HAL Id: insu-02317387
https://hal-insu.archives-ouvertes.fr/insu-02317387
Submitted on 19 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Distributed under a Creative Commons Attribution - NonCommercial - NoDerivatives| 4.0
International License
A new species of Lapparentophis from the
mid-Cretaceous Kem Kem beds, Morocco, with remarks
on the distribution of lapparentophiid snakes
Romain Vullo
To cite this version:
Romain Vullo. A new species of Lapparentophis from the mid-Cretaceous Kem Kem beds, Morocco,
with remarks on the distribution of lapparentophiid snakes. Comptes Rendus Palevol, Elsevier Masson,
2019, 18 (7), pp.765-770. �10.1016/j.crpv.2019.08.004�. �insu-02317387�
ContentslistsavailableatScienceDirect
Comptes
Rendus
Palevol
www . s c ie n c e d i r e c t . c o m
General
Palaeontology,
Systematics,
and
Evolution
(Vertebrate
Palaeontology)
A
new
species
of
Lapparentophis
from
the
mid-Cretaceous
Kem
Kem
beds,
Morocco,
with
remarks
on
the
distribution
of
lapparentophiid
snakes
Une
nouvelle
espèce
de
Lapparentophis
du
Crétacé
moyen
des
Kem
Kem,
Maroc,
et
remarques
sur
la
distribution
des
serpents
lapparentophiidés
Romain
Vullo
UniversitédeRennes,CNRS,GéosciencesRennes,UMR6118,35000Rennes,France
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received27February2019
Acceptedafterrevision29August2019 Availableonline10October2019 HandledbyNathalieBardet Keywords: Squamata Ophidia Lapparentophiidae Albian Cenomanian NorthAfrica Palaeobiogeography
a
b
s
t
r
a
c
t
Twoisolatedtrunkvertebraefromthe?uppermostAlbian–lowerCenomanianKemKem bedsofMoroccoaredescribedandassignedtoLapparentophis,anearlysnakegenusknown fromcoevaldepositsinAlgeria.TheMoroccanspecimensrepresentanewspecies, Lap-parentophisragei,whichcanbe distinguishedfromthetypeandonlyknownspecies, Lapparentophisdefrennei,byitssmallersize,itsmoreelongatevertebrae,thepresenceof parazygosphenalforamina,andparadiapophysesextendinganteroventrallyclosertothe cotyle.ThediscoveryofLapparentophisintheKemKembedsaddstotherelativelydiverse snakeassemblagepreviouslyreportedfromthisformationandextendsthegeographical rangeofthegenus.ThedistributionofLapparentophisandlapparentophiid-grade (?lap-parentophiid)snakesisdiscussed.Thispoorlyknownfamilyofterrestrialsnakesseemsto berestrictedtothelatestAlbian–earlyCenomanianofNorthAfrica,withtheexceptionof Pouitellafromtheearly–middleCenomanianofFrance.Asformanyothervertebratetaxaof thisperiod,thisdistributionisconsistentwithadispersaleventfromAfricatothewestern partoftheEuropeanarchipelago.
©2019Acad ´emiedessciences.PublishedbyElsevierMassonSAS.Thisisanopenaccess articleundertheCCBY-NC-NDlicense(http://creativecommons.org/licenses/by-nc-nd/4. 0/). Motsclés: Squamata Ophidia Lapparentophiidae Albien Cénomanien AfriqueduNord Paléobiogéographie
r
é
s
u
m
é
Deuxvertèbresdorsalesprovenantdel’Albienterminal–CénomanieninférieurdesKem Kem(Maroc)sontdécritesetattribuéesàLapparentophis,unserpentbasalconnudans desdépôtséquivalentsenAlgérie.Lesspécimensmarocainsreprésententunenouvelle espèce, Lapparentophis ragei, qui diffère dela seule autre espèce connue, Lapparen-tophis defrennei,parsapluspetite taille,sesvertèbresplusallongées,la présencede foramensparazygosphéniensetdesparadiapophysesplusrapprochées antéroventrale-mentducotyle.LadécouvertedeLapparentophisdanslesKemKemaccroîtladiversitéde l’assemblagedeserpentsdéjàdécritdanscesniveauxetétendladistributiongéographique dugenre.LadistributiondeLapparentophisetdesserpentsdegradelapparentophiidés (?Lapparentophiidae)estdiscutée.Cettefamilledeserpentsterrestres,quidemeuremal connue,sembleêtrerestreinteàl’Albienterminal–Cénomanieninférieurd’AfriqueduNord,
E-mailaddress:romain.vullo@univ-rennes1.fr https://doi.org/10.1016/j.crpv.2019.08.004
1631-0683/©2019Acad ´emiedessciences.PublishedbyElsevierMassonSAS.ThisisanopenaccessarticleundertheCCBY-NC-NDlicense(http:// creativecommons.org/licenses/by-nc-nd/4.0/).
766 R.Vullo/C.R.Palevol18(2019)765–770
àl’exceptiondePouitelladuCénomanieninférieur–moyendeFrance.Commepourde nom-breuxautresgroupesdevertébrés,cettedistributionestenaccordavecl’existenced’une migrationdepuisl’Afriqueverslapartieoccidentaledel’archipeleuropéen.
©2019Acad ´emiedessciences.Publi ´eparElsevierMassonSAS.Cetarticleestpubli ´een OpenAccesssouslicenceCCBY-NC-ND(http://creativecommons.org/licenses/by-nc-nd/ 4.0/).
1. Introduction
Althoughprimitivesnakeshavebeenreportedasearly astheMiddleJurassicbasedonafewjawfragmentsfrom theBathonianofEngland(Caldwelletal.,2015),the mid-dle partof the Cretaceous coincides with an explosive increaseinthesnakefossilrecordworldwide(e.g.,Albino
etal.,2016;ApesteguíaandZaher,2006;CaldwellandLee,
1997; Caldwellet al., 2015; Cuny et al., 1990; Gardner
andCifelli,1999;Hsiouetal.,2014;Kleinetal.,2017;Lee
etal., 1999;Rage, 1988;Rage and Escuillié, 2000;Rage
andWerner,1999;Rageetal.,2016;Tchernovetal.,2000;
Xingetal.,2018).Snakesareparticularlywellrepresented inthe vertebrate assemblagesfrom themid-Cretaceous (Albian–Cenomanian)ofNorthAfrica,withvarious,mostly terrestrial forms described from Morocco (Klein et al.,
2017;RageandDutheil,2008),Algeria(Cunyetal.,1990;
Hoffstetter,1959), Libya (Nessovet al., 1998; Rage and
Cappetta,2002),Sudan(RageandWerner,1999;Werner
and Rage, 1994), and Egypt (Nopcsa, 1925).The genus
Lapparentophis(and the family Lapparentophiidae) was recognised based on three isolated vertebrae found in depositsoflateAlbian–CenomanianageatInAkhamilin easternAlgeria(Hoffstetter,1959).Sofar,Lapparentophis isonlyknownfromthetypelocalityandincludesa sin-glespecies, L. defrennei. In addition,a few vertebraeof possiblelapparentophiidsnakeshavebeendescribedfrom coevaldepositsinnorthernAlgeria(Cunyetal.,1990)and Sudan(RageandWerner,1999;WernerandRage,1994). Outside Africa, thepoorly known genusPouitella is the onlyothertaxontentativelyreferredtoLapparentophiidae
(Rage,1988).HereIreportthefirstoccurrenceof
Lappar-entophisfromtherichanddiversevertebrateassemblage oftheMoroccanKemKembeds.Thematerial,consisting oftwowell-preservedandisolatedvertebrae,representsa newspecies.
2. Geologicalandpalaeontologicalsettings
ThetwospecimenscomefromthelowerpartoftheKem KembedscroppingoutatElBegâa,aproductivefossilsite locatedafew km eastofTaouz (southeasternMorocco)
(seeCavinetal.,2010).ThelowerpartoftheKemKem
bedsliewithintheIfezouaneFormation, whichis com-posedmainlyofsandstonecharacterizedbycross-stratified structuresand channel structures filled with microcon-glomerates (Cavin et al., 2010). These beds are ?latest AlbiantoearlyCenomanianinageandweredepositedin fluvialanddeltaicenvironments(Cavinetal.,2010).The IfezouaneFormationisrenownedforitsrichanddiverse vertebrateassemblage includingselachians,bonyfishes,
lissamphibians,turtles,squamates,pterosaurs, crocodyli-formsanddinosaurs(seeCavinetal.,2010).Thesquamate assemblage consistsmostlyof snakes, includingvarious forms such as Simoliophis cf. libycus, Norisophis begaa, an indeterminate madtsoiid, a possiblenigerophiid and an incertae sedis snake (Klein et al., 2017; Rage and
Dutheil, 2008).Inaddition,three lizardspecimens have
beenreportedsofarfromtheKemKemBeds, correspond-ingtoanisolatedvertebraofanindeterminateform(Rage
andDutheil,2008),ajawfragmentassignedtoan
acrodon-tan(Jeddaherdanaleadonta;Apesteguíaetal.,2016), and adentary ofaborioteiioid(Bicuspidonhogreli;Vulloand
Rage,2018). 3. Systematicpalaeontology LepidosauriaHaeckel,1856 SquamataOppel,1811 OphidiaBrongniart,1800 LapparentophiidaeHoffstetter,1959 LapparentophisHoffstetter,1959
Type species. Lapparentophis defrennei Hoffstetter,
1959.
Lapparentophisrageisp.nov.
Etymology.Thespecificepithetisinhonourofmylate friendandcolleagueJean-ClaudeRage,whofirstsuggested that thematerialstudiedheremaybelongto Lapparen-tophis,andinrecognitionofhisinnumerableandinvaluable contributionstopalaeoherpetologyand palaeobiogeogra-phy.
Type material. MHNM.KK387 (holotype) and
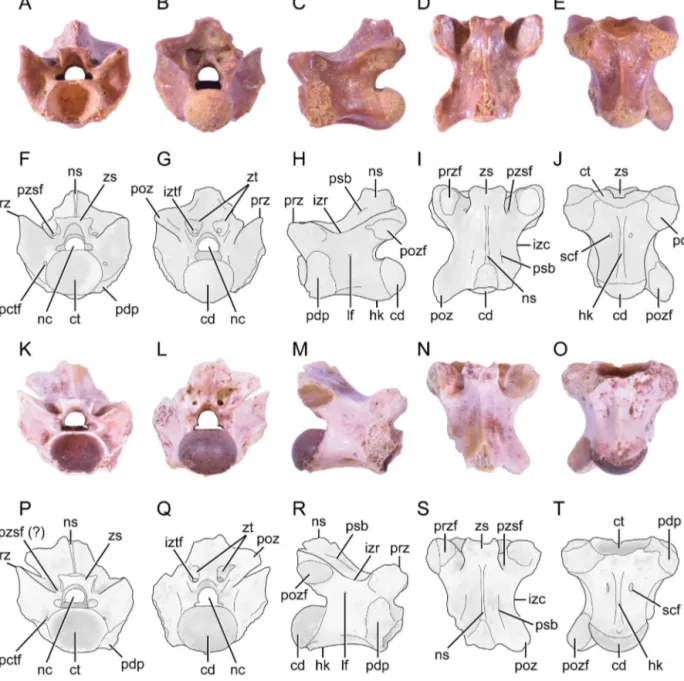
MHNM.KK388 (paratype), two nearly complete trunk vertebrae (Fig. 1) housed in the “Muséum d’Histoire NaturelledeMarrakech”(MHNM),Morocco.
Typelocalityandhorizon.ElBegâalocalitynearTaouz, southeasternMorocco;KemKembeds,Ifezouane Forma-tion,?latest Albian–earlyCenomanianin age(seeCavin
etal.,2010).
Diagnosis. Aspecies ofLapparentophisdifferingfrom thetype and onlyotherspecies, L.defrennei,in the fol-lowing features:smallersize,more elongate(aslongas wide) vertebrae,presenceof parazygosphenalforamina, paradiapophyses (synapophyses) extending anteroven-trallyclosertothecotyle.
Description. Both specimens are equally-sized, non-pachyostoticmid-trunkvertebraethatcomefromasimilar position.Thetipoftheneuralspineisbrokenonboth spec-imens.Severalfeatures(i.e.,robustgeneralmorphology, centrummoderatelyelongate,neuralcanalsmallerthan cotyleandcondyle,prezygapophyseswelldeveloped) sug-gestthatbothvertebraearefromadults(e.g.,Gardnerand
Fig.1. PhotographsandinterpretativelinedrawingsoftrunkvertebraeofLapparentophisrageisp.nov.,holotypeMHNM.KK387(A–J)andparatype MHNM.KK388(K–T)inanterior(A,F,K,P),posterior(B,G,L,Q),leftlateral(C,H),rightlateral(M,R),dorsal(D,I,N,S),andventral(E,J,O,T)views. Abbreviations:cd:condyle;ct:cotyle;hk:haemalkeel;izc:interzygapophysealconstriction;izr:interzygapophysealridge;iztf:intrazygantralforamen; lf:lateralforamen;nc:neuralcanal;ns:neuralspine;pctf:paracotylarforamen;pdp:paradiapophysis;poz:postzygapophysis;pozf: postzygapophy-sealarticularfacet;prz:prezygapophysis;przf:prezygapophysealarticularfacet;psb:parasagittalbulge;pzsf:parazygosphenalforamen;scf:subcentral foramen;zs:zygosphene;zt:zygantrum.Scalebar:5mm.
Fig.1. Photographies etdessinsinterprétatifsdesvertèbresdorsalesde Lapparentophisrageisp.nov.,holotypeMHNM.KK387(A–J)etparatype MHNM.KK388(K–T)envuesantérieure(A,F,K,P),postérieure(B,G,L,Q),latéralegauche(C,H),latéraledroite(M,R),dorsale(D,I,N,S)etventrale (E,J,O,T).Abréviations:cd:condyle;ct:cotyle;hk:carènehémale;izc:constrictioninterzygapophysaire;izr:rideinterzygapophysaire;iztf:foramen intrazygantral;lf:foramenlatéral;nc:canalneural;ns:épineneurale;pctf:foramenparacotylien;pdp:paradiapophyse;poz:postzygapophyse; pozf:facettearticulairepostzygapophysaire;prz:prézygapophyse;przf:facettearticulaireprézygapophysaire;psb:renflementparasagittal;pzsf: foramenparazygosphénien;scf:foramensubcentral;zs:zygosphène;zt:zygantrum.Barred’échelle:5mm.
Cifelli,1999).Thedescriptionprovidedhereismainlybased
ontheholotype(Fig.1A–J);theparatype(Fig.1K–T) gener-allydisplaysthesamefeatures,unlessindicatedotherwise. Measurementsofbothvertebralspecimensareprovidedin
Table1.
In anterior view (Fig. 1A, F, K, P),the type vertebra showsnodorsoventraldepression.Thecotyleisslightly widerthanhigh.Thezygospheneisrelativelysmall,clearly narrowerthanthecotyle;itshowsaslightlyconvexdorsal marginandtwolateralbulges.Theroofofthezygosphene
768 R.Vullo/C.R.Palevol18(2019)765–770
Table1
Measurements(inmm)forvertebraeofLapparentophisrageisp.nov.
Tableau1
Mesures(enmm)desvertèbresdeLapparentophisrageisp.nov.
HolotypeMHNM.KK387 ParatypeMHNM.KK388
Maximumlengthofvertebra 10.7 10.2
Centrumlength(fromcotylarrimtoposteriormostendofcondyle) 8.3 8.4
Cotyleheight 4.1 4.2
Cotylewidth 5.5 5.6
Condyleheight 3.7 3.9
Condylewidth 4.3 5.1
Widthacrossprezygapophyses 10.1 10.6
Widthofinterzygapophysealconstriction 5.8 6.2
Zygosphenewidth 3.5 3.5
isthickerthanthecotylarrim.Thezygosphenalfacetsare oblique.Theneuralcanalistrifoliateandwiderthanhigh; itismarkedlysmallerthanthecotyle.Thelateralmargins ofthehighprezygapophyseal buttressesaresubvertical andslightlyconcave.Theprezygapophysealarticularfacets aremarkedlyoblique,extendinglaterodorsallyat40◦from horizontal;theirmedialendsarepositionedatthelevel ofthedorsoventralmidpointoftheopeningfortheneural canalandtheirdistalendsliejustabovethelevelofthe zygosphenalroof.Paracotylarandsmallparazygosphenal foraminaarepresent intheholotype; however, parazy-gosphenalforaminaseemtobeabsent(orminute?)inthe paratype.Theparadiapophysesextenddorsallyjustabove thelevelofthedorsalrimofthecotyleandanteroventrally lieclose tothecotylar rim(i.e., noparacotylar notches are present). The neural arch is relatively narrow and moderatelyelevated.Posteriorly,theneuralspineiswell separatedfromtheroofoftheneuralarch.
Inposteriorview(Fig.1B,G,L,Q),thecondyleisslightly widerthanhigh.Theneuralcanal,markedlylesstrifoliate that in anteriorview, is higher thanwide; it is notice-ablysmallerthanthecondyle.Ashallowmedianridgeis presentontheflooroftheneuralcanal.Thezygantrumis slightlynarrowerthanthecondyle;itshowsoblique, well-developedarticularfacetsandapairoflargeintrazygantral foramina.Therearenoparazygantralforamina.Theneural spineiswelldifferentiatedfromthepostzygapophyses,i.e., theposteriormarginoftheneuralarchbetweentheneural spineandeachpostzygapophysisismarkedlyconcave.
Inlateralview(Fig.1C,H,M,R),thevertebraappears ratherelongate,witha moderatelyelevatedneuralarch. The neural spine is restricted to the posterior half of theneuralarch;itsobliqueanteriorborderrisessteeply. The paradiapophyses are large; they are anteroposteri-orlybroadanddorsoventrallytall.Thejunctionbetween the diapophyseal and parapophyseal articular facets is markedbyaslightconstriction;theparapophysealfacet islargerthanthediapophysealfacet.The interzygapophy-seal ridgeis salient and dorsallyconcave; small lateral foramina are present ventral to this ridge. The axis of the cotyle–condyle system is slightly oblique (i.e., the cotyleandthecondyleareorientedslightlydownwardand slightlyupward,respectively).Theventralmarginofthe centrumisconcaveandbearsashallowhaemalkeel.
Indorsal view(Fig.1D, I,N, S),the interzygapophy-seal constriction is well marked. The width across the prezygapophyses is slightly greater than across the
postzygapophyses. The neural arch shows a weak parasagittal bulge oneach side ofthe baseof the neu-ralspine.Theposteriormarginoftheneuralarchforms a broadconcavity,withoutadistinctmediannotch.The zygospheneshowsagentlyconcaveanteriormarginanda slightanterolateraltruncation.Theobovate prezygapophy-sealarticularfacetsareacuteanterolaterally.Thereareno prezygapophysealprocesses.
In ventral view (Fig. 1E, J, O, T), the centrum is subtriangular; its lateralmargins are markedly concave lateroposteriorly. Theshallow, ridge-likehaemalkeel is wellmarked;itoriginatesanteriorlyfromnearthe coty-lar rim and extends posteriorly to the condyle, where it forms a smalltubercle.A pairof subcentralforamina is present. The paradiapophysesare large, with antero-posteriorlywell-developedparapophysealarticularfacets reachingthecotylarrim(i.e.,noparacotylarnotchesare present).Thepostzygapophysealarticularfacetsare obo-vate,i.e.,longerthanwide.
4. Discussion
Lapparentophiids are rare mid-Cretaceous terrestrial snakesthatareknownonlyfromafewisolatedvertebrae. Lapparentophisdefrennei (andthefamily Lapparentophi-idae)waserectedonthebasisofthreetrunkvertebraefrom the?uppermostAlbian–CenomanianofInAkhamil,
Alge-ria(Hoffstetter,1959).Thisspeciesissignificantlylarger
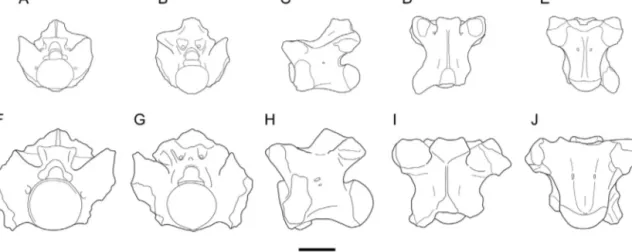
than Lapparentophis ragei and is characterized by more massivelybuilttrunkvertebrae(Fig.2).Are-examination ofthetypematerialofLapparentophisdefrenneihas con-firmed that no parazygosphenal foramina are present, unlikeinL.ragei;inaddition,prezygapophysesaremore extended laterally,withmoredeveloped prezygapophy-sealarticularfacets;theneuralarchbearsawell-developed bulgeposteriortotheprezygapophysealarticularfacets; paracotylarnotchesarepresent;thezygospheneismore truncatedanterolaterally;theinterzygapophysealridgeis lessmarked;thecentrumismorerobust;theaxisofthe cotyle–condylesystemislessoblique;andthehaemalkeel islessmarked(Fig.2).
Threeotherisolatedvertebraefromthemid-Cretaceous ofNorthAfricaweretentativelyassignedto Lapparentophi-idae(oridentifiedasbelongingtolapparentophiid-grade snakes). A single,poorlypreserved vertebra wasbriefly describedfromtheuppermostAlbianofElKohol,Algeria (Cunyetal.,1990).Thisincompletespecimen, consisting
Fig.2. ComparisonoftheholotypetrunkvertebraeofLapparentophisrageisp.nov.(A–E)andL.defrennei(F–J)inanterior(A,F),posterior(B,G),leftlateral (C,H),dorsal(D,I),andventral(E,J)views.LinedrawingsofthevertebraofLapparentophisdefrenneiafterHoffstetter(1959:fig.2a).Scalebar:5mm.
Fig.2. Comparaisondesvertèbresdorsales(holotypes)deLapparentophisrageisp.nov.(A–E)etL.defrennei(F–J)envuesantérieures(A,F),postérieures (B,G),latéralesgauches(C,H),dorsales(D,I),etventrales(E,J).DessinsdelavertèbredeLapparentophisdefrenneid’aprèsHoffstetter(1959:fig.2a).Barre d’échelle:5mm.
only of the centrum,is about half thesize of the Kem Kem specimens described here. In ventral view, it is similarinshape tothecentrumofLapparentophisragei; however, in the El Kohol vertebra, the lateral margins of the centrum are more inflated and the haemal keel is wider and less sharp (Cuny et al., 1990: fig. 1). In addition, two isolated vertebrae representing two dis-tinct?Lapparentophiidae(“lapparentophiid-gradesnakes AandB”)weredescribedfromtheCenomanianofWadi Abu Hashim, Sudan (Rage and Werner, 1999; Werner
andRage,1994).Thelapparentophiid-gradesnakeAfrom
Wadi Abu Hashim, which may represent a new genus and species according to Rage and Werner (1999), is known from a single small trunk vertebra, wider than long.ThisformissimilarinsizetoLapparentophisragei; however, it canbe distinguished fromthe latter by its less elongatevertebrae,itshigherneuralarch,itswider and blunterhaemalkeel,andtheabsenceofboth para-cotylarandparazygosphenalforamina(RageandWerner, 1999: fig. 2). The lapparentophiid-grade snake B from WadiAbuHashimisonlyrepresentedbyasmall(2.7mm long),elongatecaudalvertebraand,therefore,cannotbe compared witheither of the Kem Kem trunk vertebral specimens. The generally accepted Cenomanian age of the WadiMilk Formation, which yielded a rich assem-blageofsnakes(includingthetwolapparentophiid-grade forms A and B discussed above) and other vertebrates atWadiAbuHashim(Rauhut,1999;Werner,1994),has recently been challenged based on palynological data
(Eisawi,2015)anddetritalzircongeochronology(Owusu
Agyemang et al., 2019).These two studies suggested a
Campanian–Maastrichtian age for this formation. How-ever,thepalynological datapresented byEisawi (2015)
arefromtheShendiFormation,asupposedlateral equiva-lentoftheWadiMilkFormation,andthusdonotprovide a direct age estimate for the Wadi Abu Hashim verte-brateassemblage.Similarly,thesixLateCretaceouszircon grainsreportedbyOwusuAgyemangetal.(2019)come
fromthe Jebelel Gamman region, whileno Cretaceous zirconswerefoundintheWadiAbuHashimregion.In addi-tion,thefivepotentiallyage-constraininggrainssuggesting a maximum depositional age of 79.2±2.4 Ma (middle Campanian)comefromfoursandstonesamplesthat cor-respondtofourdistincthorizonsdistributedthroughthe upperand lower membersof theWadiMilkFormation
(Owusu Agyemanget al., 2019: figs. 3 and 7).
Accord-ingtoDickinsonandGehrels(2009)andRossignoletal.
(2019),suchresultsshouldbeconsiderednotstatistically
robust.Furthermore,the occurrenceof somevertebrate taxa(e.g.,distobatidsharks;VulloandNéraudeau,2008) in the WadiAbu Hashim assemblage is not consistent with a post-Cenomanian age. For these reasons, the Sudaneselapparentophiid-gradesnakesdescribedbyRage
andWerner(1999)arestillregardedhereasCenomanian
inage.
Pouitella (type and only known species: P. pervetus
Rage,1988),fromtheearly–middleCenomanianof
west-ernFrance(holotypeandonlyknownspecimenfromBrézé andstillundescribedmaterialfromLussant;seeCunyetal., 1990),isapoorlyknownbasalsnakeshowingageneral morphologyandalevelofevolutionsimilarto Lapparen-tophis(Rage,1988).ThisledRage(1988)toprovisionally placePouitellawithinLapparentophiidae.Thisplacementis herereinforcedbythepresenceofparazygosphenal foram-inainLapparentophisragei,anunusualapomorphicfeature alsoobservedinPouitellapervetus(Rage,1988)and oth-erwiseknownamongsnakesonlyintwoorthreelatest Cretaceousmadtsoiids(Ladukeetal.,2010)andinafew post-Cretaceoustaxa(Head,2005;McCartneyetal.,2018;
Rage,1983,2001,2008;Rageetal.,2003,2008).Pouitella
isthereforeconfirmedasamemberofLapparentophiidae, thusrepresentingtheonlyknownoccurrenceofthisfamily inLaurasia.ThissuggeststhatadispersaleventfromAfrica toEuropemayhaveoccurredinlapparentophiidsnakes, whichisconsistentwiththepresenceofseveralother ver-tebratetaxawithGondwananaffinitiesintheCenomanian
770 R.Vullo/C.R.Palevol18(2019)765–770
ofwesternEurope(seeCsiki-Savaetal.,2015;Vulloand
Rage,2018).
Acknowledgments
IwarmlythankNour-EddineJalilforprovidingMHNM collectionnumbers,DamienGendryforhishelpwiththe photographsofthespecimens,andMarcPoujolfor use-ful discussion on detrital zircon geochronology. I wish to thank Nathalie Bardet for her editorial assistance, andMichaelCaldwell,JamesGardnerandananonymous reviewerfortheirconstructivecommentsthatimproved themanuscript.Finally,Iamdeeplyindebtedtomylate friendandcolleagueJean-ClaudeRageforveryuseful dis-cussions,andforhiscontinuousand mostvaluablehelp inexaminingandinterpretingfossilmaterialfromvarious localitiesduringthelasttwodecades.
References
Albino,A.,Carrillo-Brice ˜no,J.D.,Neenan,J.M.,2016.Anenigmaticaquatic snakefromtheCenomanian ofnorthernSouthAmerica.PeerJ4, e2027.
Apesteguía,S.,Zaher,H.,2006.ACretaceousterrestrialsnakewithrobust hindlimbsandasacrum.Nature440,1037–1040.
Apesteguía,S.,Daza,J.D.,Simões,T.R.,Rage,J.-C.,2016.Thefirstiguanian lizardfromtheMesozoicofAfrica.R.Soc.OpenSci3,160462. Caldwell,M.W.,Lee,M.S.Y.,1997.Asnakewithlegsfromthemarine
Cre-taceousoftheMiddleEast.Nature386,705–710.
Caldwell,M.W.,Nydam,R.L.,Palci,A.,Apesteguía,S.,2015.Theoldest knownsnakesfromtheMiddleJurassic–LowerCretaceousprovide insightsonsnakeevolution.Nat.Comm.6,5996.
Cavin,L.,Tong,H.,Boudad,L.,Meister,C.,Piuz,A.,Tabouelle,J.,Aarab,M., Amiot,R.,Buffetaut,E.,Dyke,G.,Hua,S.,LeLœuff,J.,2010.Vertebrate assemblagesfromtheearlyLateCretaceousofsoutheasternMorocco: anoverview.J.Afr.EarthSci.57,391–412.
Csiki-Sava,Z.,Buffetaut,E., ˝Osi,A.,Pereda-Suberbiola,X.,Brusatte,S.,2015. IslandlifeintheCretaceous–faunalcomposition,biogeography, evo-lution,andextinctionofland-livingvertebratesontheLateCretaceous Europeanarchipelago.Zookeys469,1–161.
Cuny,G.,Jaeger,J.-J.,Mahboubi,M.,Rage,J.-C.,1990.Lesplusanciens ser-pents(Reptilia,Squamata)connus.Miseaupointsurl’âgegéologique desserpentsdelapartiemoyenneduCrétacé.C.R.Acad.Sci.Paris, Sér.II311,1267–1272.
Dickinson,W.R.,Gehrels,G.E.,2009.UseofU–Pbagesofdetritalzirconsto infermaximumdepositionalagesofstrata:atestagainstaColorado PlateauMesozoicdatabase.EarthPlanet.Sci.Lett.288,115–125. Eisawi, A.A.M., 2015. Palynological evidence of a
Campanian–Maastrichtian age of the Shendi Formation (Shendi Basin,centralSudan).Am.J.EarthSci.2,206–210.
Gardner,J.D.,Cifelli,R.L.,1999.AprimitivesnakefromtheCretaceousof Utah.Spec.Pap.Palaeontol.60,87–100.
Head,J.J.,2005. SnakesoftheSiwalik Group(Mioceneof Pakistan): Systematicsandrelationshiptoenvironmentalchange.Palaeontol. Electron.8,18A.
Hoffstetter,R.,1959.UnserpentterrestredansleCrétacéinférieurdu Sahara.Bull.Soc.geol.France,Sér.71,897–902.
Hsiou,A.S.,Albino,A.M.,Medeiros,M.A.,Santos,R.A.B.,2014.Theoldest BraziliansnakesfromtheCenomanian(earlyLateCretaceous).Acta Palaeontol.Pol.59,635–642.
Klein,C.G.,Longrich,N.R.,Ibrahim,N.,Zouhri,S.,2017.Anewbasalsnake fromthemid-CretaceousofMorocco.CretaceousRes.72,134–141. Laduke,T.C.,Krause,D.W.,Scanlon,J.D.,Kley,N.J.,2010.ALateCretaceous
(Maastrichtian)snakeassemblagefromtheMaevaranoFormation, MahajangaBasin,Madagascar.J.Vert.Paleontol.30,109–138. Lee,M.S.Y., Caldwell, M.W., Scanlon, J.D., 1999. A second primitive
marinesnake:PachyophiswoodwardifromtheCretaceousof Bosnia-Herzegovina.J.Zool.248,509–520.
McCartney,J.A.,Roberts,E.M.,Tapanila,L.,O’Leary,M.A.,2018. Large palaeophiidandnigerophiidsnakesfromPaleogeneTrans-Saharan SeawaydepositsofMali.ActaPalaeontol.Pol.63,207–220.
Nessov,L.A.,Zhegallo,V.I.,Averianov,A.O.,1998.AnewlocalityofLate Cretaceoussnakes,mammalsandothervertebratesinAfrica(western Libya).Ann.Paleontol.84,265–274.
Nopcsa,F.,1925.ErgbenissederForschungsreisenProf.E.Stromersin denWüstenÄgyptens,II.Wirbeltier-RestederBaharîje-Stufe (unter-stesCenoman).5.DieSymoliophis-Reste.Abh.Bayerisch.Akad.Wiss. Math.-naturwiss.Abt.30,1–27.
OwusuAgyemang,P.C.,Roberts,E.M.,Bussert,R.,Evans,D.,Müller,J.,2019. U–Pbdetritalzirconconstraintsonthedepositionalageand prove-nanceofthedinosaur-bearingUpperCretaceousWadiMilkFormation ofSudan.CretaceousRes.97,52–72.
Rage,J.-C.,1983.Palaeophiscolossaeusnov.sp.(leplusgrandserpent connu ?)del’Eocènedu Malietle problèmedugenrechezles Palaeopheinae.C.R.Acad.Sci.Paris,Sér.II296,1741–1744. Rage,J.-C.,1988.Unserpentprimitif(Reptilia,Squamata)dansle
Céno-manien(baseduCrétacésupérieur).C.R.Acad.Sci.Paris,Sér.II307, 1027–1032.
Rage,J.-C.,2001.FossilsnakesfromthePalaeoceneofSãoJosédeItaboraí, Brazil.PartII.Boidae.Palaeovertebrata30,111–150.
Rage,J.-C.,2008.FossilsnakesfromthePalaeoceneofSãoJosédeItaboraí, Brazil.PartIII.Ungaliophiinae,booidsincertaesedis,andCaenophidia. Summary,update,anddiscussionofthesnakefaunafromthelocality. Palaeovertebrata30,37–73.
Rage,J.-C.,Cappetta,H.,2002.VertebratesfromtheCenomanian,andthe geologicalageoftheDraaUbariFauna(Libya).Ann.Paleontol.88, 79–84.
Rage, J.-C.,Dutheil, D.B.,2008. Amphibiansandsquamates fromthe Cretaceous (Cenomanian) ofMorocco–A preliminary study, with descriptionofanewgenusofpipidfrog.PalaeontographicaAbt.A 285,1–22.
Rage,J.-C.,Escuillié,F.,2000.UnnouveauserpentbipèdeduCénomanien (Crétacé).Implicationsphylétiques.C.R.Acad.Sci.Paris,Sér.IIa330, 513–520.
Rage,J.-C.,Werner,C.,1999.Mid-Cretaceous(Cenomanian)snakesfrom WadiAbuHashim,Sudan:Theearliestsnakeassemblage.Palaeont. Afr.35,85–110.
Rage,J.-C.,Bajpai,S.,Thewissen,J.G.M.,Tiwari,B.N.,2003.EarlyEocene snakesfromKutch,WesternIndia,withareviewofthePalaeophiidae. Geodiversitas25,695–716.
Rage,J.-C.,Folie,A.,Rana,R.S.,Singh,H.,Rose,K.D.,Smith,T.,2008.A diversesnakefaunafromtheearlyEoceneofVastanLigniteMine, Gujarat,India.ActaPalaeontol.Pol.53,391–403.
Rage,J.-C.,Vullo,R.,Néraudeau,D.,2016.Themid-Cretaceoussnake Simo-liophisrochebruneiSauvage,1880(Squamata:Ophidia)fromitstype area(Charentes,southwesternFrance):Redescription,distribution, andpalaeoecology.CretaceousRes.58,234–253.
Rauhut,O.W.M.,1999.AdinosaurfaunafromtheLateCretaceous (Ceno-manian)ofnorthernSudan.Palaeontol.afr.35,61–84.
Rossignol,C.,Hallot,E.,Bourquin,S.,Poujol,M.,Jolivet,M.,Pellenard,P., Ducassou,C.,Nalpas,T.,Heilbronn,G.,Yu,J.,Dabard,M.-P.,2019.Using volcaniclasticrockstoconstrainsedimentationages:towhatextent arevolcanismandsedimentationsynchronous?Sediment.Geol.381, 46–64.
Tchernov,E.,Rieppel,O.,Zaher,H.,Polcyn,M.J.,Jacobs,L.L.,2000.Afossil snakewithlimbs.Science287,2010–2012.
Vullo, R., Néraudeau, D., 2008. When the “primitive” shark Tribo-dus(Hybodontiformes)meetsthe“modern”rayPseudohypolophus (Rajiformes):theuniqueco-occurrenceofthesetwodurophagous CretaceousselachiansinCharentes(SWFrance).ActaGeol.Pol.58, 249–255.
Vullo,R.,Rage,J.-C.,2018.ThefirstGondwananborioteiioidlizardandthe mid-CretaceousdispersaleventbetweenNorthAmericaandAfrica. Sci.Nat.105,61.
Werner, C.,1994. Die kontinentale Wirbeltierfauna ausder unteren OberkreidedesSudan(WadiMilkFormation).Berlinergeowiss.Abh. E13,221–249.
Werner,C.,Rage,J.-C.,1994.Mid-CretaceoussnakesfromSudan.A pre-liminaryreportonanunexpectedlydiversesnakefauna.C.R.Acad. Sci.Paris,Sér.II319,247–252.
Xing,L.,Caldwell,M.W.,Chen,R.,Nydam,R.L.,Palci,A.,Simões,T.R., McKellar,R.C.,Lee,M.S.Y.,Liu,Y.,Shi,H.,Wang,K.,Bai,M.,2018.A mid-Cretaceousembryonic-to-neonatesnakeinamberfromMyanmar.Sci. Adv.4,eaat5042.