Modelling plant compensatory effects in plant-insect dynamics
Texte intégral
Figure
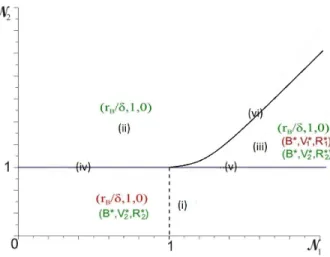
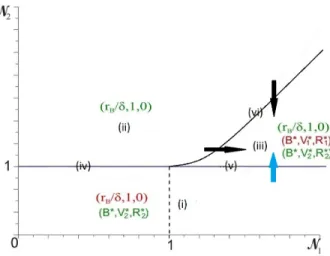
Documents relatifs
As a result, the simulation of the functional model allowed the module diameters and the total plant height of the two varieties to be recovered correctly (Fig.. We still need
The development parameters, defining the module number of phytomers and branching rules are nearly stable and close for both varieties under the various environmental
These microbial assemblages, collectively referred to as the plant microbiota, impact plant fitness through the modification of a number of traits including: biomass
Nous avons mis en évidence, dans le cadre de ce travail, que l'hétérogénéité spatiale et temporelle des paysages agricoles - et notamment la taille moyenne du
For instance, in many biological pest control models, the plant compartment is not present, even if it is well known that plants have developed many defenses strategies against
If the main objective of the control is to maintain the crop yield above a critical threshold, then plant growth and plant-insect interactions have to be taken into account.. They
However, up to now, biological control modelling has primarily focused on pests-natural enemies interactions considering somehow that crop yield is not affected by the pests..
By the Frobenius theorem the norm of the quadratic form above is bounded by the row sum of the matrix, which is bounded above by an absolute constant α 1.. Proof of Theorem
