Courtship communication in the wild chimpanzees of Budongo, Uganda
144
0
0
Texte intégral
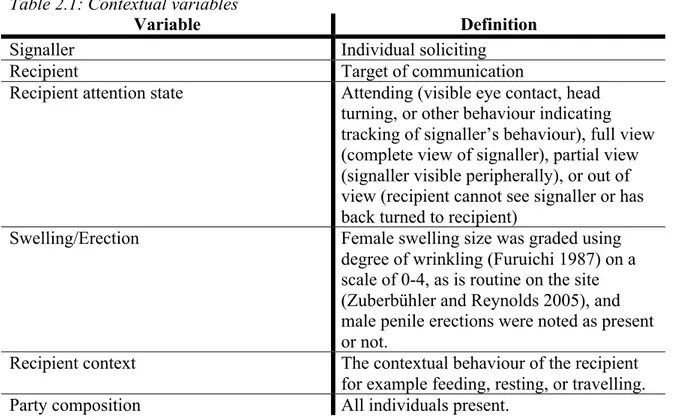
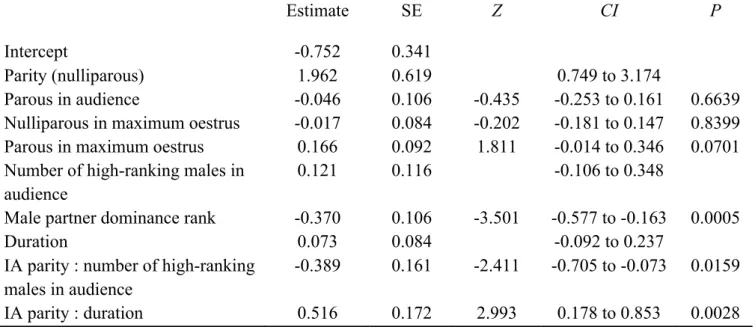
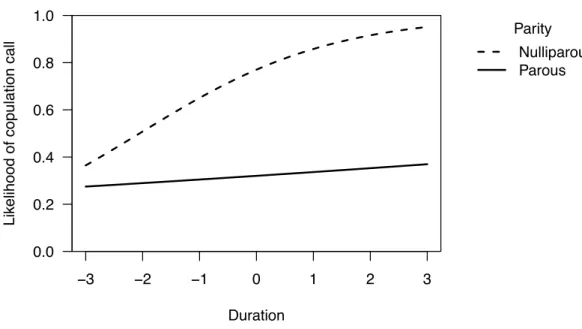
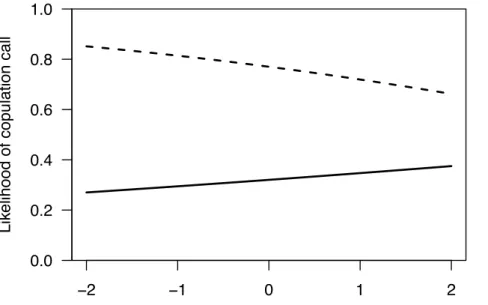
Figure
+7

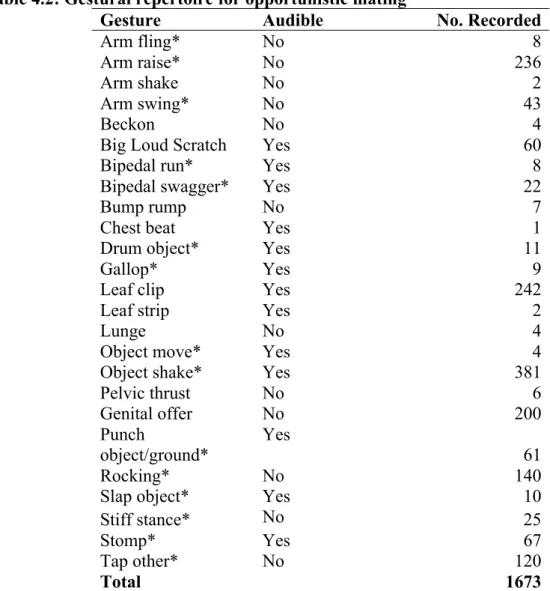

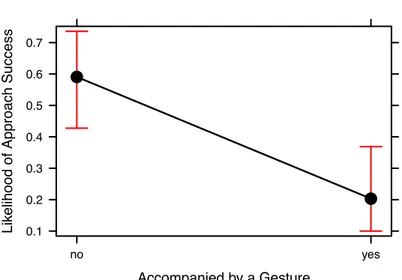

Documents relatifs
