V1-bypassing thalamo-cortical visual circuits in blindsight and developmental dyslexia
Texte intégral
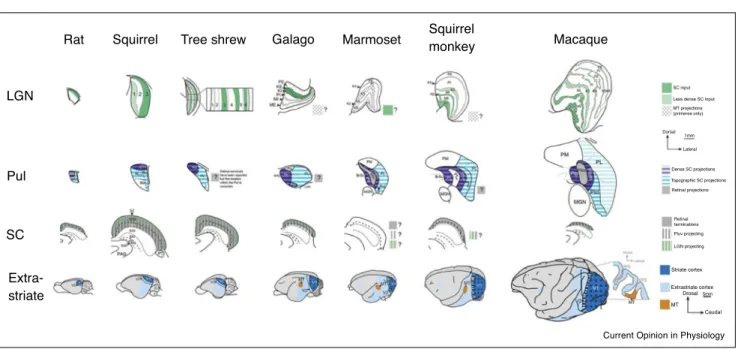
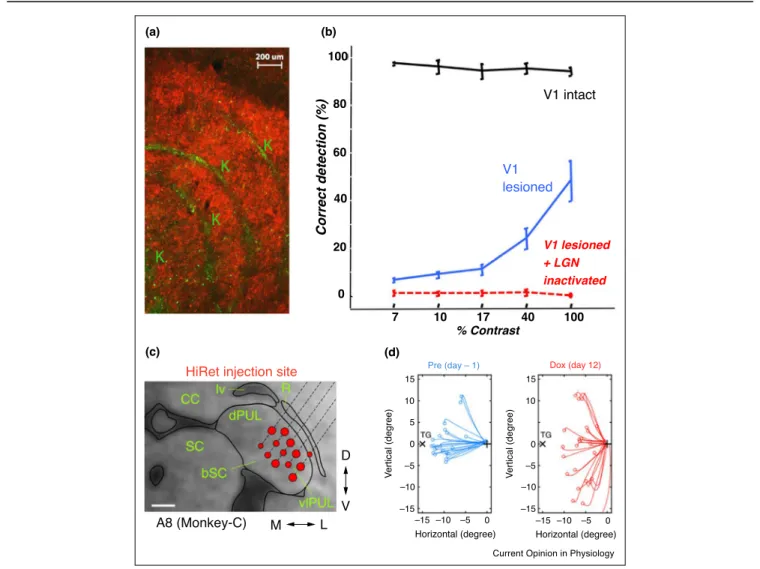
Figure
Documents relatifs
They live in a lovely cottage in Stratford-upon-Avon, England. The house is not very big but my grandmother keeps it clean and tidy. It has only got one floor and the attic. In
Although we have not explored all the cases (the transmission and coupling of a P and general S wave across a lam- ination with non-normal incidence), we are confident that for
In the Southern variety, the absence of low-mid (half-open) front vowels in final open syllables (in words like béquet, Bobet), and the lack of high-mid (half-closed) vowels in
3 A – Today many of the Eskimos live in houses in small towns B because they used to cook inside so sometimes it got too hot.. C their means of transport nowadays are
– We show in Section 2 that accelerated gradient descent, averaged gradient descent and the heavy-ball method for non-strongly-convex problems may be reformulated as constant
If as expected VA span abilities are primarily related to exception word reading performance and reading speed, matching the dyslexic and control groups on their ability to read
We would like to underline the fact that, by explaining this process of an inward vision, physicians (Timothy Bright, Thomas Willis) have discovered that the power of
The absence of evidence for an associated disorder in developmental surface dyslexia and the fact that their reading pattern -- inconsistent words read with less accuracy
