Intense Competition can Drive Selfish Explorers to Optimize Coverage
20
0
0
Texte intégral
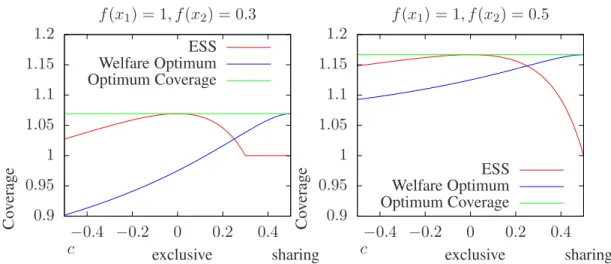
Figure
Documents relatifs