Negative Regulation of NKG2D Expression by IL-4 in Memory CD8 T Cells
42
0
0
Texte intégral
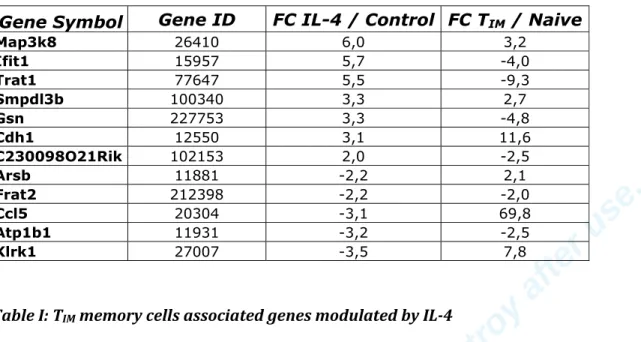
Figure
Documents relatifs
