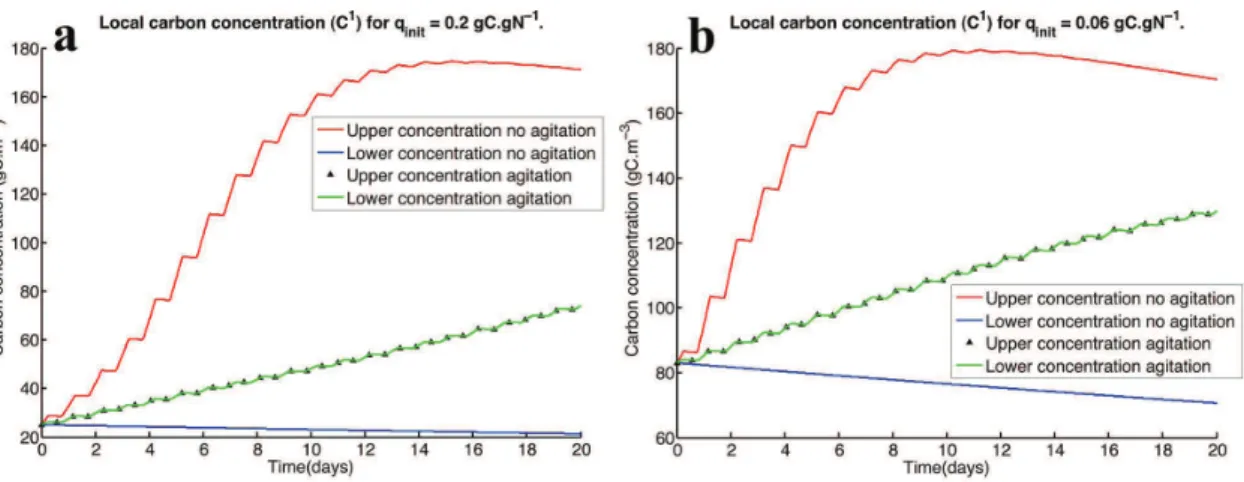
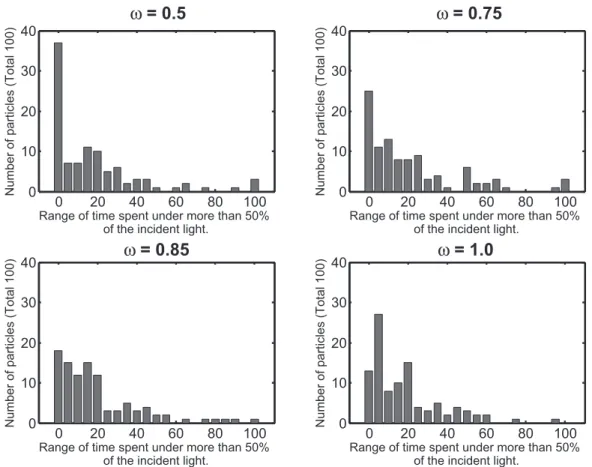
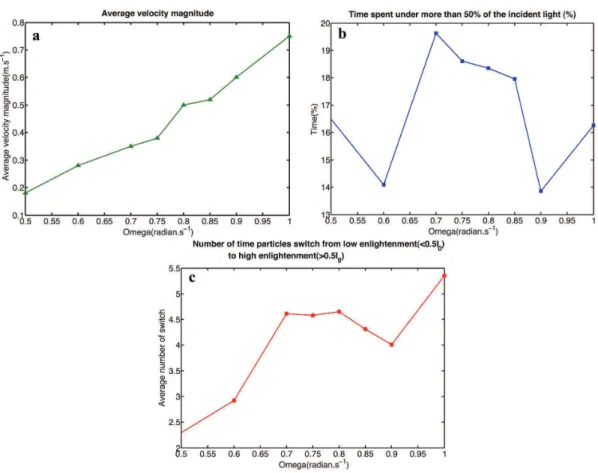
Figure



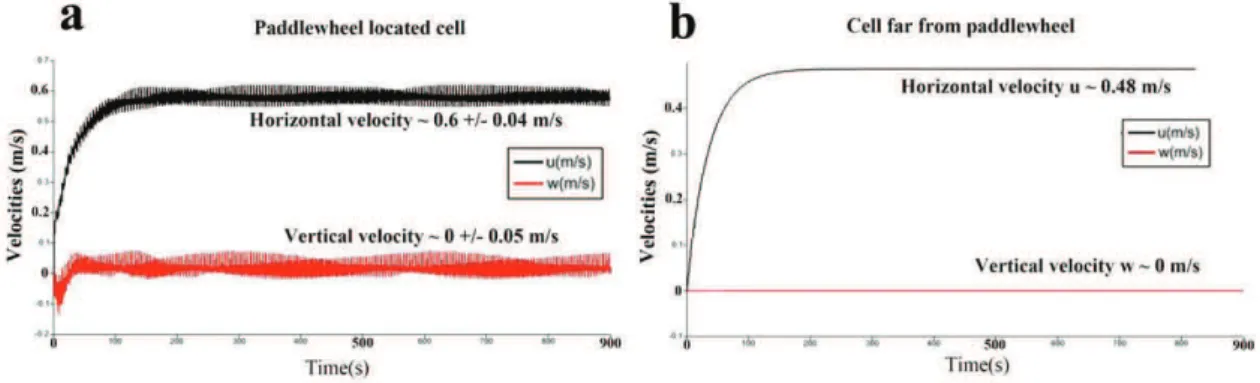
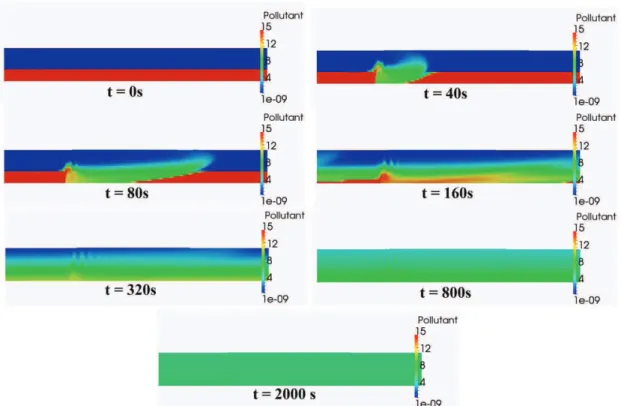
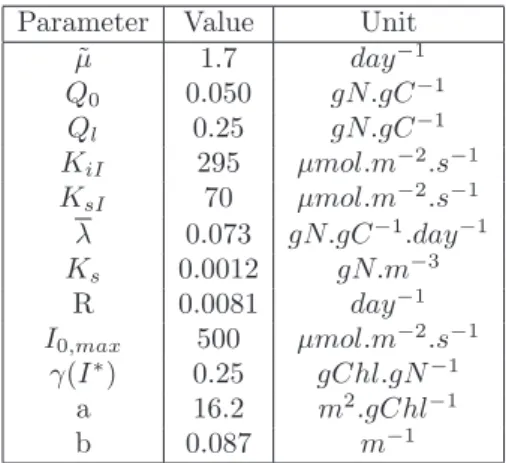
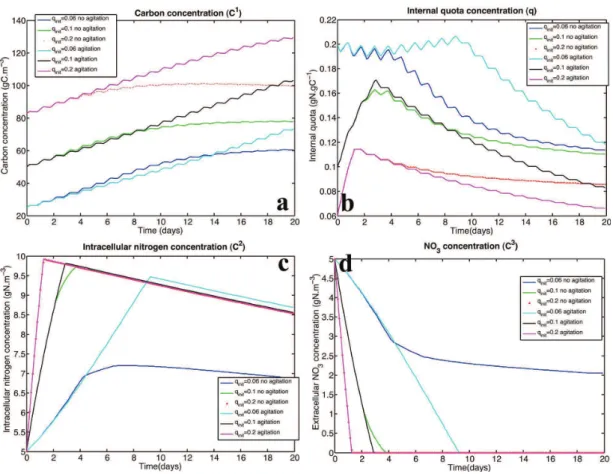
Documents relatifs
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Lorraine : LORIA, Technopôle de Nancy-Brabois
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Futurs : Parc Club Orsay Université - ZAC
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Futurs : Parc Club Orsay Université - ZAC
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Futurs : Parc Club Orsay Université - ZAC
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Lorraine : LORIA, Technopôle de Nancy-Brabois
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Futurs : Parc Club Orsay Université - ZAC
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Lorraine : LORIA, Technopôle de Nancy-Brabois
Unité de recherche INRIA Sophia Antipolis 2004, route des Lucioles - BP 93 - 06902 Sophia Antipolis Cedex France Unité de recherche INRIA Lorraine : LORIA, Technopôle de Nancy-Brabois
