HAL Id: tel-01921990
https://tel.archives-ouvertes.fr/tel-01921990
Submitted on 14 Nov 2018HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Influence de variations de conditions environnementales
sur l’évolution des biofilms oraux
Darrène Nguyen
To cite this version:
Darrène Nguyen. Influence de variations de conditions environnementales sur l’évolution des biofilms oraux. Microbiologie et Parasitologie. Université de Bordeaux, 2018. Français. �NNT : 2018BORD0029�. �tel-01921990�
THÈSE PRÉSENTÉE POUR OBTENIR LE GRADE DE
DOCTEUR DE
L’UNIVERSITÉ DE BORDEAUX
ÉCOLE DOCTORALE des SCIENCES DE LA VIE ET DE LA SANTE SPÉCIALITÉ MICROBIOLOGIE-IMMUNOLOGIE
Par Darrène LOPEZ-NGUYEN
INFLUENCE DE VARIATIONS DE CONDITIONS
ENVIRONNEMENTALES SUR L’EVOLUTION DES BIOFILMS
ORAUX
Sous la direction du : Dr Marie-Cécile BADET
Soutenue le 28 Février 2018 à 14h30
Membres du jury :
Mme DUPUIS Véronique, PU-PH, Université de Bordeaux Présidente Mme PRECHEUR Isabelle, PU-PH, Université de Nice Rapporteur Mme ROQUES Christine, PU-PH, Université de Toulouse Rapporteur Mme NANCY Javotte, MCU-PH, Université de Bordeaux Examinateur
Remerciements
« Soyons reconnaissants aux personnes qui nous donnent du bonheur ; elles
sont les charmants jardiniers par qui nos âmes sont fleuries. »
Marcel Proust
Au Docteur Badet,Les mots me manquent pour vous exprimer ma gratitude. Merci d’avoir cru en moi. Merci de m’avoir fait confiance et de m’avoir offert l’opportunité de passer ces années d’études à vos côtés depuis la P2… Vous m'avez toujours témoigné d'un soutien sans faille et bienveillant ; je vous en remercie sincèrement. Cette thèse représente la consécration de 9 années de travaux dans votre laboratoire, 9 années dont je chérirai l’éternel souvenir.
Au Professeur Dupuis,
Je vous suis infiniment reconnaissante pour votre bienveillance et votre accompagnement durant ces années d’études et d’assistanat. Je vous remercie également très sincèrement d’avoir accepté de siéger à mon jury de thèse, malgré votre emploi du temps que je sais plus que chargé. Je garde en mémoire tous les conseils précieux que vous m'avez enseignés tout au long de mon parcours. C'est pour moi un honneur de vous compter aujourd'hui comme membre du jury de ma thèse de Doctorat.
Au Docteur Nancy,
Je vous suis très reconnaissante pour tout votre enseignement, vos encouragements, votre patience et votre bienveillance. Je vous remercie vivement d’avoir accepté de m’accompagner tout au long de cette thèse et de siéger à ma soutenance aujourd'hui. J'ai été très heureuse de partager mes vacations cliniques à Saint André à vos côtés. Votre dynamisme sans faille est pour moi un exemple de motivation.
Au Professeur Précheur,
Je vous suis extrêmement reconnaissante d’avoir accepté de juger ce travail, d’avoir reporté vos congés et de vous déplacer à Bordeaux afin que je puisse soutenir ma thèse aujourd’hui. C’est un réel honneur pour moi de vous avoir comme rapporteur, je vous en remercie très sincèrement et vous exprime ici toute ma gratitude et tout mon respect.
Au Professeur Roques,
Je vous remercie de tout cœur d’avoir accepté de juger mon travail en tant que rapporteur. Merci de m’avoir guidée lors de mon comité de mi-thèse, vos précieux conseils m’ont grandement aidé dans la suite de la réalisation des travaux. Je garde en mémoire de très bons moments passés à vos côtés à Nashville.
A Yoann, l’amour de ma vie, mon âme sœur, my one and only et le père de mon futur enfant. Merci de ton amour incommensurable, de ton soutien sans faille, et de ton aide de tous les instants. Je nous souhaite une nouvelle page de vie encore plus intense, joyeuse et heureuse que ces 14 ans inoubliables passés à tes côtés. Je t’aime.
A mes parents, mes modèles, mes appuis. Merci de votre amour infini, de votre dévotion, de votre accompagnement. Vous êtes ma fierté, sans vous, je n’en serai pas là où je suis. Je mesure à chaque seconde la chance inouïe que j’ai de vous avoir. Je nous souhaite d’être d’aussi bons parents que ce que vous êtes pour moi. Je vous aime.
A mon Yoyi, loin des yeux, mais tellement proche du cœur. Merci de ton amour et de ton oreille attentive. Tu arrives à être si près de moi, en habitant loin, et pour cela merci. Je suis heureuse que tu aies trouvé ta voie et je suis très fière d’être ta sœur. Je t’aime.
A mon Yanyan, mon p’tit frérot. Merci de ton amour et de ton soutien. Les moments que l’on partage sont toujours aussi intenses, et même en grandissant tu restes toujours mon Zouni. Je te souhaite beaucoup de réussite pour les mois qui viennent, saches que je serai toujours là pour t’aider et te soutenir. Je t’aime.
A Mami, je suis très fière et heureuse que tu puisses assister à ma thèse aujourd’hui. Merci de ton amour et de ta présence. Je souhaite que nous passions encore de merveilleux moments ensemble, 4 générations ensemble, c’est une chance inouïe que je chéris ! Je t’aime Mami.
A Ong Noi, Ba Noi, Ma Tu, Bac Hai, Chu Nam, Thim Nam and BoiBoi, Thank you for your support and the wonderful moments we always spend together! Even they are very few, they are very intense and I cherish every second of them! Let’s pray we can soon party and share delightful time together again. I love you from the bottom of my heart.
A mon Papi, parti bien trop tôt. Chaque nouvelle étape de ma vie est teintée d’une larme car je sais que tu n’es plus là pour le voir. Merci pour cet héritage que tu m’as laissé, cette force qui me conduit à toujours donner le meilleur de moi-même pour que tu sois fier de moi. Je t’aime et tu me manques.
A Ba Di, our Angel. I know that you are always protecting and guiding me. Thank you for blessing every second of my life. I wish you could physically be here with me. I love you and I miss you very hard.
A toute ma famille, soutien inconditionnel sans lequel je n’y arriverai pas.
--A Johan, pour tous tes conseils, ton soutien, ta bonne humeur et notre bonne entente. J’ai beaucoup apprécié (et continue d’apprécier) de travailler avec toi. Mille mercis pour ton aide dans l’élaboration de cette thèse, à chaque étape.
A Noëlie, je vous remercie très chaleureusement de tous vos conseils, de votre gentillesse et votre grande bienveillance à mon égard. Un immense merci pour tout ce que vous m’avez appris et pour ces supers vacations du mercredi passées ensemble !
A mes thésardes, les thésardes idéales, le trio de chic choc. Jennyfer, Samantha, Hinarii, sans vous, le labo n’aurait pas été ce qu’il a été ! Précision, application, motivation, une aide si précieuse, vous avez grandement contribué à cette thèse, et je vous en remercie du fond du cœur !
A Martine, pour ton aide et ta gentillesse tout au long de ces années. Je te souhaite beaucoup de bonheur et d’épanouissement dans le nouveau labo ☺
A Olivier Claisse, pour votre accueil, votre patience et votre aide précieuse dans toutes ces manips de PCR quantitative.
--A Julie, ma meilleure amie, toujours là à mes côtés. Merci d’être toi, et d’être là aujourd’hui, comme à chaque étape de ma vie. Je te souhaite le meilleur de ce que la vie peut offrir avec Vincent.
A Aurore, à ces manips et à ce voyage à Nashville ! Et dire qu’on serait passées à côté de cette magnifique amitié sinon… Je te souhaite beaucoup de bonheur, tu le mérites.
A JC, à Marie, à vos retrouvailles avec Yoann, à cette amitié grandissante et déjà si intense. Merci de votre soutien… Et à ma mascotte préférée, à la princesse de tous les temps, j’ai nommé ma Romance adorée !
--
A tous ceux qui m’ont soutenue et que je n’ai pas la place de citer. A tous ceux qui ont fait le déplacement aujourd’hui pour venir à cette soutenance. Merci, je vous porte dans mon cœur.
Table des matières
Introduction ... 8
Rappel des Connaissances ... 10
Ecosystème buccal ... 10 Rappels anatomo-histologiques ... 12 Milieu buccal ... 13 Flore microbienne ... 15 Biofilm buccal ... 19 Interactions bactériennes ... 22
Pouvoir pathogène du biofilm dentaire ... 30
Etude des modèles de biofilms ... 36
1. Modèles de biofilms in vitro ... 36
2. Modèles de biofilms ex vivo ... 39
3. État des recherches en microbiologie orale : les modèles de biofilms oraux ... 39
4. Identification bactérienne ... 46 Matériel et Méthodes ... 77 Matériel ... 77 1. Souches utilisées ... 77 2. Milieux de culture ... 77 Modèle de biofilms ... 77 1. Description ... 77
2. Mise en place du biofilm ... 79
3. Recueil du biofilm ... 79
4. Etude et identification ... 80
5. Recherches pour identifier F. nucleatum ... 93
Variations de conditions environnementales... 94
1. Influence de conditions pro-cariogènes ... 94
2. Introduction d’un candidat probiotique ... 95
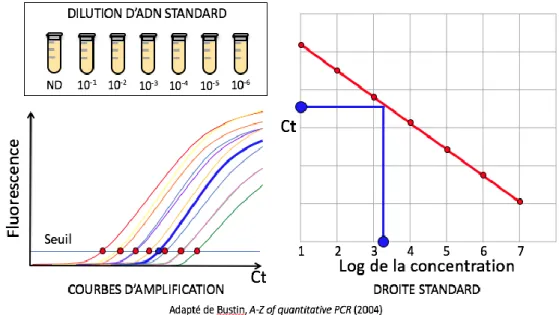
Résultats ... 118 Protocole 1 ... 120 1. Disque ... 120 2. Milieu ... 120 Protocole 2 ... 121 1. Type 1 ... 121 2. Type 2 ... 121 Protocole 3 ... 122 1. Type 1 ... 122 2. Type 2 ... 122 3. Type 3 ... 122 4. Type 4 ... 123 5. Type 5 ... 123 6. PCR quantitative ... 124
Recherches pour identifier F. nucleatum... 135
1. Observation au MO du dépôt après coloration de Gram ... 135
2. Isolement de F. nucleatum sur gélose TGY vert brillant ... 135
3. Co-cultures ... 135
Variations de conditions environnementales... 136
1. Influence des paramètres pro-cariogènes ... 136
2. Effets du candidat probiotique sur le biofilm selon la séquence d’inoculation ... 139
3. Résultats de qPCR des prélèvements sur géloses ... 140
Liste de tableaux et figures ... 148
Annexes ... 149
Publications et Communications ... 160
Introduction
L’écosystème buccal est un environnement complexe dans lequel cohabitent plus de 700 espèces de bactéries différentes [1][2]. Le biofilm, en traduction littérale : pellicule de vie [3], est composé par la coexistence de toutes ces bactéries, dans un même habitat. Il est communément appelé « plaque dentaire ». Un déséquilibre au sein de ce biofilm est à l’origine des principales maladies de la cavité buccale : les maladies carieuses et parodontales [4].
Pendant plusieurs années, l’étude de l’écosystème buccal s’est faite par une approche réductionniste : les microbiologistes étudiaient les espèces bactériennes individuellement. Cette stratégie a permis d’examiner et de comprendre tous les différents composants de cet écosystème, sans pour autant pouvoir appliquer les conclusions de ces études au biofilm buccal dans son ensemble. En effet, les bactéries ne se comportent pas de la même façon lorsqu’elles sont à l’état planctonique, ou lorsqu’elles sont organisées en biofilm.
La flore microbienne buccale est reconnue comme étant l’une des plus complexes dans le corps humain. La multitude d’espèces en présence complique l’étude de ce biofilm. En effet, sa reproduction in vitro est rendue difficile par la complexité des relations entre chacune des espèces. De plus, son recueil, et ses décomptes qualitatif et quantitatif restent très délicats.
Dans la littérature sont décrits plusieurs modèles de biofilm [5]: mono-espèces, pluri-espèces statiques, et pluri-espèces dynamiques, à côté des modèles ex vivo, et in vivo. Chaque type de modèle présente des avantages et des inconvénients.
Les modèles pluri-espèces dynamiques in vitro ont l’avantage de se rapprocher des conditions retrouvées in vivo, permettant un certain flux de milieu, le contrôle de paramètres tels que le pH et la température, ainsi que l’élimination des déchets produits. Cependant, ces modèles restent très onéreux et difficiles de mise en place, ce qui complique l’étude du biofilm buccal. Egalement, l’identification des bactéries mises en présence reste un sujet d’étude délicat : en effet, les méthodes traditionnelles de culture montrent des limites, et ne permettent pas une analyse quantitative des résultats, essentielle à la compréhension des phénomènes mis en jeu dans cet écosystème.
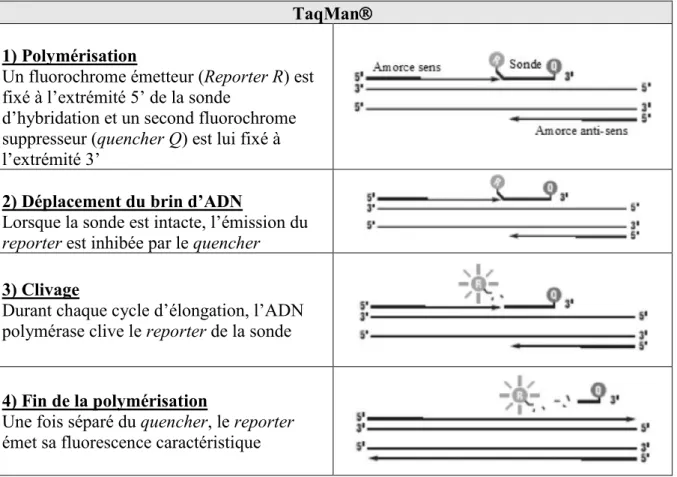
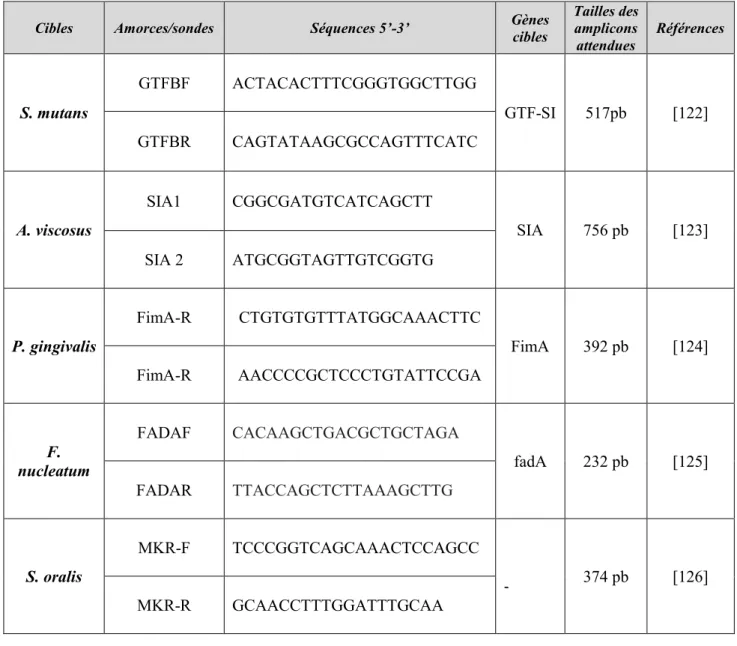
Le but de notre travail ici est l’élaboration d’un modèle de biofilm pluri-espèces dynamique, fiable et reproductible, facile à mettre en œuvre et moins onéreux que ceux décrits dans la littérature. Ce modèle de biofilm devrait permettre l’étude de l’influence de variations de conditions environnementales sur ce dernier, ainsi que celle de candidats probiotiques ayant déjà prouvé leur efficacité sur des supports in vitro statiques. Enfin, toujours dans une optique de simplification, il est envisagé de comparer les différentes méthodes d’identification des biofilms formés (méthodes de culture traditionnelle, PCR conventionnelle, spectrométrie de masse MALDI-TOF, et qPCR), afin d’établir un protocole d’identification reproductible permettant une analyse qualitative et quantitative des résultats.
Rappel des Connaissances
Ecosystème buccal
Dans la cavité buccale, la plaque dentaire est organisée sous forme de biofilm. Il s’agit de micro-organismes d’une ou plusieurs espèces adhérant à une surface submergée ou soumise à un environnement aqueux [6].
Contrairement à celles évoluant à l’état planctonique, le biofilm a l’avantage d’offrir aux bactéries une protection face à des facteurs environnementaux défavorables (mécanismes de défense de l’hôte, substances toxiques, antibiotiques, etc...) et de faciliter les échanges entre elles [7].
Le biofilm est composé d’une fraction cellulaire, les micro-colonies bactériennes qui représentent 15 à 20% du volume et qui ont une disposition bien définie au sein d’une composante acellulaire, et de la matrice qui représente les 75 à 80% du volume restant [7].
Un milligramme de biofilm contient environ cent millions à un milliard de bactéries [8]. Le nombre d’espèces microbiennes de la cavité orale est estimé entre 500 et 700 [1], [9].
La matrice est formée à 80% d’eau et à 20% de polymères, principalement des polysaccharides mais aussi des protéines, des lipides, des oligoéléments, des éléments minéraux et des acides nucléiques produits par certains micro-organismes. Elle a, en particulier, pour rôle de permettre la survie des micro-organismes en favorisant leur nutrition et leur croissance [6] [8].
Les polysaccharides extracellulaires comprennent des fructanes (polymères de fructose) et des glucanes (polymères de glucose) [8].
Pour caractériser précisément l’écosystème buccal, il est important d’introduire plusieurs autres concepts :
« Hôte » : l’hôte est un organisme qui héberge un parasite, un partenaire synergique ou un partenaire commensal. Cet hébergement est nécessaire à l’accomplissement du cycle de vie de l’hébergé.
« Commensalisme » : étymologiquement « partage de la même table » décrit une interaction entre deux espèces dans laquelle l’hôte ne tire aucun bénéfice de la présence de l’organisme étranger qu’il héberge.
Certaines espèces commensales dites indigènes sont d'ailleurs des éléments au rôle ambigu, car elles peuvent aussi bien être nécessaires à l'organisme humain dans certaines situations, que pathogènes à son égard dans d’autres situations.
« Saprophyte » : organisme se nourrissant de matière organique en décomposition par libération d’enzymes.
« Symbiose » : la symbiose exprime le bénéfice mutuel entre au moins deux types microbiens. C’est le cas de la croissance des anaérobies au sein d’aérobies, ces derniers consomment l'oxygène et créent un potentiel d'oxydo-réduction faible favorisant la croissance des premiers.
« Antibiose » : l’antibiose répond par définition à un antagonisme entre espèces. Les substances supports de ce phénomène pourraient être responsables de la stabilité des communautés.
« Synergisme » : le synergisme qualifie une relation bénéfique entre espèces pathogènes qui, isolément, seraient incapables de provoquer une pathologie.
« Parasitisme » : Faculté pour un organisme de se développer aux dépens d’un autre organisme.
Dans l’écosystème buccal, le Biotope est constitué par la cavité buccale de l’espèce Homo Sapiens Sapiens qui est ainsi qualifié d’hôte. La biocénose est constituée d’un ensemble de plus de 700 espèces bactériennes, majoritairement commensales mais donc certaines peuvent s’avérer pathogènes à la faveur de modifications physico-chimiques et moléculaires de l’écosystème permettant leur développement.
Rappels anatomo-histologiques
La bouche est une cavité appendue à la partie inférieure du massif facial, comportant les dents, la langue, les gencives et les muqueuses buccales. Elle est ouverte à l’avant par la fente labiale, limitée en bas par le plancher de la langue, en haut par le palais dur et mou qui la sépare des fosses nasales et des sinus maxillaires, postérieurement par l’oropharynx dont l’entrée est limitée par les tonsilles palatines et latéralement par les joues.
Majoritairement revêtue d’une muqueuse richement vascularisée, elle est constamment maintenue dans un milieu humide par l’action des glandes salivaires principales (parotides, sub-mandibulaires et sub-linguales) et accessoires qui tapissent l’intégralité de la muqueuse buccale. Les dents sont intimement liées à leur parodonte.
Le parodonte est constitué de quatre tissus :
- L’os alvéolaire : composé d’une corticale et d’un os trabéculaire.
- La gencive qui est une fibro-muqueuse kératinisée en raison des contraintes mécaniques et physico-chimiques qu’elle subit. Sous la couche épithéliale se trouve un tissu conjonctif ou chorion en deçà de la membrane basale.
- Le cément est un tissu minéralisé recouvrant la surface radiculaire des dents issu embryologiquement de la gaine épithéliale de Hertwig. Il reçoit des fibres en provenance du desmodonte et participe ainsi à l’attache épithéliale.
- Le desmodonte ou ligament alvéolo-dentaire unit par des fibres de collagène multidirectionnelles la dent à l’os alvéolaire.
Par ailleurs ces quatre tissus sont richement vascularisés et innervés par des pédicules vasculo-nerveux cheminant en leur sein.
La gencive du sillon gingivo-dentaire cesse d’être kératinisée et prend le nom de gencive attachée. Elle se poursuit apicalement par l’attache épithéliale qui laisse place à l’attache conjonctive puis au desmodonte.
C’est dans cette zone appelée espace biologique que se trouve le biofilm parodontal responsable des gingivites et des parodontites liées à la plaque. Ce biofilm est l’objet de nos études.
En réalité, toutes les zones à l’abri du nettoyage dentaire prophylactique sont susceptibles d’abriter des niches écologiques propices au développement d’un biofilm, en particulier : les capuchons des dernières molaires, le sillon gingivo-dentaire (et son développement pathologique, la poche parodontale), les puits des faces occlusales, les fissures de l'émail, les points de contact interdentaires, les caries, les reconstitutions iatrogènes (amalgames, composites ou coiffes débordants) et les chevauchements dentaires, les zones exposées, dotées de villosités, comme le dos de la langue, ou apparemment lisses, comme les gencives, constituent des supports appréciés des micro-organismes, en raison des récepteurs d'adhérence que comportent leurs cellules superficielles.
Milieu buccal
Chez le sujet sain la température sous-gingivale moyenne se situe à 34.7° C± 0°6 (soit de 33.4°C à 36.1°C). Cette température s’élève de quelques dixièmes de degrés en présence d’une inflammation. Par ailleurs, les acides lactique, acétique, formique, propionique, et butyrique présents après mastication d’aliments gras et sucrés sont responsables d’une augmentation de la température sous-gingivale.
L’écosystème buccal présente, en outre, un haut niveau d’hygrométrie grâce à la salive. Son
important pouvoir tampon conféré par sa forte concentration en Acide
Carbonique/Hydrogénocarbonate permet une homéostasie du pH entre 6,7 et 8,5 (dont la prise de nourriture est la principale cause de variation). Outre 99% d’eau, la salive apporte à l’écosystème buccal des sels minéraux dont des carbonates de calcium et des phosphates qui concourent à la formation du tartre mais également des mucines et des glycoprotéines en grande quantité. Plus l’on progresse profondément dans le sillon gingivo-dentaire et a fortiori, dans la poche parodontale, plus la pression partielle d’oxygène décroit et plus la pression partielle en dioxyde de carbone s’accroit.
Précisons qu’à l’échelle cellulaire et moléculaire, le sillon gingivo-dentaire d’un hôte immunocompétent est riche en cellules de l’immunité innée et acquise ainsi qu’en molécules antibactériennes non spécifiques telles que le lysozyme, la sialoperoxydase, les agglutinines salivaires d’une part et spécifiques d’autre part telles que les inhibiteurs de protéases, des immunoglobulines, en particulier IgA et IgG, et divers facteurs de croissances cellulaires. Elles proviennent de la salive mais aussi du liquide créviculaire ou fluide gingival qui est un exsudat du sérum sanguin secrété dans le sillon.
D’après Marsh & Martin [10]
Figure 1 : Description du milieu buccal
Joues, lèvres, palais
- La salive a une influence majeure - La microflore est peu diversifiée - Présence d’anaérobies facultatives - Streptococcus ssp. prédominants - Quelques pathogènes parodontaux
persistent en envahissant les cellules
Dents
- Surfaces non desquamantes - Sites rétentifs (ex : sillons)
- Microflores diverses, dépendant du site - Beaucoup d’anaérobies strictes
- Influence de la salive et du fluide gingival
- Streptococcus, Actinomyces, Veillonella, Fusobacterium…
Langue
- Présence de nombreuses papilles - Quelques sites anaérobies
- Desquamation - Microflores diverses
- Anaérobies facultatives et strictes
Flore microbienne
L’estimation du nombre total d’espèces bactériennes présentes dans notre monde est de l’ordre du million même si, à ce jour, nous n’en connaissons que 10.000 et que seules 538 sont considérées comme infectieuses pour l’homme. Environ 1010 bactéries résident dans le biotope
buccal d’un individu sain. Elles s’acquièrent initialement par contamination maternelle vagino-buccale puis, par la suite, la cavité orale est accessible aux bactéries aéroportées (poussières, aérosols…), manuportées ou contenues dans les aliments (fruits, viandes crues…). Elle est par conséquent extrêmement variée. On considère que plus de 700 espèces bactériennes différentes sont hébergées dans une bouche saine. Selon la localisation et l’état pathologique de l’hôte, la flore buccale peut être drastiquement différente en qualité et en quantité.
Les nombreuses modifications physiologiques qui ont lieu lors de la grossesse puis au moment de la naissance, particulièrement hormonales, métaboliques et immunitaires sont fortement influencées et à leur tour influencent fortement le microbiote, que celui-ci soit buccal, intestinal, vaginal ou placentaire [11], [12] .
Contrairement à la croyance soutenue dès 1900 par Tissier partagée jusqu’à une période récente par la communauté scientifique, l’on ne nait pas indemne de contamination microbienne. Des preuves de présence microbienne au sein du complexe foeto-placentaire ont été apportées par Hu et al. en 2013 [13] et Aagaard et al. en 2014 [14].
Cette dernière étude a mis en évidence que le microbiome placentaire avait un profil taxonomique comparable à celui du microbiome oral. Une forte similitude a en effet été observée entre le placenta et la langue, les amygdales, la salive, et les profils taxonomiques sous-gingivaux.
La composition des microbiotes de chaque site du corps humain est soumise à des paramètres environnementaux tels que le pH, la pression partielle en dioxygène et en dioxyde de carbone, le taux d'humidité, la température ainsi que la présence de nutriments. Il est démontré que le microbiote affecte le métabolisme de l’hôte [15], module son immunité (et y contribue parfois) [16] ainsi que ses taux hormonaux [17].
La modification de la composition des microbiotes peut entraîner des effets délétères pour la santé tels que l’obésité, le syndrome du côlon irritable, le diabète et le syndrome métabolique.
Pendant la grossesse, les taux de certaines hormones, particulièrement de progestérone et d’œstrogène s’accroissent de façon considérable. Ces modifications provoquent une modulation de la réponse immunitaire qui est nécessaire au développement du propre système immunitaire du fœtus tout en continuant à protéger les infections pouvant mettre en danger la mère et le fœtus.
Par ailleurs, les modifications métaboliques associées à l’état gravidique s’apparentent fortement à celles qui décrivent le Syndrome Métabolique : prise de poids, augmentation de la glycémie à jeun, résistance à l’insuline, intolérance au glucose, inflammation modérée et modifications hormonales [18]–[21].
Parallèlement aux modifications immunitaires, endocrines et métaboliques précédemment décrites, les microbiotes des différents sites du corps connaissent également des modifications notables.
Citons en particulier pour le microbiote oral :
• Augmentation du nombre des sept espèces bactériennes les plus courantes à tous les trimestres de la grossesse, par rapport à la femme non gravidique [22].
• Pourcentage des bactéries pathogènes Porphyromonas gingivalis et Aggregatibacter actinomycetemcomitans dans la plaque sous-gingivale significativement plus élevé lors des 6 premiers mois de grossesse par rapport à la femme non enceinte [22], ainsi qu’au troisième trimestre pour Aggregatibacter actinomycetemcomitans [23].
• La présence de parodontite maternelle augmente le risque d’accouchement prématuré [24]. El Attar en 1976 [25] puis Kim et Amar en 2006 [26] suggèrent que les LPS (lipopolysaccharides) des bactéries Gram négatif telles que Porphyromonas gingivalis pourraient entraîner la production de prostaglandines et autres médiateurs inflammatoires circulants. Aagaard et al. [14] suggèrent une possible dissémination hématogène de bactéries depuis la bouche vers le placenta comme facteur favorisant les naissances prématurées.
Pour le microbiote vaginal :
• Diminution globale de la diversité microbienne chez la femme enceinte, en particulier au début de la grossesse avec une normalisation au cours du troisième trimestre [27]
• Diminution du pH vaginal et augmentation des sécrétions vaginales [28], [29] • Corrélation entre diminution de Lactobacillus et accouchement prématuré [30], [31] • Modifications postpartum qui s’observent jusqu’à un an après l’accouchement :
diminution du pourcentage de Lactobacillus et apparition ou augmentation du pourcentage de Peptoniphilus, Prevotella et Anaerococcus [32] et Actinobacteria [33].
Pour le microbiote placentaire :
• Corrélation entre une présence importante de germes alpha-hémolytiques placentaires et naissance prématurée [34].
• La comparaison des microbiotes de placenta dont la grossesse s’est poursuivie jusqu’au terme avec des placentas de naissances prématurées montre une composition significativement différente : Burkholderia, Streptosporangium et Anaeromyxobacter sont plus présents lors des accouchements prématurés ; Paenibacillus est davantage présent dans les placentas ayant atteint le terme [35], [36]
Rappelons que les complications liées à la prématurité sont les premières causes de mortalité néonatale. Une étude de Costello et al. montrait que les bébés prématurés ayant un faible poids à la naissance présentaient dans le mois suivant leur naissance une forte proportion de Staphylococcus dans la cavité orale et une forte proportion de Streptococcus sur la peau contrairement à leur habitat classique [37] .
A l’instar de l’évolution des microbiotes de la mère au cours de la grossesse, le microbiote de la cavité orale du nouveau-né va fortement évoluer depuis la naissance jusqu'à l’âge de 2 ans, âge à partir duquel s’opère une lente stabilisation du microbiote, selon Nuriel-Ohayon et al. [12]
Plusieurs études suggèrent une transmission materno-foetale de bactéries pendant la grossesse [38]–[40].
Généralement, le microbiote intestinal du nouveau-né par voie vaginale est composé de bactéries appartenant au microbiote vaginal maternel telles que Prevotella, Sneathia et Lactobacillus. Il inclut également des bactéries présentes dans l’intestin maternel [41], [42].
A contrario, les nouveau-nés par césarienne présentent un microbiote oral ressemblant à celui de la peau maternelle, particulièrement avec la présence de Propionibacterium, Corynebacterium et Streptococcus [33], [43].
En 2011, Barfod et al. ont montré que la prévalence salivaire à la naissance de Streptococcus salivarius, Lactobacillus curvata, Lactobacillus salivarius et Lactobacillus casei était significativement plus élevé chez les bébés nés par voie vaginale [44].
Les bébés nés par césarienne montrent un retard de colonisation orale du phylum Bacteroidetes, une moindre diversité alpha pendant les deux premières années de vie, ainsi que des taux moins élevés de chémokines CXCL10 et CXCL11 liés aux Lymphocytes T helper, ce qui suggère un retard de maturation du système immunitaire [45], [46].
L’allaitement maternel est une source importante de colonisation bactérienne de la cavité orale du bébé. Les bactéries les plus présentes dans le colostrum (premier lait très riche en anticorps) sont Streptococcus, Staphylococcus, Lactococcus et Weisella tandis que le lait produit 6 mois post-partum est surtout riche en Veillonella et Prevotella [47].
Les bébés de 3 mois allaités au sein contiennent des Lactobacilles ayant des propriétés antimicrobiennes dans la cavité orale contrairement aux enfants allaités artificiellement [48], [49].
Enfin, le passage à une alimentation diversifiée est également un moment de grande diversification et de maturation du microbiote oral [43].
Biofilm buccal
Les bactéries peuvent vivre et se développer selon deux modes distincts, passant de l’un à l’autre à différentes étapes du cycle de vie :
- L'état planctonique, libre flottaison unicellulaire dans un médium fluide. - Les biofilms.
Dans des conditions optimales de croissance le biofilm peut rapidement devenir macroscopiquement visible. L'organisation du biofilm bactérien fut mise en évidence pour la première fois en 1976 par Marshall. Il est estimé que 80% de la biomasse bactérienne de la planète réside au sein de biofilms.
Les biofilms sont majoritairement observés dans les milieux aqueux et se développent sur tout type de surface naturelle ou manufacturée, minérale ou organique. Les microbiologistes distinguent habituellement deux types de biofilms : les biofilms mono espèces qui ne sont composés que d’une espèce et les biofilms pluri espèces au sein desquels plusieurs espèces bactériennes sont associées.
99,9% des bactéries de la cavité buccale sont organisées au sein d’un biofilm pluri espèces. Ces bactéries sont caractérisées par la sécrétion d’une matrice protectrice de polymères exo-cellulaires constitués de polysaccarides hydratés qui constitue 85% du biofilm. Les 15% restants constituent la fraction cellulaire.
Macroscopiquement, le biofilm également appelé plaque dentaire, a un aspect mou, jaunâtre et collant, capable de se fixer tant sur les surfaces dentaires que sur des éléments prothétiques, orthodontiques ou sur certaines obturations, et présente une odeur nauséabonde de décomposition organique. Le recours à des techniques de microscopie confocale à balayage laser a pu montrer que la plaque dentaire a une architecture similaire à celle des biofilms provenant d’autres sites. Ainsi, il se développe souvent de façon hétérogène sous forme de micro-colonies discontinues, séparées par des espaces permettant la circulation de liquides et de molécules.
Le développement du biofilm dentaire est un processus dynamique qui peut être décomposé en plusieurs étapes successives :
➢ Etape 1 : Adhésion réversible
Trente secondes après le nettoyage des surfaces bucco-dentaires, il se forme une fine couche (inférieure à 1 micron) appelée pellicule acquise exogène. La composition de cette pellicule salivaire est dépendante de la surface qu’elle recouvre. Elle peut être composée de centaines de protéines, glycoprotéines, glycolipides et lipides différents présents dans la salive ou encore de composants issus des cellules bactériennes mais aussi de l’hôte. Ce sont autant d’éléments qui vont faciliter l’adhésion des colonisateurs primaires [50]
Le rapprochement spécifique via des récepteurs membranaires bactériens est appelé chimiotactisme. D’autres facteurs sont impliqués dans l’attachement initial des bactéries aux surfaces dentaires : la force hydrodynamique, les forces électrostatiques de Van der Waals (attraction-répulsion) [8] et les interactions hydrophobes. A cette étape, l’adhésion reste réversible et les bactéries peuvent encore retourner à l’état planctonique [6].
En ce qui concerne le biofilm dentaire, les colonisateurs primaires sont pour 60% des streptocoques, avec principalement Streptococcus oralis, Streptococcus mitis, Streptococcus sanguinis, Streptococcus parasanguinis et Streptococcus gordonii. [50][51] Les autres bactéries retrouvées sont des bactéries du genre Actinomyces [52][53].
➢ Etape 2 : Adhésion irréversible
Essentiellement, à cette étape les bactéries produisent une grande quantité d’exopolymères, la matrice extracellulaire, qui permettent de renforcer la cohésion hétérogène inter bactérienne et conduisent à une adhésion irréversible des bactéries [8].
C’est l’attachement à la surface qui induit l’expression de phénotypes différents comme cette synthèse d’exo polysaccharides. Par ailleurs, l’adhérence irréversible des bactéries peut aussi être liée à des modifications physiologiques au niveau des organites de surface. Par exemple, les flagelles qui permettent la mobilité sont remplacés par des pili de type IV [6].
➢ Etape 3 : Formation des micro-colonies
Les colonisateurs primaires, constituent les points d’ancrage aux colonisateurs secondaires, notamment grâce à l’expression d’adhésines. Ces derniers peuvent être sous forme de cellules planctoniques individualisées ou de co-agrégats cellulaires.
Les Streptococci utilisent des protéines de surface pour adhérer aux glycoprotéines présentes dans l’environnement oral [54]. Actinomyces naeslundii (oris) utilisent les fimbriae de type 1 afin d’adhérer aux stathérines et aux Protéines Riches en Proline présentes dans la pellicule acquise exogène formée sur les surfaces amélaires [55]. Les Actinomyces sont également capables d’interagir par leur fimbriae de type II avec les phosphopolysaccharides exprimés à la surface de Streptococcus oralis, Streptococcus mitis et Streptococcus sanguinis [56].
C’est par ces mécanismes de co-adhésion que se forment les micro-colonies [6]. Cela conduit à une diversification des communautés du biofilm en développement [57].
Les premiers colonisateurs secondaires comme Fusobacterium nucleatum peuvent faire le lien entre les colonisateurs primaires et d’autres colonisateurs secondaires [58]. Veillonella est également un des premiers colonisateurs secondaires puisqu’il a une affinité particulière pour les streptocoques [59][60]. Candida albicans se lie aux streptocoques et aux autres colonisateurs primaires [61]. Des interactions physiques, une signalisation chimique et des réponses transcriptionnelles permettent la liaison entre streptocoques et Porphyromonas gingivalis. Les fimbriae majeures de P. gingivalis interagissent avec les glyceraldehyde-3-phosphate dehydrogénases présentes à la surface des streptocoques [62], tandis que les fimbriae mineures se lient aux adhésines SspB [63].
➢ Etape 4 : Maturation
Pendant cette phase, la taille de la structure connaît une forte augmentation de son volume. Ceci résulte des multiplications bactériennes, mais aussi de l’épaississement de la matrice extracellulaire. Cela entraine une modification des gradients d’oxygène, de substrats et voire de pH au sein du biofilm. C’est lors de cette étape également que les phénomènes de communications intercellulaires, se mettent en place durablement. La biodiversité microbienne évolue alors quantitativement et qualitativement au cours de la maturation du biofilm. [8]
Le biofilm acquiert sa structure mature avec la formation de canaux internes entre les micro-colonies et de pores. [6] Ces canaux permettent le passage, la diffusion à travers le biofilm de l’eau, des nutriments et autres substances aux cellules. [7]
➢ Etape 5 : Détachement cellulaire
Après une phase de croissance importante, le biofilm connaît un appauvrissement au niveau nutritionnel. Ceci entraine alors le détachement d’un grand nombre de cellules. Ce phénomène est appelé « sloughing ». Les bactéries détachées pourront ainsi coloniser d’autres sites [8].
Interactions bactériennes
Comme vu précédemment, l’architecture d’un biofilm n’est pas due au hasard. Elle dépend pour partie des métabolismes microbiens et des affinités des divers micro-organismes entre eux, qui conduisent à la formation d’une structure tridimensionnelle bien organisée [6]. Ce sont donc de nombreuses relations interdépendantes, aussi bien synergiques qu’antagonistes, qui régissent le développement, le maintien et l’évolution de cet écosystème [64].
➢ Coopération bactérienne
• Co-adhésion et co-agrégation
L’agrégation entre genres bactériens repose sur une reconnaissance de cellule à cellule liée à la reconnaissance spécifique des polysaccharides bactériens. Néanmoins, deux paires de bactéries peuvent avoir une même bactérie en commun puisqu’elles ne reconnaissent pas forcément le même site sur cette bactérie commune [65].
Les phénomènes de co-adhésion et de co-agrégation participent donc à la survie bactérienne et à l’épaississement du biofilm.
Le mécanisme de co-agrégation se produit lors de la formation du biofilm entre les colonisateurs secondaires qui ne peuvent se fixer directement à un colonisateur primaire [65].
En particulier, F. nucleatum est une des espèces les plus connues faisant le lien entre les streptocoques et les bactéries anaérobies strictes [58]. Sans elle, la quantité d’anaérobies strictes dans la cavité orale chuterait [64][65].
La co-agrégation établit une proximité entre les cellules voisines et leurs métabolites [64]. • Communication métabolique
La communication métabolique est l’utilisation comme source d’énergie d’un métabolite ou substrat produit par un micro-organisme par un autre [64].
Les exopolysaccharides du biofilm constituent le milieu de cette communication [65].
Plusieurs études ont montré l’association symbiotique entre Veillonella et les streptocoques via l’acide lactique produit par ces derniers [59][60][64]. De la même manière, les streptocoques permettent la survie de Aggregatibacter actynomicetemcomitans [57].
De plus, Treponema denticola et Porphyromonas gingivalis sont souvent retrouvés ensemble dans le biofilm dentaire chez des sujets atteints de parodontite.
Le rapprochement entre ces deux espèces serait dû à la production d’un facteur de croissance, l’acide succinique, par T. denticola et qui serait utilisé pour la formation de composants de l’enveloppe cellulaire de P. gingivalis [64].
Des échanges en oxygène se produisent également. Ils permettent la survie des bactéries anaérobies strictes par consommation de l’oxygène par les bactéries aérobies. Cela conduit à un appauvrissement local en oxygène et à un gradient de potentiel redox qui produit un environnement propice aux bactéries anaérobies strictes [65].
Aussi, certaines espèces peuvent aider d’autres espèces à survivre en modifiant favorablement l’environnement local par réduction de l’acidité du milieu. Par exemple, F. nucleatum et Prevotella intermedia peuvent croître dans des conditions de pH allant de 5,0 à 7,0 alors que P. gingivalis est très sensible au pH inférieur à 6,5. Cependant, en produisant de l’ammoniac et des acides organiques, F. nucleatum et P. intermedia créent alors un environnement plus neutre et plus propice à P. gingivalis et à d’autres bactéries acido-sensibles [65].
D’autres types de communications métaboliques inter-espèces impliquent par exemple les quinones exogènes ou la vitamine K qui n’est pas produite par l’organisme humain [64].
• Résistance aux facteurs externes et échanges génétiques
Au sein des bactéries pathogènes, le matériel génétique est porteur d’informations, notamment concernant les facteurs de pathogénicité.
Les plasmides contiennent de l’ADN extra chromosomique. Ils sont capables de réplication autonome. Ils sont le plus souvent circulaires, et se transmettent de cellule mère à cellule fille au cours de la croissance.
Les îlots génomiques sont des zones du chromosome bactérien ayant des caractéristiques différentes du reste du génome.
L’Integrative and Conjugative Element (ICE) est acquis par transformation, transduction ou conjugaison. Il code pour des informations biochimiques ou des facteurs de pathogénicité.
Les transferts de gènes sont unidirectionnels, d’un donneur à un receveur. Le donneur donne rarement un chromosome entier. Ce phénomène peut se réaliser entre espèces différentes.
On parle de transformation lorsque le transfert de gènes résulte de l’intégration de fragments d’ADN d’un donneur mort ou vivant. La taille de l’ADN, sa sensibilité aux nucléases, et la compétence du récepteur peuvent affecter cette transformation.
On appelle conjugaison le transfert de gènes d’un donneur à un receveur par contact direct entre cellule.
La transduction correspond à un transfert de gènes d’un donneur à un receveur par l’intermédiaire d’un bactériophage.
La conversion lysogénique définit le transfert d’un bactériophage à une bactérie.
Elle nécessite des enzymes spéciales telles qu’une intégrase ou une transposase. Il existe deux types de transposition :
- La transposition conservatrice : une séquence d’ADN est transférée d’un site à un autre, entre un site donneur et un site accepteur.
- La transposition réplicative : l’élément transposable est transféré d’un site à un autre, tout en restant au site original. Cela conduit à une augmentation du nombre de copies de l’élément transposable.
En s’associant dans un biofilm, les bactéries bénéficient d’une résistance très supérieure à celle des bactéries à l’état planctonique.
En effet, les exopolysaccharides qui forment la matrice extracellulaire offrent une barrière relativement hermétique contre la diffusion d’agents antimicrobiens, les environnements difficiles, et les réponses immunitaires de l’hôte [65]. De plus, cette fonction de barrière est renforcée par la rétention dans le biofilm de facteurs de résistance comme des enzymes d’inactivation des biocides [64]. Par ailleurs, plus le biofilm est complexe et les espèces diverses, et plus grande est cette résistance [65].
Concernant la résistance aux antibiotiques, il a été observé que l’inhibition de la croissance d’une souche bactérienne organisée en biofilm nécessitait une concentration d’antibiotique environ 250 fois plus importante que pour la même souche mais évoluant à l’état planctonique [64].
La dégradation des antibiotiques par certaines espèces bénéficie aux autres espèces voisines. De plus, la proximité entre les micro-organismes au sein d’un biofilm favorise le transfert horizontal de gènes par les mécanismes de conjugaison, de transduction ou de transformation. Chez les streptocoques, l’échange entre individus de plasmides contenant des gènes de résistance par conjugaison est bien connu [64].
➢ Compétition bactérienne
Chaque espèce bactérienne possède ses propres stratégies pour survivre face aux autres dans un espace et une disponibilité en nutriments limités.
• Compétition pour l’adhésion aux surfaces
Les bactéries ayant la capacité de se lier aux protéines de la pellicule acquise exogène sont plus aptes à se fixer aux surfaces dentaires et à former la plaque dentaire précoce. `
C’est le cas de nombreux streptocoques qui sont capables de reconnaître les protéines salivaires telles que l’alpha-amylase, les protéines riches en proline ou les glycoprotéines riches
• Compétition pour la co-agrégation
Lorsque deux bactéries reconnaissent les mêmes récepteurs d’une bactérie, alors elles entrent en compétition pour ce même site [65].
• Compétition pour les nutriments
Dans la cavité buccale, les nutriments peuvent provenir de l’ingestion des aliments au cours des repas, de la salive ou des métabolites produits par les micro-organismes. Il peut exister des grandes variations dans l’apport des nutriments, et on assiste à une succession de phases « fastueuses » et de « famine », qui implique la nécessité d’adaptation, et explique l’apparition de compétition nutritionnelle. La disponibilité de ces différents nutriments selon le site joue un rôle important dans la composition du biofilm. En effet, les organismes capables de métaboliser les nutriments présents dans une niche écologique spécifique bénéficient d’un avantage indéniable quant à leur survie par rapport à ceux ne pouvant métaboliser les nutriments environnants. De plus, une espèce bactérienne ayant plus d’affinité pour une ressource aura un avantage compétitif vis-à-vis d’autres espèces [67].
• Bactériocines, lantibiotiques et autres métabolites d’inhibition
Pasteur et Joubert avaient découvert l’interaction antagoniste entre Escherichia coli et Bacillus anthracis. Elle provenait de protéines douées d’activité bactéricide, contre des bactéries d’espèce proches.
Les micro-organismes du biofilm sont aussi capables de produire des métabolites qui inhibent la croissance d’autres micro-organismes. Les plus létaux d’entre eux sont les bactériocines et le peroxyde d’hydrogène [51].
Les bactériocines sont des protéines bactéricides produites par des bactéries et qui en affectent d’autres. A la différence des antibiotiques, les bactériocines sécrétées par les bactéries agissent généralement uniquement sur les bactéries de leur propre souche ou sur celles de souches similaires [65].
La majorité des bactéries (Gram positif et Gram négatif) sécrètent des bactériocines, malgré l’importante dépense énergétique correspondante. De nombreuses molécules ont été identifiées : la colicine chez Escherichia coli, la brévicine chez Lactobacillus brevis, la mutacine chez Streptococcus mutans. Elles proviennent d’un propeptide qui, clivé, donnera un peptide actif. Les lantibiotiques (lanthionine-containing antibiotic) sont, quant à eux, sécrétés seulement par des bactéries Gram positif. Ils proviennent d’un propeptide qui change de configuration et se clive pour donner le peptide actif.
Un certain nombre de gènes code pour ces protéines : des opérons portés par le chromosome bactérien, des gènes portés par des plasmides, et des gènes portés par des transposons. On distingue quatre différentes classes :
- Classe I : lantibiotiques
Il s’agit de petits peptides hydrophobes. Ils contiennent des acides aminés non usuels : la lantionine, la bêta-méthyllantionine. Il en existe 2 sous-groupes :
▪ Type A : molécules allongées de plus de 34 acides aminés qui agissent au niveau de la membrane
▪ Type B : molécules globulaires de plus de 19 acides aminés qui agissent au niveau des enzymes.
- Classe II
Il s’agit de petites bactériocines dont le poids moléculaire est inférieur à 10 kDa, qui sont thermostables. Cette classe comprend une plus grande variété de structures, d’où la création de plusieurs sous-groupes.
▪ Sous-classe IIa : bactériocines de 27 à 48 acides aminés
▪ Sous-classe IIb : bactériocines ayant besoin de deux peptides pour avoir une activité. Il en existe 2 types :
▪ Type E (enhancing) où la fonction de l’un des deux peptides est d’augmenter l’activité de l’autre
▪ Type S (synergy) où les deux peptides sont
complémentaires.
▪ Sous-classe IIc : bactériocines ne pouvant pas être classées dans les autres sous-classes.
- Classe III
Elle a été créée pour rassembler les bactériocines de taille supérieure à 10 kDa. Elles sont thermolabiles et peuvent être inactivées après traitement à 60°-100°C pendant 10 à 15 minutes. Cette classe contient quatre bactériocines :
▪ L’helvéticine J produite par Lactobacillus helveticus ▪ L’entérolysine A produite par Enterococcus faecium ▪ La zoocine A produite par Streptococcus zooepidemicus ▪ La millérine B porduite par Streptococcus milleri
- Classe IV
Les bactériocines et les lantibiotiques possèdent différents modes d’action. Ils peuvent être des peptides colonisateurs, tueurs, ou encore des peptides « signal ».
▪ Les peptides colonisateurs produisent des bactériocines qui donnent un avantage compétitif par rapport aux autres souches bactériennes. Par exemple, l’expérience de Hilman et al. en 1987 a montré que la colonisation de la cavité buccale par une souche mutée de S. mutans produisant une mutacine persistait pendant 14 ans. Aucune autre souche de S. mutans n’était retrouvée.
▪ Les peptides tueurs du type IIa, IIb et les lantibiotiques du sous-groupe B agissent au niveau de la membrane cytoplasmique des bactéries cibles. Ils forment des pores dans la membrane cytoplasmique, et entraînent des changements dans la perméabilité membranaire. La production d’énergie nécessaire au métabolisme s’arrête, on assiste alors à la mort cellulaire. A concentrations proches de la CMI, les bactériocines tuent plus rapidement que les antibiotiques. Les peptides tueurs de type lantibiotiques, sous-groupe B, peuvent également agir au niveau de la synthèse du peptidoglycane en bloquant le transporteur lipidique principal. Ils interfèrent donc sur l’activité enzymatique.
▪ Les peptides signaux. Certaines bactériocines joueraient un rôle dual avec une action antibactérienne à haute concentration, et un rôle de peptide signal à basse concentration : un signal qui induit sa propre biosynthèse dans la bactérie, et un signal de communication inter espèces.
En santé bucco-dentaire, Streptococcus salivarius K12, reconnue comme une souche productrice de deux lantibiotiques, aurait une action sur les caries dentaires et les pharyngites à streptocoques, sans perturber l’écosystème buccal.
D’autres molécules interviennent dans ces phénomènes de compétition. La production de peroxyde d’hydrogène par les streptocoques est bien connue in vitro pour inhiber la croissance de bactéries d’autres espèces. Entre autre, S. sanguinis produirait du peroxyde d’hydrogène qui contribuerait à l’antagonisme contre S. mutans [64].
Les acides gras à chaine courte, comme l’acide lactique, issus de la fermentation des hydrates de carbone peuvent également être des inhibiteurs de croissance [64].
Quorum-sensing
Le quorum-sensing est la sécrétion auto-induite d’une ou plusieurs molécules signal très spécifiques en réponse aux changements locaux de densité cellulaire et d’environnement et qui mène à la régulation de l’expression de gènes chez les bactéries. Cela conduit à l’adaptation de nombreuses fonctions comme l’adhésion bactérienne, la production de matrice extracellulaire, la tolérance acide, etc. Il s’agit d’une cible de la thérapie antibactérienne. En effet, un grand nombre de bactéries l’utilise pour contrôler leurs facteurs de virulence. Le blocage de ce phénomène entraînerait donc le blocage de la virulence.
Les cellules planctoniques n’expriment pas ces régulations de leurs fonctions physiologiques [65]. Il a été suggéré que ce phénomène jouerait un rôle possible dans le contrôle des biofilms pour la première fois en 1995.
Un des systèmes de quorum-sensing implique des molécules appelées « autoinducers ». Elles peuvent passer à travers la paroi et la membrane bactériennes. Elles sont produites en phase de croissance bactérienne. Elles diffusent librement dans le milieu : pour les bactéries planctoniques, les molécules sont diluées dans le milieu environnant, et pour les bactéries localisées dans un biofilm, elles sont concentrées dans ce dernier.
Il existe différents types de molécules selon les espèces bactériennes.
Les autoinducers 1 (AI-1) sont spécifiques des bactéries génétiquement identiques [6] et, ce malgré la diversité de l’écosystème buccal.
Les auto-inducers 2 (AI-2) sont considérés comme des signaux universels qui assurent la communication entre des espèces différentes [6]. Ce sont les seuls médiateurs sécrétés aussi bien par les bactéries à Gram positif que négatif. [68] Les AI-2 contrôlent la formation des biofilms pluri-espèces en lien avec la densité bactérienne [6][69] et la régulation de l’expression de facteurs de virulence [68]. AI-2 a été suggérée comme molécule signal potentielle entre les espèces très différentes qui composent le biofilm dentaire.
Les Acyl Homosérines Lactone (AHL) sont produites par beaucoup de bactéries différentes. Elles sont notamment retrouvées chez de nombreuses bactéries Gram négatif telles que Vibrio, Pseudomonas, Escherichia coli. La longueur de leur chaîne est différente selon les espèces (on retrouve de 4 à 18 carbones). Elles activent un facteur transcriptionnel, la protéine R. Il existe une spécificité entre les AHL et la protéine R. Cependant, certaines protéines R peuvent être sensibles à des différentes molécules d’AHL.
A faible densité, les molécules d’AHL sont activement transportées du milieu extérieur vers le cytoplasme par un processus de transport dépendant de l’ATP.
A forte concentration, le transport s’effectue par diffusion passive. Lorsque la concentration d’AHL atteint un seuil appelé le Quorum State, les molécules d’AHL interagissent avec la protéine régulatrice R. Le complexe se lie alors au promoteur des gènes cibles et initie leur transcription.
Les polypeptides courts sont retrouvés chez de nombreuses bactéries Gram positif. Ils sont issus des pré-peptides synthétisés au niveau des ribosomes : parmi eux on trouve les Competence-Stimulating Peptides (CSP) qui sont d’autres médiateurs de la communication cellule à cellule. Les CSP seraient produits par de nombreuses espèces de streptocoques [64]. Selon la revue de la littérature de Guo et al. [70], ils modifieraient la production de bactériocines, la virulence, l’autolyse, la résistance au stress et la formation du biofilm via la modification de la transcription des gènes et de la synthèse des protéines [69]. De plus, les CSP accroissent les compétences génétiques par l’augmentation du transport d’ADN exogène à l’intérieur de la cellule [64].
Pour exemple, en ce qui concerne Streptococcus pneumoniae, la molécule signal CSP est un peptide de 17 acides aminés, hautement spécifique. Il provient d’un pré-peptide appelé ComC, qui est pris en charge par un transporteur, ComAB. ComAB reconnaît spécifiquement ComC et clive son site glycine-glycine. Le CSP est alors fonctionnel. A partir d’un certain seuil de concentration, le CSP se lie à son récepteur, qui est une histidine kinase, appelée ComD, qui fait partie d’un complexe ComDE. On obtient alors la phosphorylation de ComE, et l’activation d’environ 190 gènes.
Ainsi, Welch et al. ont proposé un modèle radial d’organisation du biofilm dentaire basé sur les interactions entre les micro-organismes. Dans ce modèle, entre autres, les bactéries anaérobies s’établiraient vers l’intérieur alors que les aérobies strictes et aérobies facultatives se situeraient plus en périphérie, et les producteurs et consommateurs d’un même métabolite seraient retrouvés à proximité les uns des autres [71].
Pouvoir pathogène du biofilm dentaire
Le biofilm repose sur une organisation complexe relativement stable dans le temps malgré les variations environnementales. Cependant, lorsque cet équilibre se rompt, certaines espèces potentiellement pathogènes prolifèrent au détriment d’autres et il peut y avoir, dans un contexte environnemental particulier, l’apparition de pathologies comme les maladies carieuses et parodontales qui représentent les principales pathologies orales [72].
L’étiologie bactérienne de ces maladies a donc un caractère polymicrobien de par la diversité des bactéries qui agissent sur la destruction tissulaire, mais aussi par les interactions bactériennes qui vont dans le sens de leur survie.
Cette théorie concernant les modalités du pouvoir pathogène du biofilm dentaire est appelée Théorie écologique de la plaque de Marsh. Il s’agit de la théorie la plus actuelle selon laquelle le pouvoir pathogène de la plaque proviendrait d’une dysbiose, un déséquilibre entre les espèces non pathogènes et les espèces pathogènes. Un biofilm dit « sain » présente une composition microbienne très complexe, comportant plus de 100 espèces bactériennes différentes. Parmi ces espèces, trois semblent jouer un rôle important, Streptococcus sanguinis, Streptococcus salivarius, et Veillonella sp.
En effet, ces bactéries ont une action sur les bactéries productrices d’acides potentiellement cariogènes. S. sanguinis et S. salivarius produisent une arginine désaminase et une uréase, qui forme de l’urée et de l’ammonium, et augmente le pH au sein du biofilm. Veillonella utilise le lactate formé par les bactéries fermentaires.
Au contraire, un biofilm « cariogène » a une composition microbienne plus simple, où l’on retrouve majoritairement des Streptococcus mutans, Lactobacillus sp. et Actinomyces sp.
Il existe des streptocoques oraux antagonistes de S. mutans : une étude de Becker et al. sur la carie précoce a démontré un taux de S. mutans très élevé chez les enfants atteints de caries dentaire, alors qu’un taux de S. sanguinis très important était retrouvé chez les enfants exempts de caries [73].
Des interactions antagonistes entre S. mutans et S. sanguinis ont été décrites. Elles entraînent des compétitions nutritionnelles (notons que S. mutans est mieux armé), des pressions environnementales, notamment au niveau du pH, et la production de bactériocines ainsi que de peroxyde d’hydrogène. Le même type d’interaction a également été démontré entre S. mutans et S. gordonii (S. gordonii sécrète une protéase capable de détruire le CSP de S. mutans), et entre S. mutans et S. oligofermentans (ce dernier étant capable d’utiliser l’acide lactique produit par S. mutans, et de produire du peroxyde d’hydrogène). Il contrebalance donc l’action cariogène de S. mutans.
➢ Espèces bactériennes cariogènes
Il existe des critères de cariogénicité, décrits par Newbrun en 1983. Un organisme est défini comme cariogène s’il montre un tropisme pour les dents, s’il est acidogène, s’il est acidurique, et s’il sait utiliser le saccharose.
- Tropisme pour les dents : la capacité d’adhérence provient des fimbriae, des adhésines et des glucanes. En présence des glycosyltransférases, le saccharose est transformé en glucane, soluble ou insoluble. Le mélange de glycosyltransférases entraîne la synthèse de glucanes insolubles adhésifs. Les bactéries en cause s’accumulent donc à la surface des dents, et consolident la plaque.
- Pouvoir acidogène : un organisme acidogène diminue le pH en-dessous de 5,5. Il amène la dissolution des phosphates de calcium (hydroxyapatite) de l’émail, puis de la dentine.
- Production d’acides et fermentation : la fermentation homolactique transforme 1 glucose en 2 acides lactiques. La fermentation hétérolactique transforme 1 glucose en 1 acide lactique + 1 CO2 + 1 acide acétique. Une atmosphère acide inhibe la croissance des
bactéries dites « bénéfiques » et promeut la croissance des bactéries aciduriques.
L’implication des bactéries dans la formation de lésions carieuses a été démontrée par Miller en 1890 [74]. La carie est considérée comme une infection opportuniste causée par les bactéries de la flore commensale orale dans un contexte environnemental particulier [6].
Les études montrent que la carie est associée à une augmentation des proportions des streptocoques du groupe mutans, des lactobacilles et des Actinomyces au sein du biofilm [75]. Ces bactéries sont reconnues comme étant particulièrement acidogènes et acido-tolérantes [6]. En effet, leur potentiel cariogène est conditionné par leur capacité à synthétiser des acides à fort pouvoir de dissolution des phosphates de calcium amélaires à partir de glucides fermentescibles dans des conditions d’anaérobiose en créant localement un pH bas, mais aussi elles doivent pouvoir survivre et croître en milieu acide [8][75][76].
Les streptocoques sont des cocci à Gram positif majoritairement anaérobies facultatifs, non mobiles et non capsulés de diamètre compris entre 0,5 et 1 µm. Ils peuvent être isolés, en diplocoques ou en chaînettes de 5 à 20 éléments. Leur métabolisme fermentatif et leur activité catalase négative les différencient des staphylocoques. Certaines souches possèdent un polysaccharide pariétal antigénique, le polyoside C. Ils sont dits groupables car on peut les classer en 19 sérogroupes différents. Ceux qui ne possèdent pas de polyoside C sont dits « non groupables » par la méthode de Lancefield.
Ils sont responsables de plus de 50% des endocardites infectieuses. De même, la vaste majorité des streptocoques commensaux de la cavité buccale sont non groupables.
Par ailleurs, contrairement aux streptocoques qui ont un pouvoir hémolytique total sur gélose au sang, dits bêta-hémolytiques, les commensaux de la cavité buccale sont alpha-hémolytiques ou viridans, car ils provoquent une hémolyse partielle qui entoure leurs colonies d’un halo vert. La distribution des différentes espèces de streptocoques buccaux est liée à leur capacité d’adhérence aux différentes surfaces.
Les streptocoques du groupe mutans représentent en réalité 7 espèces proches de Streptococcus mutans [77]. S. mutans est la principale bactérie impliquée dans les premiers stades de développement des caries. Elle est présente aussi dans la dentine cariée. Elle possède le potentiel cariogène le plus important parmi les bactéries de la plaque dentaire [76]. Il a été observé que les individus présentant un plus grand nombre de lésions carieuses présentent une quantité de S. mutans plus élevée. Aussi, l’apparition de lésions est précédée 6 à 24 mois plus tôt d’une augmentation de la quantité de S. mutans [78].
Plusieurs facteurs peuvent influencer la colonisation de S. mutans : - Facteur bactérien : par la transmission,
- Facteur biofilm : S. mutans présente une faible compétition avec les autres espèces,
- Facteurs de l’hôte : avec l’hérédité. En effet, l’hôte peut présenter des gènes HLA en rapport avec une mauvaise immunité muqueuse,
- Facteur salive : avec la diminution de la quantité et de la qualité de la salive, - Facteur hygiène : avec l’absence d’hygiène buccale,
- Facteur alimentaire : avec l’ingestion fréquente de snacks et de boissons sucrées. D’autres streptocoques, parmi lesquels Streptococcus sobrinus, Streptococcus mitis, Streptococcus salivarius, présentent eux aussi un potentiel cariogène mais moindre que celui de Streptococcus mutans [76].
A l’instar de Streptococcus mutans, Streptococcus oralis est un groupe de 5 espèces de streptocoques dont l’espèce S. oralis.
Celle-ci fait partie des toutes premières espèces à coloniser la cavité buccale de l’enfant. Elle est aussi pionnière dans la formation de la plaque dentaire au sein de laquelle ces bactéries abondent.
Les lactobacilles sont plutôt responsables de l’évolution des lésions. Ce sont des colonisateurs secondaires des lésions carieuses. Ces bactéries à Gram positif, anaérobies facultatifs de la famille des Lactobacillaceae, sont immobiles, non flagellées, et non sporogènes [79].
Les lactobacilles possèdent un métabolisme fermentaire qui peut être homolactique (production d’'acide lactique à partir du glucose) ou hétérolactique (production d’une quantité équimolaire d'acide lactique, de dioxyde de carbone, d'éthanol et/ou d'acide acétique).
Ils ont une activité catalase ou pseudo-catalase et se développent en ayant recours à des besoins nutritionnels complexes et divers (milieu riche en glucides, acides aminés, peptides, lipides, sels, vitamines).
Les Actinomyces peuvent être retrouvés au niveau des caries radiculaires [6]. Ce sont des bacilles à Gram positif non sporulés anaérobies facultatifs. A. viscosus est très polymorphe se présentant volontiers sous forme de coccobacilles pléiomorphes voire sous forme de filaments ramifiés. Hormis A. naeslundii et A. viscosus, ils sont tous catalase négatif.
A. viscosus est une espèce indigène de la cavité buccale. Il est retrouvé tant à la surface des dents que dans la salive, sur la langue, dans les cryptes amygdaliennes, les surfaces muqueuses et le sillon gingivo-dentaire. C’est un membre important de la plaque dentaire car en permettant la fixation à la Pellicule Acquise Exogène puis la fixation à leur surface de nombreuses autres espèces bactériennes, ils contribuent à assurer une diversité bactérienne au sein de la plaque. Il est retrouvé en abondance lors d’apparition de gingivites et de parodontites débutantes et en proportion moindre dans le fond des poches parodontales profondes. Ainsi, il serait un responsable étiologique dans la maladie parodontale favorisant la transition d’un état sain vers la maladie. Il participe aussi à des infections mixtes avec formation d’abcès, diffusion loco-régionale (cellulite, actinomycose cervico-faciale) et parfois bactériémie avec dissémination sanguine.
➢ Espèces bactériennes parodontopathogènes
Le concept actuel de l’étiologie des maladies parodontales repose sur un complexe bactérien, des facteurs environnementaux et la réponse immunitaire de l’hôte [80].
Socransky et al. ont énoncé en 1977 les critères définissant une bactérie comme parodontopathogène. La bactérie doit être retrouvée en quantité importante au niveau des zones atteintes, entraîner une réponse immunitaire de l’hôte, produire des facteurs de virulence, induire des lésions quand elle est introduite dans un modèle animal, et enfin son élimination doit conduire à une amélioration de la situation clinique [80].
Les bactéries à l’origine des infections parodontales font partie de la flore commensale de la cavité buccale [80]. Mais, avec l’accumulation de la plaque dentaire, la composition du biofilm sain habituellement dominée par les bactéries à Gram positif évolue et la proportion des bactéries anaérobies à Gram négatif devient de plus en plus importante [64].
![Tableau 1 : Liste non exhaustive de biofilms in vitro les plus couramment employés [83]](https://thumb-eu.123doks.com/thumbv2/123doknet/14739692.754861/39.892.86.811.112.1056/tableau-liste-exhaustive-biofilms-in-couramment-employés.webp)
![Figure 2 : Schéma résumant le déroulement de la procédure expérimentale utilisée pour étudier la formation des biofilms selon le modèle de Zurich [87]](https://thumb-eu.123doks.com/thumbv2/123doknet/14739692.754861/43.892.101.772.107.592/schéma-résumant-déroulement-procédure-expérimentale-utilisée-étudier-formation.webp)