A novel method for assessing adherent single-cell stiffness in tension: design and testing of a substrate-based live cell functional imaging device
Texte intégral
Figure

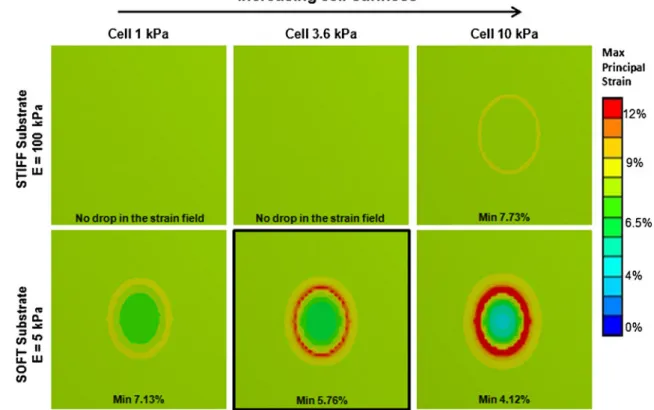
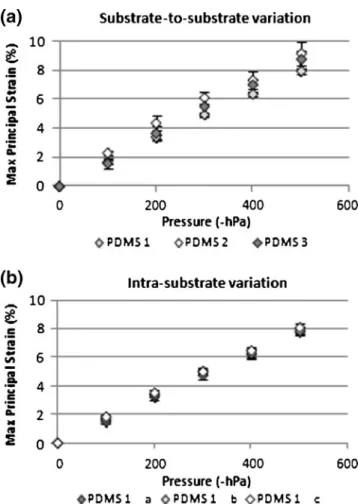
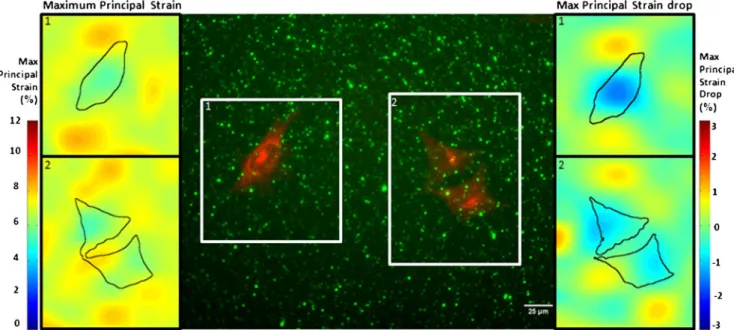
Documents relatifs
We applied FunRes to 177 cell types in 10 different tissues and demonstrate that the detected states cor- respond to known functional cell states of various immune and non-immune
Our genomic analysis in one mouse lemur individual and cDNA sequencing in three individuals (two mouse lemurs and one black- and-white ruffed lemur) revealed that ‘lower’
Aurora-A transβript anγ protein levels were signifiβantly inβreaseγ in high Fuhrman graγe tumours anγ in VHLwt/wt tumours.. These results suggest that Aurora-A
(d) Cell count time courses of two spots containing an siRNA (MCO_0020444) targeting the poorly characterised gene CCDC9. The overall high proportion of mitotic cells, compared to
In the result session I will introduce four studies that explicitly quantify nuclear interactions at the single molecule level in the nucleus of human cells,
The inhibition of energy flux shown by the decreased deep photospheric temperatures relative to the quiet Sun appears re- lated to the reduced magnetic heating with increasing
The pituitary gland has traditionally been regarded as a rather static organ, but using mul- tiphoton time-lapse imaging methods, we clearly show the existence of cell motility in
To this end, infor- mation on the firm-level adoption for seventeen technology applications has been grouped in four broader categories of technologies, which in turn have
