HAL Id: hal-02787042
https://hal.inrae.fr/hal-02787042
Submitted on 5 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Suivi de la croissance et établissement d’allométries
pour 4 génotypes de peupliers en pépinière, en réponse à
la sécheresse
Fabrice Petel
To cite this version:
Fabrice Petel. Suivi de la croissance et établissement d’allométries pour 4 génotypes de peupliers en pépinière, en réponse à la sécheresse. Sciences du Vivant [q-bio]. 2017. �hal-02787042�
Université de Lorraine – Master FAGE –
Fonctionnement et Gestion des Ecosystèmes
Mémoire de stage de Master première année, stage effectué du 24 Avril au 23 Juin 2017, année universitaire 2016 - 2017
Par : Fabrice PETEL
Réalisé sous la direction de : Didier LE THIEC, Directeur de recherches et Maxime DURAND,
doctorant
Centre Inra de Nancy-Lorraine
UMR Ecologie et Ecophysiologie Forestière Equipe PhysioDiv
Rue d'Amance 54280 Champenoux
SUIVI DE LA CROISSANCE ET ÉTABLISSEMENT D’ALLOMÉTRIE DE QUATRE GÉNOTYPES DE PEUPLIERS EN PÉPINIÈRE EN RÉPONSE À LA SÉCHERESSE
Remerciements
Je remercie M. Didier LE THIEC de m’avoir accordé sa confiance pour ce stage, de m’avoir soutenu à chaque étape et de m’avoir fait découvrir le monde de la recherche. Je remercie infiniment Maxime DURAND de m’avoir accompagné tout au long de ce stage et de m’avoir apporté son aide précieuse, notamment lors de mes face à face avec le logiciel R. Je remercie Cyril BURE pour son aide lors de nos sessions de mesures venteuses ou caniculaires. Enfin je remercie Jimmy et son ventilateur, d’avoir partagé mon bureau et mes journées lors de ce stage.
Sommaire
Introduction ... 1
Matériel et méthodes ... 2
1) Matériel végétal et dispositif expérimental... 3
2) Application du déficit hydrique ... 3
3) Suivi de croissance ... 3 4) Analyses statistiques ... 4 5) Expérience en serre ... 4 Résultats ... 5 1) Météorologie ... 5 2) Croissance en pépinière ... 5 3) RGR en pépinière ... 6
3-1) Comparaison des génotypes ... 6
3-2) Effet sécheresse ... 6
4) RGR en serre... 7
4-1) Comparaison entre génotypes ... 7
4-2) Effet sécheresse ... 7
5) Comparaison expérience en serre et pépinière ... 7
Discussion ... 12
Abréviations
Ca : Carpaccio
CO2 : dioxyde de carbone
Dpval : p-value du diamètre Fr : 6J29
GTP : génotype
Hpval : p-value de la hauteur It : N38 I2 : I214 JJ : jour julien NA : données manquantes p : p-value Pep : pépinière
RGR : taux ou vitesse de croissance
RDpval : p-value du taux de croissance en diamètre RHpval : p-value du taux de croissance en hauteur RVpval : p-value du taux de croissance en volume Vpval : p-value du volume
WUE : efficience d’utilisation de l’eau
1
Introduction
La concentration en CO2 dans l’atmosphère a subi une augmentation de 40% depuis 1750 et n’a
pas été aussi élevée depuis 800 000 ans (GIEC 2014). Cette hausse a progressé au rythme le plus rapide jamais observé au cours de la période 2002-2011, essentiellement due à une croissance économique florissante et à l’usage de procédés industriels et de combustibles fossiles (GIEC 2014). L’augmentation prévue de la température de la surface de la Terre, de l’ordre de 0.8°C à 2.5°C d’ici 2050 risque d’entraîner une augmentation de l’évaporation de l’eau des sols et induire des sécheresses plus fréquentes et plus intenses et donc une diminution des rendements de culture (GIEC 2014). Une diminution de la disponibilité en eau induit une réduction de croissance et une diminution de la production de biomasse de l’arbre (Monclus, 2006) ce qui aurait un impact certain sur les espèces grandes consommatrices d’eau comme l’eucalyptus ou le peuplier. L’efficience d’utilisation de l’eau est un indicateur utilisé pour étudier le rendement de l’arbre par unité d’eau consommée et se définit au niveau de la plante entière par le rapport entre le gain de biomasse et la consommation en eau de la plante pendant une période de temps donnée. Comprendre pourquoi certain génotypes arrivent à maintenir leur gain de biomasse tout en limitant leurs pertes en eau en réponse à un déficit hydrique, est un des enjeux essentiels pour pallier aux futures contraintes économiques et environnementales, liées aux changements globaux.
La production de biomasse des végétaux est notamment dépendante de la quantité d’eau présente dans le sol et de la photosynthèse (en lien avec l’interception de la lumière ; Casella et al., 2002). Plusieurs facteurs physiologiques et morphologiques sont responsables d’une différence de croissance entre les différents génotypes. A l’échelle de la plante entière, une forte efficience d’utilisation de l’eau (WUE) est dépendante soit d’une forte accumulation de biomasse soit d’une faible perte en eau. Au niveau foliaire, l’efficience d’utilisation de l’eau intrinsèque (WUEi), est définie comme le rapport entre
le CO2 assimilé par la feuille et la conductance stomatique. Une étude de Monclus et al. (2006) démontre
que la variabilité génotypique de la WUEi est surtout liée à la conductance stomatique plutôt qu’aux
capacités photosynthétiques. De plus, la sécheresse limite grandement la conductance stomatique des feuilles de toutes les espèces d’angiospermes (Bahari et al., 1985). La réponse à un déficit hydrique se caractérise par la fermeture des stomates pour minimiser les pertes en eau (Kozlowski et al., 2002). Nous pouvons définir comme génotype efficients, des arbres qui s’acclimatent à la contrainte de sécheresse en limitant leur perte en eau par contrôle stomatique de la transpiration et en maintenant leur productivité. Néanmoins, la relation entre les paramètres d'échanges gazeux foliaires et les bilans de carbone et d’eau à l’échelle d’une plante entière n’est pas encore bien connue et est difficile d’accès. Néanmoins différentes techniques sont utilisées pour estimer ces paramètres. Des mesures de flux de sève peuvent être réalisées pour estimer la transpiration d’un arbre et des relations allométriques peuvent être utilisées pour estimer la biomasse.
2 La production de biomasse peut être abordée par les traits architecturaux des arbres. Le déploiement en trois dimensions des feuilles dans l'espace a une grande importance dans l'interception de la lumière. La surface foliaire, la position et l’orientation des feuilles peuvent fortement influencer l’efficacité de la capture de l’énergie solaire par le houppier (Barigah et al., 1994). D’autre part, les arbres avec de grandes feuilles ou avec une grande surface foliaire ne sont pas nécessairement les arbres les plus productifs (Niinemets et al., 2003). Les dimensions de la couronne foliaire et le feuillage changent rapidement lorsque les conditions deviennent plus ou moins favorables, c’est pourquoi le diamètre et la hauteur de la couronne sont de bons indicateurs pour prédire la biomasse d’un arbre (Forrester et al., 2017). De plus, la relation entre diamètre et biomasse peut varier notamment selon l’âge, la densité et les conditions du site. La prédiction de la biomasse à l’aide d’un diamètre par exemple diffère souvent de plus de 50% pour une même espèce (Forrester et al., 2017). L’architecture du houppier est donc intimement liée à la production de biomasse (Afas et al., 2007) et peut permettre de quantifier et prédire la biomasse d’un arbre via des relation allométriques (Rae et al., 2004).
Mieux comprendre les mécanismes de réponse à la sécheresse chez le peuplier pourrait élargir les possibilités d’implantation de peupleraies dans des zones où l’eau est présente en plus faible quantité. En effet, le peuplier est la deuxième espèce productrice de bois en France (Rasheed, 2012) et a toujours eu un impact économique majeur dans l’industrie du bois. Une partie importante du bois d’énergie provient du peuplier. Il peut aussi être utilisé en phytoremédiation des sites pollués. Cependant c’est une espèce très sensible aux maladies (e.g. rouille) et à la sécheresse (Monclus et al., 2006). La populiculture est donc essentiellement concentrée dans les sylvoécorégions de vallées alluviales où l’eau n’est pas un facteur limitant (Monclus et al., 2005). Depuis plusieurs dizaines d’années, le peuplier a subi de nombreuses sélections et hybridations afin d’obtenir des clones résistants à la sécheresse, amenant à l’existence de nombreux génotypes et clones différents.
Au sein de la pépinière du site INRA de Champenoux, un suivi de croissance en hauteur et en diamètre de quatre génotypes de peupliers (deux peupliers euramericains : Populus deltoides x nigra et deux peupliers noirs : Populus nigra) a été réalisé depuis 2015. L’objectif de ce stage est de décrire l’évolution de la croissance en hauteur et en diamètre de ces peupliers, d’établir des relations allométriques pour estimer le volume de bois futur de ces quatre génotypes et leur biomasse selon leur régime hydrique. Une comparaison avec les résultats d’un suivi de croissance effectué courant 2016 sur des peupliers cultivés en serre avec les mêmes génotypes sera aussi effectuée afin d’observer si les relations restent les mêmes dans des conditions de cultures différentes.
Matériel et méthodes
L’expérience menée s’inscrit dans un projet de recherche européen, WATBIO, incluant comme modèle biologique le peuplier et qui a pour objectif général d’étudier la réponse à la sécheresse sur plusieurs traits et notamment la production de biomasse. Cette expérience s’inscrit aussi dans le projet
3 UPTRANS (Labex ARBRE) qui évalue l’importance des mécanismes adaptatifs impliqués dans le transport de l’eau en réponse à la sécheresse.
1) Matériel végétal et dispositif expérimental
L’expérience est localisée sur le site de l’INRA-Nancy Grand-Est à Champenoux. Le département de la Meurthe et Moselle subit un climat semi-continental caractérisé par une forte amplitude thermique annuelle avec des hivers froids et des étés assez chauds. Les températures estivales sont en moyenne de 18 à 19,5°C en juillet et les températures hivernales moyennes de 1,5 à 2°C. Sur la station météo de Nancy-Essey, proche du site de l’INRA-Nancy, la température moyenne en janvier est de 1,2°C et de 18,3°C en juillet. En moyenne, il y a 1665 heures de soleil par an à Nancy. Les précipitations sont abondantes toute au long de l’année avec une moyenne de 775,1mm par an en moyenne sur la période 1981 - 2010 (MétéoFrance). On peut noter des précipitations moyennes de 537,3mm pour l’année 2015 et de 809,9mm pour l’année 2016, avec une moyenne de 3.5°C en janvier et de 21,4°C en juillet pour ces deux années combinées (Infoclimat).
144 boutures de peupliers ont été plantées en pépinière au printemps 2014. Les arbres commencent, à l’année 2017, leur 4ème année de croissance. Ces peupliers sont issus de 4 génotypes différents et ont été sélectionnés en fonction de leur tolérance à la sécheresse : 2 génotypes de peuplier noir, Populus nigra (L.) (Français 6J29 et Italien N38) et 2 génotypes (I214 et Carpaccio) d’hybrides euraméricains, Populus canadensis (Moench) (P. nigra (L.) x P. deltoïdes (W. Bartram ex Marshall)). I214 étant classé comme sensible à la sécheresse. La pépinière a une dimension de 53,9 mètres sur 40,22 mètres et se situe en légère pente. Les plants ont été distribués aléatoirement dans cette parcelle mais néanmoins équitablement (Annexe 1). Le dispositif est séparé en 4 blocs (I, II, III et IV) comportant 3 triplets de chaque génotype (soit 9 arbres de chaque génotype) dans chacun des blocs. Le dispositif est entouré d’une rangée de triplets de peupliers, ainsi qu’un triplet de bordure entre le traitement sec et le traitement humide. Dans chaque triplet de génotype, un arbre a été coupé pour estimer la biomasse aérienne initiale. Les mesures s’effectuent donc sur 96 arbres.
2) Application du déficit hydrique
Avant le début de l’expérimentation, les peupliers sont irrigués régulièrement afin de favoriser leur croissance. Le traitement sec se situe en haut de pente et ne concerne que les blocs I et II. La sécheresse progressive est mise en place en juin 2016 avec un arrêt de l’irrigation et à l’aide de bâches imperméables noires, appliquées au sol, qui laissent passer la vapeur d’eau. Cette bâche noire est superposée avec une autre bâche de couleur verte qui réfléchit les rayons lumineux et limite l’impact thermique sous la bâche. Des gouttières sont installées pour évacuer l’eau de pluie.
3) Suivi de croissance
Les mesures de croissance en hauteur et en diamètre sont effectuées toutes les semaines à partir du 27 avril 2017. Une première mesure avant débourrement a été effectuée le 13 mars 2017.
4 La hauteur des arbres est estimée à 1cm du sol jusqu’au bourgeon terminal, avec une perche graduée télescopique de 10 mètres.
Le diamètre est mesuré avec un pied à coulisse au niveau de la bouture. Pour avoir un diamètre moyen et augmenter la précision, deux mesures de diamètre sont effectuées à 90° l’une de l’autre. Les arbres devenant assez haut, à partir du 10 mai 2017, une deuxième mesure de diamètre est effectuée à 1.30 mètre au dessus de la première mesure. Comme lors de la première mesure de diamètre au niveau de la bouture, deux mesures sont effectuées à 1.30 mètre à 90° l’une de l’autre pour ainsi augmenter la fiabilité en effectuant une moyenne de ces deux mesures. La prise de mesure du diamètre à 1.30 mètre a été effectuée en parallèle de la prise de mesure à la base pendant 3 semaines après quoi seule la mesure à 1.30 mètre était effectuée. Une relation allométrique est alors établie permettant d’extraire le coefficient directeur et l’ordonnée à l’origine de chaque arbre et ainsi retrouver le diamètre moyen à la base de la bouture. Le volume a été calculé de la manière suivante :
Le taux de croissance (RGR) est calculé pour la hauteur, le diamètre et le volume entre chaque semaine avec la formule suivante :
4) Analyses statistiques
Après avoir effectué les mesures, les données sont encodées sous Excel puis analysées sous le logiciel R. La normalité des résidus est testée, si elle est vérifiée une ANOVA est appliquée. Si l’ANOVA est significative au seuil p-value = 0,05 soit 5%, on peut effectuer une comparaison en faisant un test de Tukey. Des tests de Student ont été réalisés pour tester jour par jour si les différences entre traitements étaient significatives. Une ANCOVA est appliquée pour la comparaison entre les expériences en serre et en pépinière (script développé par Pierre Montpied).
5) Expérience en serre
48 boutures de peupliers ont été mises en pots dans un mélange tourbe-sable (50/50 v/v) avec du nutricot NPK (13-13-13 + 2 oligoéléments) en serre climatisée. Du gravier a été disposé pour limiter les pertes en eau. Les 4 génotypes sélectionnés pour cette expérience sont les mêmes que ceux utilisés pour l’expérience en pépinière (soit 12 plants de chaque génotype). La sécheresse est mise en place le 31 mai 2016 par arrêt de l’arrosage pour 6 boutures de chaque génotype, soit 24 plants. L’humidité volumique du sol en condition de sécheresse est de 18%. 24 autres boutures font par ailleurs office de témoin avec un régime hydrique normal (arrosage pour maintenir 80% de la capacité au champ, équivalent à 24% d’humidité volumique du sol dans notre cas).
5 Les mesures des paramètres de croissance ont été effectuées du 26 avril au 14 juin 2016, toutes les semaines. La hauteur du plant a été mesurée avec une règle graduée de la tige principale au bourgeon terminal. Les diamètres ont, quant à eux, été mesurés avec un pied à coulisse à 1 cm de la base de la tige principale du jeune plant. Les arbres ont été coupés le 22 juin 2016.
Résultats
1) Météorologie
Les données utilisées sont celles de la station météo de Tomblaine (54526), située à 14km de la pépinière, en Meurthe-et-Moselle, France. L’année 2015 a été marquée par une sécheresse bien établie (537,3 mm de cumul de pluie) contrairement à l’année 2016 qui a été assez pluvieuse (755,1 mm de cumul de pluie). L’année 2017 semble suivre la tendance de l’année 2015 avec peu de pluie, un rayonnement global important et des températures assez élevées. La température moyenne sur l’année 2015 était de 13,4°C et de 10,85 °C pour 2016.
2) Croissance en pépinière
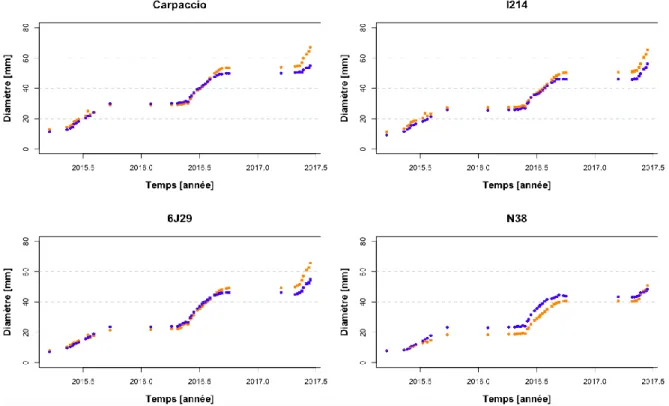
Les tests montrent une différence significative entre tous les génotypes pour la croissance en hauteur et en volume (p < 0,0001) (Figures 1 et 3). Seule la croissance en diamètre entre le génotype I214 et le génotype 6J29 n’est pas significative (p=0,07) (Figure 2). La hauteur moyenne avec les différents traitements et sur les 3 années de croissance des peupliers euraméricains est plus élevée que celle des peupliers noirs (Carpaccio : 304,2 cm ; I214 : 275,5 cm ; 6J29 : 260,7 cm; N38 : 250,7 cm). De plus, la tendance est la même pour le diamètre moyen et le volume moyen (diamètre : Carpaccio : 37,1 mm ; I214 : 34,4 mm ; 6J29 : 32,3 mm ; N38 : 28,7 mm; volume : Carpaccio : 1530,0 cm3 ; I214 : 1233,7 cm3 ; 6J29 : 1158,1 cm3 ; N38 : 861,0 cm3).
La hauteur, le diamètre et le volume sont significativement différents entre les traitements (respectivement : p < 0,0001 ; p=0,01 ; p=0,0004). Pour l’ensemble des années, la hauteur est significativement plus élevée dans le traitement contrôle pour les peupliers noirs (6J29 : du jour 120 de l’année 2016 et du jour 180 au jour 238 de la même année ; N38 : du jour 217 de l’année 2015 au jour 166 de l’année 2017), alors qu’il n’y a pas de différence significative entre les traitements pour les peupliers euraméricains. De plus, la croissance en diamètre est significativement plus élevée en traitement sec qu’en traitement contrôle pour tous les génotypes (Carpaccio : du jour 251 de 2016 au jour 166 de 2017 ; I214 : le 78e jour de l’année 2015, le 198e jour de l’année 2015 et du 137 au 166e jour de l’année 2017 ; 6J29 : du jour 117 au jour 166 de l’année 2017) sauf pour le génotype N38 où le diamètre est significativement plus élevé en traitement humide (du jour 203 de l’année 2015 au jour 230 de l’année 2016).
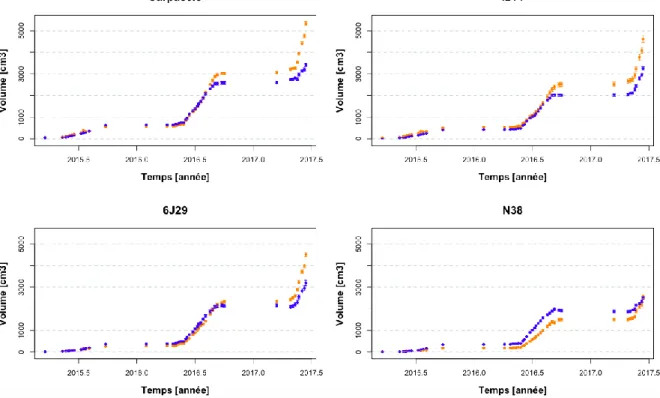
La croissance en volume est significativement plus forte en traitement contrôle pour le génotype N38 du jour 217 de l’année 2015 au jour 251 de l’année 2016. Pour les autres génotypes, le contraire est
6 mis en évidence par une croissance en volume significativement plus forte en traitement sous sécheresse (Carpaccio : jour 125 de 2017 et du 152 au 166 de 2017 ; I214 et 6J29 : du 152 au 166e jour de 2017). En effet, les arbres ont un volume moyen qui augmente pour tous les génotypes et traitements pour chaque année successive (2015 : 600,2 cm3 ; 2016 : 2486,5 cm3 ; 2017 : 3840,7 cm3).
3) RGR en pépinière
3-1) Comparaison des génotypes
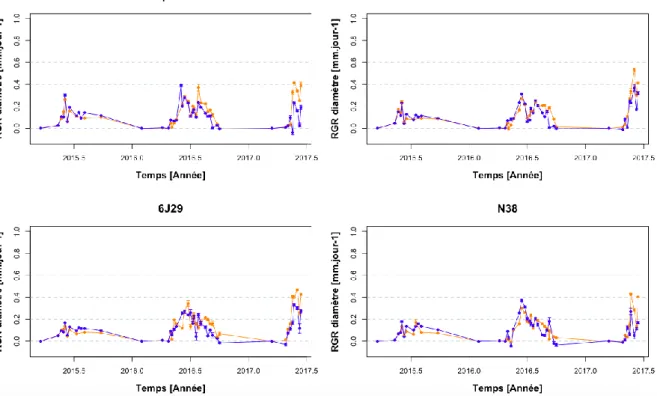
Les 4 génotypes semblent avoir des RGR en hauteur et en diamètre similaires (Figures 4 et 5). Les tests statistiques n’ont pas montré de différence significative entre les différents génotypes pour le RGR en hauteur pour les 3 années (p=0,88). Par contre une différence significative entre les génotypes pour le RGR en diamètre a pu être mise en évidence entre N38 et 6J29 (p=0,01 ; Carpaccio : 0,12 mm.jour-1 ; I214 : 0,12 mm.jour-1 ; 6J29 : 0,13 mm.jour-1 ; N38 : 0,11 mm.jour-1). De plus, les tests ont montré une différence significative entre N38 et les trois autres génotypes sur le RGR en volume (dans tous les cas p < 0,0001; Carpaccio : 11,4 cm3.jour-1 ; I214 : 10,6 cm3.jour-1 ; 6J29 : 10,9 cm3.jour-1 ; N38 : 7,11 cm3.jour-1) (Figure 6).
3-2) Effet sécheresse
Les tests statistiques n’ont pas montré de différence significative entre les 2 traitements pour le RGR en hauteur pour les 3 années (p=0,80) (Figure 4). Par contre une différence significative entre les 2 traitements pour le RGR en diamètre et en volume a pu être mise en évidence pour tous les génotypes hormis le RGR en volume de N38 (diamètre : p=0,0004 pour tous les génotypes car l’interaction génotype-traitement n’est pas significative; volume : respectivement pour Carpaccio, I214, 6J29 et N38 : p < 0,0001 ; p=0,003 ; p=0,005 ; p=0,99) (Figures 5 et 6). En 2015, le RGR en hauteur est significativement plus élevé dans le traitement contrôle que dans le traitement sous sécheresse pour les jours 203 à 267 pour Carpaccio (dans tous les cas p < 0,04), des jours 198 à 267 pour I214 (dans tous les cas p < 0,05), des jours 198 à 267 pour 6J29 (dans tous les cas p < 0,02), des jours 169 à 267 pour N38 (dans tous les cas p < 0,02). Il en va de même pour le RGR en diamètre pour les jours 198 à 217 pour Carpaccio (dans tous les cas p < 0,02), des jours 198 à 217 pour I214 (dans tous les cas p < 0,04), des jours 190 et 203 pour 6J29 (dans tous les cas p < 0,01) et des jours 203 à 267 pour N38 (dans tous les cas p < 0,05). Enfin, le RGR en volume est significativement plus élevé dans le traitement contrôle que dans le traitement sous sécheresse des jours 198 à 217 pour Carpaccio (dans tous les cas p < 0,03), des jours 198 à 203 pour I214 (dans tous les cas p < 0,03), des jours 203 à 217 pour 6J29 (dans tous les cas p < 0,04) et des jours 203 à 267 pour N38 (dans tous les cas p < 0,04).
En 2016, le RGR en hauteur, diamètre et volume est plus élevé sur le traitement sous sécheresse que sur le traitement contrôle pour quelques dates en fin de saison de croissance (Annexe 2). Cette tendance se poursuit sur l’année 2017 du jour 137 à 166 pour les 3 taux de croissance du génotype Carpaccio (dans tous les cas p=0,002). Le RGR en hauteur est significativement plus élevé en 2017 qu’en 2015 ou 2016 (pour
7 2015-2016 : p=0,48; pour 2016-2017: p < 0,0001). En effet, le RGR en hauteur moyen en 2015 pour tous les génotypes et traitements est de 0,81 cm.jour-1 alors qu’il est de 0,76 cm.jour-1 en 2016 et 0,40 cm.jour-1 en 2017. Le RGR en diamètre et volume augmente de manière significative chaque année (dans tous les cas p < 0,0001). Effectivement, le RGR en diamètre moyen pour tous les génotypes et traitements est de 0,08 ; 0,12 ; 0,16 mm.jour-1 et le RGR en volume de 1,84 ; 9,88 ; 21,24 cm3.jour-1 respectivement pour 2015, 2016 et 2017.
4) RGR en serre
4-1) Comparaison entre génotypes
Les tests statistiques ont montré une différence entre les génotypes pour la vitesse de croissance en hauteur (p=0,03) et en volume (p < 0,0001) (Figures 7 et 9). En effet, le génotype Carpaccio a un RGR en hauteur significativement plus fort que les 2 génotypes de peupliers noirs (6J29 : p=0,03 ; N38 : p=0,009). De plus, les tests montrent une différence significative entre les peupliers euraméricains et les peupliers noirs sur le RGR en volume (Carpaccio : 1,66 cm3.jour-1 ; I214 : 1,56 cm3.jour-1 ; 6J29 : 1,03 cm3.jour-1 ; N38 : 0,95 cm3.jour-1). En revanche, le RGR en diamètre n’est pas significativement différent entre les génotypes (p=0,12) (Figure 8).
4-2) Effet sécheresse
En faisant un test de Student pour chaque jour, on s’aperçoit qu’après mise en condition de sécheresse au jour julien 153, une différence significative entre les traitements pour la vitesse de croissance en hauteur est mise en évidence pour tous les génotypes au jour 159 (p < 0,0002 pour tous les cas) et au jour 166 pour les génotypes 6J29 et N38 (p < 0,002 dans les deux cas) avec un RGR en hauteur plus fort en traitement contrôle (Figure 7). La vitesse de croissance en volume est significativement différente en fonction du traitement au jour 159 pour les génotypes Carpaccio et 6J29 (p < 0,007 pour les deux cas) et au jour 166 pour les génotypes I214 et 6J29 (p < 0,02 pour les deux cas) avec un RGR en volume plus important en traitement contrôle (Figure 9). En ce qui concerne la vitesse de croissance en diamètre après mise en sécheresse des plants, seul le génotype Carpaccio au jour 159 a une différence significative entre les deux traitements (p=0,006) avec un RGR en volume plus fort pour le contrôle (Figure 8).
5) Comparaison expérience en serre et pépinière
L ‘ANCOVA a montré une différence très significative entre les deux types de culture, tous génotypes, tous traitements et toutes années confondus (p < 0,0001 pour l’ordonnée à l’origine et pour la pente). Les arbres du type de culture en serre ont tendance à gagner beaucoup plus en hauteur qu’en diamètre que les peupliers du traitement contrôle (Figure 10). Les tests statistiques ont aussi montré un effet génotype significatif tous traitements, toutes les années et tous types de culture confondus (dans tous les cas p < 0,002 pour l’ordonnée à l’origine et p < 0,03 pour la pente), sauf pour les deux génotypes de
8 peupliers euraméricains qui ont une ordonnée à l’origine presque équivalent (p=0,99) et pour les génotypes I214 et 6J29 qui ont des pentes très semblables (p=0,27).
Dans cette comparaison des expériences en serre et en pépinière, il y a aussi un effet traitement significatif (p < 0,0001 pour l’ordonnée à l’origine et pour la pente). Les peupliers en traitement sec ont une tendance à pousser plus en hauteur qu’en diamètre que les peupliers en traitement contrôle.
Figure 1 : Evolution de la hauteur (en cm) des 4 génotypes en pépinière entre 2015, 2016 et 2017. Le traitement contrôle est
représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
Figure 2 : Evolution du diamètre (en mm) des 4 génotypes en pépinière entre 2015, 2016 et 2017. Le traitement contrôle est
9
Figure 3 : Evolution du volume (en cm3) des 4 génotypes en pépinière entre 2015, 2016 et 2017. Le traitement contrôle est représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
Figure 4 : Evolution du taux de croissance en hauteur (RGR en cm.jour-1) des 4 génotypes en pépinière entre 2015, 2016 et 2017. Le traitement contrôle est représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
10
Figure 5 : Evolution du taux de croissance en diamètre (RGR en mm.jour-1) des 4 génotypes en pépinière entre 2015, 2016 et 2017. Le traitement contrôle est représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
Figure 6 : Evolution du taux de croissance en volume (RGR en cm3.jour-1) des 4 génotypes en pépinière entre 2015, 2016 et 2017. Le traitement contrôle est représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
11
Figure 7 : Evolution du taux de croissance en hauteur (RGR en cm.jour-1) des 4 génotypes en serre en 2016. Le traitement contrôle est représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
Figure 8 : Evolution du taux de croissance en diamètre (RGR en mm.jour-1) des 4 génotypes en serre en 2016. Le traitement contrôle est représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
12
Figure 9 : Evolution du taux de croissance en volume (RGR en cm3.jour-1) des 4 génotypes en serre en 2016. Le traitement contrôle est représenté en bleu ( ) et le traitement sous sécheresse est représenté en jaune ( ).
Figure 10 : Comparaison des différents génotypes en fonction du type de culture et selon le traitement (sécheresse ou humide).
La culture en serre avec le traitement contrôle est représentée en bleu clair ( ) et la culture en serre avec le traitement sous sécheresse est représentée en bleu foncé ( ). La culture en pépinière avec le traitement contrôle est représentée en rouge clair ( ) et le traitement sous sécheresse est représentée en rouge foncé ( ).
Discussion
L’étude de la croissance a permis de mettre en évidence une différence interspécifique entre les peupliers noirs et euraméricains. La hauteur, le diamètre et le volume des peupliers euraméricains sont
13 plus importants (en particulier chez le génotype Carpaccio) que chez les peupliers noirs et ce dans n’importe quel traitement. Le génotype N38 quant à lui se démarque avec la croissance la plus faible pour tous les paramètres de croissance. Ce génotype a tendance à faire des sylleptiques ce qui pourrait expliquer en partie son retard dans la croissance en hauteur, du fait qu’il alloue plus de carbone dans les branches latérales. Les peupliers noirs possèdent une forme de croissance plus arbustive que les peupliers euraméricains avec bien plus de sylleptiques. Toutefois, mon stage s’est concentré sur la croissance de la tige principale. Le volume du houppier des peupliers noirs serait donc plus imposant alors que les peupliers euraméricains seraient plus axés sur une croissance en hauteur de la tige principale. L’analyse de la hauteur, du diamètre et du volume par rapport aux différents traitements montre que les peupliers euraméricains seraient moins sensibles à la sécheresse et que le génotype N38 le serait le plus. Effectivement, la sécheresse entraîne une baisse de la conductivité hydraulique chez les peupliers noirs qui peut aller jusqu’à la cavitation des vaisseaux du xylème les rendant non fonctionnels (Fichot et al., 2015). Cette baisse est d’autant plus importante chez les peupliers noirs que chez les peupliers euraméricains. Néanmoins pour le diamètre et le volume, le traitement sous sécheresse est marqué par des diamètres et des volumes plus élevés à partir de la fin de l’année 2016 sur les arbres à la pépinière.
Il faut signifier que la hauteur et le diamètre moyen augmentent d’année en année. Or lors des résultats ce n’est pas le cas pour l’année 2017. Dans ce jeu de données, toutes les mesures pour l’année 2017 n’ont pas encore été effectuées, les hauteurs et les diamètres moyens sont donc logiquement significativement plus faibles que l’année 2016 car les moyennes sont biaisées.
L’étude de la vitesse de croissance en hauteur, en diamètre et en volume des peupliers en pépinière a montré que la vitesse de croissance en volume pour le génotype N38 est plus faible que chez les 3 autres génotypes. L’effet du traitement n’est pas mis en évidence dans l’expérience en pépinière, notamment depuis mi/fin 2016. D’après les données météorologiques récoltées, une sécheresse était bien établie à l’année 2015, et l’année 2016 était une année pluvieuse. Jusqu’au mois de juin de cette année, l’année 2017 a une tendance à rejoindre l’année 2015 et si cette tendance perdure on devrait donc observer une baisse de la croissance sur le traitement sec. Néanmoins, les résultats montrent une croissance et un taux de croissance plus importants pour ce traitement. Ce phénomène peut être expliqué par un éventuel effet de la bâche qui pourrait augmenter la température du sol et entraîner une activité des microorganismes plus importante et favoriser la prise d’éléments minéraux. Des analyses du sol ont montré une concentration en azote plus importante dans le traitement sec. Malgré ces contraintes indépendantes de l’expérience, le génotype N38 semble moins bien supporter la sécheresse que les autres génotypes, la différence de croissance entre traitement contrôle et sous sécheresse étant plus importante. L’analyse de la croissance en fonction de l’année montre que la vitesse de croissance est de plus en plus forte au fil des années. En effet, au fur et à mesure des années, les arbres augmentent leur surface foliaire totale et donc leur capacité d’assimilation photosynthétique, la mise en réserve notamment des composés
14 carbonés serait donc plus importante pour l’année suivante. Ce carbone stocké sera remobilisé au printemps pour la croissance (Gilson, 2015). Néanmoins, le traitement des résultats pour la vitesse de croissance de ces 3 paramètres en fonction de l’année est aussi biaisé par le fait que toutes les mesures de l’année 2017 n’ont pas encore été effectuées. La vitesse de croissance en hauteur pour 2017 est donc significativement plus faible que les deux années précédentes mais pour la vitesse de croissance en diamètre et en volume, l’augmentation est telle que malgré le biais on observe quand même une augmentation de la vitesse de croissance de ces deux paramètres.
Les tests statistiques sur l’expérience en serre mettent en évidence une vitesse de croissance en volume plus élevée pour les peupliers euraméricains. Le génotype Carpaccio possède aussi une vitesse de croissance en hauteur supérieure aux deux génotypes de peupliers noirs. L’analyse vis à vis du traitement sous sécheresse a montré une baisse significative de la vitesse de croissance en hauteur une semaine après la mise en sécheresse, pour tous les génotypes. En revanche, le taux de croissance en diamètre et en volume semble très peu impacté par ce traitement (il est à noter que la sécheresse appliquée est modérée).
Enfin en comparaison des deux expériences en serre et en pépinière, la croissance des peupliers cultivés en serre se fait plus en hauteur qu’en diamètre comparé aux peupliers en pépinière. De plus, du fait des structures de la serre et de la densité de plantation dans l’expérience avec les plants en serre, la lumière est plus difficilement accessible, ce qui pousserait les arbres à réduire l’investissement dans une croissance en diamètre pour privilégier une croissance en hauteur pour ainsi aller chercher la lumière. Par ailleurs, le vent est un facteur majeur qui influence la croissance, qui s’ajuste en fonction de cette contrainte. Les arbres exposés au vent ont tendance à avoir une réduction de croissance de la tige longitudinale et à avoir une plus grande croissance radiale que les plantes abritées (Gardiner et al., 2016). Bonnesoeur et al. (2016) ont aussi montré une augmentation de la croissance radiale chez le hêtre en imposant un stress mécanique (simulant un stress d’épisode très venteux). On peut donc supposer que le vent joue en rôle important dans la prise en diamètre des peupliers en pépinière.
Nous pouvons conclure que pour un rendement important dans les cultures à cycle court de peupliers, le génotype Carpaccio de l’espèce P. Canadensis paraît être le meilleur. Les peupliers euraméricains privilégient une croissance en hauteur par rapport à une croissance des sylleptiques, ce qui est plus avantageux du point de vue commercial pour faire des grumes par exemple. Le génotype Carpaccio est plus apte à être utilisé que le génotype I214 car ce dernier à une moins bonne réponse aux contraintes de sécheresse et aux maladies du fait d’une conductance stomatique forte même lorsque l’eau devient un élément limitant (MD et DLT, communication personnelle). Le génotype N38 ne serait pas un bon choix car il résiste moins bien à la sécheresse comme l’a démontré l’année 2015 sur RGR. Les travaux que j’ai menés ici font partie intégrante d’une thèse qui s’intéresse à l’efficience d’utilisation de l’eau et ils seront bien sûr à mettre en relation avec tous les autres paramètres physiologiques enregistrés comme la transpiration et la photosynthèse.
15
Références
GIEC, 2014. Climate change 2014, synthesis report.
AFAS NAL., MARRON N., CEULEMANS R. 2007. Variability in Populus leaf anatomy and
morphology in relation to canopy position, biomass production, and varietal taxon. Annals of Forest
Science. Vol. 64, n°5, p. 521-532.
BAHARI ZA., PALLARDY SG., PARKER WC. 1985. Photosynthesis, water relations, and drought
adaptation in six woody species of oak-hickory forests in Central Missouri. Forest Science. Vol. 31, n°3, p. 557-569.
BARIGAH TS., SAUGIER B., MOUSSEAU M., GUITTET J., CEULEMANS R. 1994.
Photosynthesis, leaf area and productivity of 5 poplar clones during their establishment year. Annals
of Forest Science. Vol. 51, n°6, p. 613-625.
BONNESOEUR V., CONSTANT T., MOULIA B., FOURNIER M., 2016. Forest trees filter chronic
wind-signals to acclimate to high winds. New Phytologist. Vol. 210, p. 850-860.
CASELLA E., CEULEMANS R. 2002. Spatial distribution of leaf morphological and physiological
characteristics in relation to local radiation regime within the canopies of 3-year-old Populus clones in coppice culture. Tree Physiology. Vol. 22, p. 1277-1288.
FICHOT R., BRIGNOLAS F., COCHARD H., CEULEMANS R. 2015. Vulnerability to
drought-induced cavitation in poplars: synthesis and future opportunities. Plant, Cell & Environment. Vol. 38, p. 1233-1251.
FORRESTER DI., TACHAUER IHH., ANNIGHOEFER P., BARBEITO I., PRETZSCH H., RUIZ-PEINADO R., STARK H., VACCHIANO G., ZLATANOV T., CHAKRABORTY T., SAHA S., SILESHI GW. 2017. Generalized biomass and leaf area allometric equations for
European tree species incorporating stand structure, tree age and climate. Forest Ecology and
Management. Vol. 396, p. 160-175.
GARDINER B., BERRY P., MOULIA B. 2016. Review: Wind impacts on plant growth, mechanics
and damage. Plant Science. Vol. 245, p. 94-118.
GILSON Angelique, 2015. Dynamique saisonnière et répartition du Carbone et de l’Azote : de l’organe
au peuplement. Rôle des réserves et effets de l’âge chez le chêne sessile (Quercus petraea). Ecologie, Environnement. Université Paris Sud - Paris XI.
KOZLOWSKI TT., PALLARDY SG. 2002. Acclimatation and adaptative responses of woody plants to
environmental stresses. The Botanical Review. Vol. 68, p. 270-334.
MONTCLUS R., DREYER E., DELMOTTE FM., VILLAR M., DELAY D., BOUDOURESQUE E., PETIT J-M., MARRON N., BRECHET C., BRIGNOLAS F. 2005. Productivity, leaf traits
and carbon isotope discrimination in 29 Populus deltoides x P. nigra clones. New Phytologist. Vol. 167, p. 53-62.
MONTCLUS R., DREYER E., VILLAR M., DELMOTTE FM., DELAY D., PETIT J-M., BARBAROUX C., LE THIEC D., BRECHET C., BRIGNOLAS F. 2006. Impact of drought on
productivity and water efficiency in 29 genotypes of Populus deltoides x Populus nigra. New
Phytologist. Vol. 169, p. 765-777.
NIINEMETS U., AFAS NAL., CESCATTI A., PELLIS A., CEULEMANS R. 2004. Petiole length
and biomass investment in support modify light- interception efficiency in dense poplar plantations.
Tree Physiology. Vol. 24, p. 141-154.
RAE AM., ROBINSON KM., STREET NR., TAYLOR G. 2004. Morphological and physiological
traits influencing biomass productivity in short-rotation coppice poplar. Revue canadienne de
recherche forestière. Vol. 34, p. 1488-1498.
RASHEED Fahad, 2012. Components of transpiration efficiency in poplars : genetic diversity, stability
with age and scaling from leaf to whole plant level. PhD thesis. Silviculture, forestry. AgroParisTech.
Annexes
Annexe 1 : Plan de l’expérience en pépinière. Le traitement sec est représenté en orange, le traitement humide en bleu.
Une succession de trois points verts représente les trois arbres d’un plot tandis qu’une succession de trois points orange représente les arbres de bordure. Ca = Carpaccio (P.nigra X P.deltoïdes). I2 = I214 (P.nigra X P.deltoïdes). Fr = 6J29 (P.nigra). It = N38 (P.nigra).
Tableau 1
Culture GTP Année JJ Hpval Dpval Vpval RHpval RDpval RVpval
Pep Carpaccio 2015 78 0.669 0.054 0.099 NA NA NA Pep Carpaccio 2015 131 0.776 0.065 0.132 0.586 0.939 0.385 Pep Carpaccio 2015 141 0.761 0.096 0.160 0.848 0.299 0.646 Pep Carpaccio 2015 148 0.761 0.071 0.131 0.933 0.055 0.081 Pep Carpaccio 2015 155 0.796 0.186 0.253 0.638 0.271 0.952 Pep Carpaccio 2015 162 0.912 0.200 0.284 0.153 0.872 0.840 Pep Carpaccio 2015 169 0.972 0.307 0.386 0.138 0.181 0.777 Pep Carpaccio 2015 190 0.949 0.386 0.423 0.827 0.871 0.627 Pep Carpaccio 2015 198 0.939 0.103 0.121 0.257 0.017 0.023 Pep Carpaccio 2015 203 0.752 0.780 0.757 0.028 0.002 0.002 Pep Carpaccio 2015 217 0.609 0.914 0.954 0.031 0.000 0.017 Pep Carpaccio 2015 267 0.214 0.629 0.542 0.019 0.078 0.142 Pep Carpaccio 2016 29 0.257 0.617 0.558 0.274 1.000 0.871 Pep Carpaccio 2016 96 0.264 0.593 0.537 0.941 0.614 0.496 Pep Carpaccio 2016 112 0.264 0.593 0.537 NA NA NA Pep Carpaccio 2016 120 0.259 0.500 0.457 0.844 0.184 0.153 Pep Carpaccio 2016 124 0.246 0.606 0.546 0.499 0.066 0.107 Pep Carpaccio 2016 133 0.325 0.552 0.536 0.002 0.478 0.696 Pep Carpaccio 2016 140 0.289 0.571 0.545 0.182 0.763 0.875 Pep Carpaccio 2016 146 0.337 0.561 0.551 0.126 0.776 0.657 Pep Carpaccio 2016 153 0.402 0.623 0.626 0.160 0.412 0.751 Pep Carpaccio 2016 158 0.498 0.649 0.667 0.104 0.634 0.712 Pep Carpaccio 2016 166 0.571 0.675 0.673 0.373 0.731 0.787 Pep Carpaccio 2016 175 0.534 0.717 0.702 0.768 0.630 0.925 Pep Carpaccio 2016 182 0.581 0.716 0.723 0.530 0.941 0.914 Pep Carpaccio 2016 189 0.556 0.892 0.833 0.787 0.003 0.230 Pep Carpaccio 2016 194 0.577 0.916 0.845 1.000 0.629 0.951 Pep Carpaccio 2016 202 0.551 0.985 0.892 0.637 0.783 0.844 Pep Carpaccio 2016 208 0.548 0.679 0.850 0.820 0.178 0.146 Pep Carpaccio 2016 216 0.726 0.559 0.725 0.161 0.148 0.152 Pep Carpaccio 2016 230 0.795 0.201 0.353 0.411 0.000 0.000 Pep Carpaccio 2016 238 0.840 0.149 0.285 0.652 0.085 0.095 Pep Carpaccio 2016 246 0.856 0.099 0.185 0.010 0.126 0.031 Pep Carpaccio 2016 251 0.931 0.048 0.103 0.974 0.033 0.396 Pep Carpaccio 2016 266 0.566 0.043 0.076 0.734 0.394 0.705 Pep Carpaccio 2016 274 0.565 0.044 0.078 0.947 0.888 0.940 Pep Carpaccio 2017 72 0.631 0.034 0.069 0.075 0.223 0.314 Pep Carpaccio 2017 117 0.736 0.021 0.055 0.301 0.882 0.734 Pep Carpaccio 2017 125 0.528 0.018 0.040 0.010 0.959 0.255 Pep Carpaccio 2017 130 0.540 0.039 0.074 0.457 0.298 0.243 Pep Carpaccio 2017 137 0.424 0.001 0.005 0.167 0.000 0.000 Pep Carpaccio 2017 144 0.500 0.000 0.002 0.359 0.000 0.000 Pep Carpaccio 2017 152 0.223 0.000 0.000 0.000 0.000 0.000 Pep Carpaccio 2017 159 0.100 0.000 0.000 0.000 0.001 0.000 Pep Carpaccio 2017 166 0.024 0.000 0.000 0.000 0.016 0.000
Pep I214 2015 78 0.126 0.044 0.047 NA NA NA Pep I214 2015 131 0.082 0.073 0.056 0.073 0.754 0.083 Pep I214 2015 141 0.065 0.074 0.052 0.042 0.162 0.045 Pep I214 2015 148 0.058 0.078 0.050 0.068 0.222 0.048 Pep I214 2015 155 0.062 0.118 0.061 0.460 0.759 0.108 Pep I214 2015 162 0.072 0.110 0.061 1.000 0.253 0.087 Pep I214 2015 169 0.090 0.173 0.081 0.707 0.027 0.482 Pep I214 2015 190 0.298 0.215 0.122 0.096 0.806 0.398 Pep I214 2015 198 0.410 0.038 0.031 0.002 0.021 0.025 Pep I214 2015 203 0.517 0.287 0.167 0.014 0.009 0.025 Pep I214 2015 217 0.628 0.378 0.216 0.042 0.032 0.894 Pep I214 2015 267 0.970 0.516 0.343 0.049 0.481 0.984 Pep I214 2016 29 0.962 0.438 0.310 0.120 0.137 0.205 Pep I214 2016 96 0.931 0.483 0.325 0.078 0.399 0.991 Pep I214 2016 112 0.931 0.483 0.325 NA NA NA Pep I214 2016 120 0.992 0.544 0.364 0.139 0.214 0.378 Pep I214 2016 124 0.972 0.405 0.283 0.657 0.130 0.344 Pep I214 2016 133 0.961 0.539 0.369 0.853 0.181 0.270 Pep I214 2016 140 0.989 0.549 0.373 0.212 0.762 0.766 Pep I214 2016 146 0.980 0.544 0.373 0.471 0.750 0.618 Pep I214 2016 153 0.915 0.711 0.498 0.301 0.162 0.612 Pep I214 2016 158 0.970 0.826 0.565 0.329 0.014 0.311 Pep I214 2016 166 0.796 0.929 0.697 0.039 0.088 0.277 Pep I214 2016 175 0.808 0.945 0.706 0.999 0.842 0.811 Pep I214 2016 182 0.586 0.877 0.723 0.199 0.213 0.966 Pep I214 2016 189 0.531 0.792 0.675 0.446 0.060 0.214 Pep I214 2016 194 0.492 0.832 0.737 0.476 0.445 0.430 Pep I214 2016 202 0.357 0.802 0.759 0.024 0.504 0.877 Pep I214 2016 208 0.444 0.820 0.747 0.102 0.758 0.651 Pep I214 2016 216 0.556 0.820 0.754 0.220 0.996 0.915 Pep I214 2016 230 0.631 0.568 0.543 0.493 0.001 0.010 Pep I214 2016 238 0.652 0.395 0.406 0.871 0.003 0.027 Pep I214 2016 246 0.943 0.414 0.390 0.005 0.877 0.407 Pep I214 2016 251 0.935 0.257 0.261 0.078 0.000 0.000 Pep I214 2016 266 0.701 0.142 0.152 0.014 0.001 0.000 Pep I214 2016 274 0.688 0.139 0.143 0.763 0.576 0.337 Pep I214 2017 72 0.779 0.121 0.140 0.067 0.754 0.848 Pep I214 2017 117 0.727 0.065 0.082 0.330 0.003 0.002 Pep I214 2017 125 0.721 0.082 0.100 0.840 0.095 0.118 Pep I214 2017 130 0.742 0.058 0.080 0.541 0.011 0.012 Pep I214 2017 137 0.814 0.048 0.072 0.350 0.675 0.755 Pep I214 2017 144 0.812 0.027 0.052 0.905 0.329 0.208 Pep I214 2017 152 0.765 0.010 0.029 0.345 0.094 0.033 Pep I214 2017 159 0.719 0.004 0.016 0.351 0.011 0.001 Pep I214 2017 166 0.418 0.003 0.010 0.000 0.051 0.001 Pep 6J29 2015 78 0.372 0.279 0.540 NA NA NA Pep 6J29 2015 131 0.429 0.366 0.680 0.879 0.799 0.842 Pep 6J29 2015 141 0.499 0.355 0.669 0.279 0.633 0.646
Pep 6J29 2015 148 0.513 0.306 0.639 1.000 0.079 0.492 Pep 6J29 2015 155 0.556 0.434 0.752 0.556 0.066 0.773 Pep 6J29 2015 162 0.657 0.463 0.813 0.232 0.667 0.779 Pep 6J29 2015 169 0.743 0.563 0.935 0.292 0.053 0.447 Pep 6J29 2015 190 1.000 0.902 0.785 0.391 0.002 0.290 Pep 6J29 2015 198 0.655 0.291 0.687 0.000 0.091 0.165 Pep 6J29 2015 203 0.502 0.657 0.395 0.015 0.015 0.009 Pep 6J29 2015 217 0.222 0.477 0.241 0.000 0.062 0.037 Pep 6J29 2015 267 0.051 0.193 0.093 0.002 0.275 0.067 Pep 6J29 2016 29 0.054 0.283 0.132 0.908 0.044 0.090 Pep 6J29 2016 96 0.067 0.309 0.149 0.295 0.975 0.645 Pep 6J29 2016 112 0.067 0.309 0.149 NA NA NA Pep 6J29 2016 120 0.050 0.300 0.135 0.284 0.604 0.223 Pep 6J29 2016 124 0.054 0.253 0.120 0.554 0.209 0.258 Pep 6J29 2016 133 0.059 0.487 0.229 0.088 0.011 0.121 Pep 6J29 2016 140 0.052 0.563 0.245 0.210 0.279 0.729 Pep 6J29 2016 146 0.052 0.524 0.238 0.531 0.276 0.803 Pep 6J29 2016 153 0.054 0.545 0.245 0.394 0.930 0.302 Pep 6J29 2016 158 0.085 0.358 0.179 0.083 0.002 0.013 Pep 6J29 2016 166 0.090 0.394 0.202 0.873 0.997 0.432 Pep 6J29 2016 175 0.053 0.705 0.336 0.172 0.160 0.571 Pep 6J29 2016 182 0.034 0.458 0.225 0.024 0.225 0.175 Pep 6J29 2016 189 0.035 0.540 0.274 0.653 0.685 0.932 Pep 6J29 2016 194 0.044 0.580 0.301 0.687 0.799 0.786 Pep 6J29 2016 202 0.048 0.841 0.455 0.780 0.293 0.406 Pep 6J29 2016 208 0.013 0.625 0.246 0.291 0.138 0.136 Pep 6J29 2016 216 0.014 0.661 0.258 0.443 0.872 0.959 Pep 6J29 2016 230 0.049 0.942 0.562 0.324 0.117 0.132 Pep 6J29 2016 238 0.040 0.579 0.852 0.383 0.002 0.014 Pep 6J29 2016 246 0.066 0.469 0.976 0.002 0.089 0.039 Pep 6J29 2016 251 0.105 0.372 0.789 0.008 0.228 0.161 Pep 6J29 2016 266 0.261 0.329 0.629 0.000 0.409 0.044 Pep 6J29 2016 274 0.246 0.211 0.485 0.657 0.085 0.127 Pep 6J29 2017 72 0.228 0.234 0.530 0.163 0.921 0.848 Pep 6J29 2017 117 0.217 0.049 0.227 0.926 0.317 0.322 Pep 6J29 2017 125 0.220 0.028 0.165 0.907 0.358 0.193 Pep 6J29 2017 130 0.201 0.030 0.170 0.120 0.422 0.492 Pep 6J29 2017 137 0.241 0.010 0.070 0.182 0.000 0.001 Pep 6J29 2017 144 0.291 0.007 0.054 0.069 0.117 0.044 Pep 6J29 2017 152 0.393 0.003 0.026 0.010 0.006 0.002 Pep 6J29 2017 159 0.491 0.002 0.015 0.029 0.299 0.153 Pep 6J29 2017 166 0.908 0.001 0.005 0.000 0.008 0.000 Pep N38 2015 78 0.960 0.874 0.490 NA NA NA Pep N38 2015 131 0.960 0.959 0.558 0.980 0.521 0.976 Pep N38 2015 141 0.986 0.962 0.573 0.506 0.993 0.673 Pep N38 2015 148 0.905 0.922 0.520 0.045 0.594 0.261 Pep N38 2015 155 0.934 0.775 0.694 0.591 0.021 0.723 Pep N38 2015 162 0.946 0.818 0.718 0.054 0.370 0.932
Pep N38 2015 169 0.715 0.565 0.978 0.000 0.006 0.143 Pep N38 2015 190 0.269 0.186 0.467 0.000 0.000 0.038 Pep N38 2015 198 0.119 0.429 0.680 0.000 0.643 0.925 Pep N38 2015 203 0.057 0.031 0.099 0.001 0.043 0.028 Pep N38 2015 217 0.021 0.013 0.037 0.013 0.005 0.003 Pep N38 2015 267 0.000 0.006 0.007 0.000 0.015 0.003 Pep N38 2016 29 0.000 0.006 0.006 0.911 0.459 0.750 Pep N38 2016 96 0.000 0.006 0.006 0.211 0.978 0.898 Pep N38 2016 112 0.000 0.006 0.006 NA NA NA Pep N38 2016 120 0.000 0.004 0.006 0.614 0.014 0.005 Pep N38 2016 124 0.000 0.004 0.004 0.678 0.688 0.560 Pep N38 2016 133 0.000 0.006 0.006 0.646 0.049 0.124 Pep N38 2016 140 0.000 0.006 0.006 0.656 0.566 0.009 Pep N38 2016 146 0.000 0.006 0.006 0.826 0.580 0.017 Pep N38 2016 153 0.000 0.005 0.005 0.231 0.107 0.002 Pep N38 2016 158 0.000 0.005 0.004 0.034 0.005 0.008 Pep N38 2016 166 0.000 0.004 0.004 0.179 0.052 0.006 Pep N38 2016 175 0.000 0.003 0.004 0.135 0.110 0.008 Pep N38 2016 182 0.000 0.005 0.005 0.084 0.479 0.061 Pep N38 2016 189 0.000 0.004 0.004 0.831 0.719 0.046 Pep N38 2016 194 0.000 0.008 0.008 0.910 0.204 0.666 Pep N38 2016 202 0.000 0.007 0.006 0.036 0.562 0.038 Pep N38 2016 208 0.000 0.008 0.008 0.065 0.951 0.183 Pep N38 2016 216 0.000 0.008 0.007 0.329 0.383 0.238 Pep N38 2016 230 0.000 0.040 0.026 0.471 0.007 0.182 Pep N38 2016 238 0.000 0.067 0.031 0.345 0.071 0.499 Pep N38 2016 246 0.001 0.073 0.037 0.000 0.662 0.623 Pep N38 2016 251 0.002 0.051 0.023 0.371 0.359 0.209 Pep N38 2016 266 0.004 0.122 0.084 0.236 0.187 0.083 Pep N38 2016 274 0.003 0.182 0.108 0.184 0.093 0.260 Pep N38 2017 72 0.003 0.240 0.126 0.283 0.139 0.272 Pep N38 2017 117 0.002 0.232 0.129 0.119 0.964 0.891 Pep N38 2017 125 0.002 0.287 0.157 0.595 0.236 0.204 Pep N38 2017 130 0.002 0.236 0.139 0.648 0.125 0.133 Pep N38 2017 137 0.002 0.342 0.170 0.264 0.140 0.328 Pep N38 2017 144 0.002 0.625 0.306 0.132 0.107 0.208 Pep N38 2017 152 0.002 0.729 0.628 0.052 0.013 0.021 Pep N38 2017 159 0.002 0.699 0.655 0.170 0.719 0.915 Pep N38 2017 166 0.010 0.275 0.841 0.000 0.000 0.000 Serre Carpaccio 2016 91 NA NA NA NA NA NA Serre Carpaccio 2016 117 0.466 0.025 0.099 0.499 0.039 0.021 Serre Carpaccio 2016 124 0.566 0.269 0.389 0.883 0.337 0.858 Serre Carpaccio 2016 131 0.385 0.606 0.527 0.300 0.640 0.654 Serre Carpaccio 2016 140 0.944 0.804 0.832 0.277 0.960 0.987 Serre Carpaccio 2016 147 0.800 0.838 0.796 0.396 0.967 0.810 Serre Carpaccio 2016 152 0.628 0.737 0.647 0.448 0.924 0.575 Serre Carpaccio 2016 159 0.080 0.164 0.107 0.000 0.006 0.003 Serre Carpaccio 2016 166 0.028 0.249 0.124 0.101 0.961 0.420
Serre Carpaccio 2016 174 0.009 0.775 0.487 0.111 0.701 0.727 Serre I214 2016 91 NA NA NA NA NA NA Serre I214 2016 117 0.392 0.987 0.666 0.413 0.627 0.751 Serre I214 2016 124 0.179 0.747 0.745 0.141 0.632 0.787 Serre I214 2016 131 0.660 0.275 0.461 0.222 0.135 0.265 Serre I214 2016 140 0.239 0.854 0.962 0.009 0.463 0.720 Serre I214 2016 147 0.369 0.629 0.608 0.278 0.161 0.417 Serre I214 2016 152 0.329 0.646 0.656 0.642 0.943 0.857 Serre I214 2016 159 0.005 0.573 0.332 0.000 0.724 0.059 Serre I214 2016 166 0.000 0.156 0.051 0.312 0.062 0.006 Serre I214 2016 174 0.000 0.193 0.078 0.504 0.597 0.445 Serre 6J29 2016 91 NA NA NA NA NA NA Serre 6J29 2016 117 0.444 0.090 0.180 0.367 0.081 0.262 Serre 6J29 2016 124 0.440 0.065 0.050 0.183 1.000 0.050 Serre 6J29 2016 131 0.257 0.011 0.014 0.249 0.265 0.024 Serre 6J29 2016 140 0.317 0.018 0.027 0.711 0.086 0.037 Serre 6J29 2016 147 0.541 0.098 0.133 0.732 0.622 0.558 Serre 6J29 2016 152 0.476 0.295 0.308 0.388 0.250 0.947 Serre 6J29 2016 159 0.003 0.131 0.025 0.000 0.319 0.006 Serre 6J29 2016 166 0.000 0.064 0.011 0.000 0.111 0.019 Serre 6J29 2016 174 0.000 0.129 0.001 0.215 0.363 0.553 Serre N38 2016 91 NA NA NA NA NA NA Serre N38 2016 117 0.283 0.717 0.427 0.283 0.632 1.000 Serre N38 2016 124 0.612 0.154 0.254 0.809 0.116 0.227 Serre N38 2016 131 0.942 0.232 0.375 0.482 0.602 0.499 Serre N38 2016 140 0.964 0.132 0.299 0.827 0.396 0.321 Serre N38 2016 147 0.985 0.174 0.415 0.842 0.751 0.746 Serre N38 2016 152 0.988 0.061 0.229 0.903 0.069 0.096 Serre N38 2016 159 0.318 0.224 0.741 0.000 0.167 0.088 Serre N38 2016 166 0.081 0.193 0.850 0.002 0.713 0.912 Serre N38 2016 174 0.029 1.000 0.452 0.306 0.368 0.309
Annexe 2 : Tableau récapitulatif des valeurs de p-value pour chaque type de culture (Pep = pépinière ; ou serre),
génotype (Carpaccio ; I214 ; 6J29 ; N38), année (2015 ; 2016 ; 2017). Les valeurs de p-value ont été calculées pour la hauteur (Hpval), le diamètre (Dpval), le volume (Vpval), le taux de croissance en hauteur (RHpval), le taux de croissance en diamètre (RDpval) et le taux de croissance en volume (RVpval). Les valeurs manquantes sont représentées par NA.
Résumé
L’expérience menée en pépinière repose sur la réponse de deux génotypes de peuplier noir (N38 et 6J29) et de deux génotypes de peupliers euraméricains (Carpaccio et I214) à une contrainte de sécheresse. Un suivi de croissance en hauteur et en diamètre a été réalisé et les résultats ont mis en évidence une meilleure croissance des peupliers euraméricains (en particulier du génotype Carpaccio) par rapport aux peupliers noirs. Les conclusions de cette expérimentation ont été mises en relation avec les années précédentes (2015 et 2016). L’analyse de ces résultats a aussi été mise en parallèle avec une expérience effectuée en 2016 sur les mêmes génotypes sur des peupliers en serre. Les résultats ont montré une différence claire entre les deux modes de cultures.