HIV-1 Vpr—a still "enigmatic multitasker"
14
0
0
Texte intégral
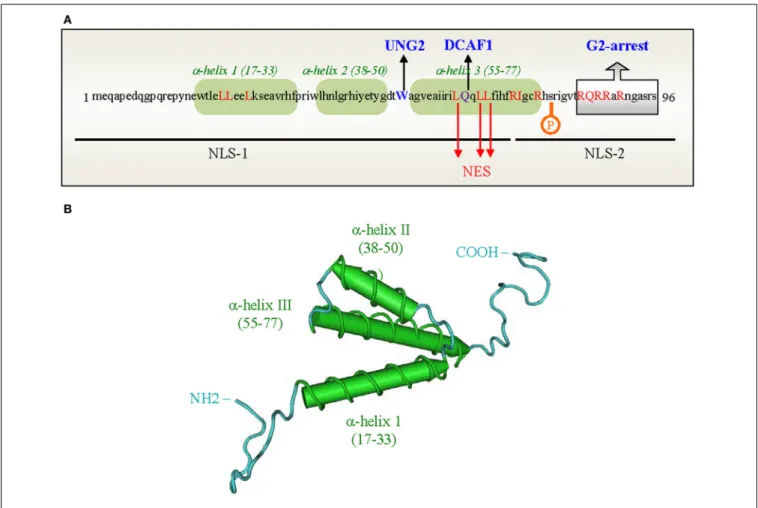
Figure

Documents relatifs