HAL Id: tel-01991687
https://tel.archives-ouvertes.fr/tel-01991687
Submitted on 24 Jan 2019
HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Économie de pêche au Mésolithique et diversité piscicole
à l’Holocène ancien dans le bassin hydrographique du
Doubs
Deborah Frontin
To cite this version:
Deborah Frontin. Économie de pêche au Mésolithique et diversité piscicole à l’Holocène ancien dans le bassin hydrographique du Doubs. Archéologie et Préhistoire. Université Panthéon-Sorbonne - Paris I, 2017. Français. �NNT : 2017PA01H007�. �tel-01991687�
Université de Paris I Panthéon-Sorbonne
UFR 03 Histoire de l’Art et Archéologie – Ecole doctorale d’Archéologie
Economie de pêche au Mésolithique et diversité piscicole à
l'Holocène ancien dans le bassin hydrographique du
Doubs
Thèse présentée et soutenue publiquement pour obtenir le grade de Docteur de l’Université Paris I Panthéon-Sorbonne en Archéologie Environnementale, spécialité Archéozoologie
Par Déborah FRONTIN
Sous la direction de Christophe PETIT, Professeur à l’Université Paris I Panthéon-Sorbonne
Le 23 mars 2017
Jury
Christophe PETIT Professeur, Université Paris I Panthéon-Sorbonne. UMR 7041ArScAn – Archéologies Environnementales.
Directeur
Anne BRIDAULT Chargée de recherche au CNRS, HDR.
UMR 7041ArScAn – Archéologies Environnementales.
Co-directrice
Marie-Christine MARINVAL PRAG, Dr. Université Paris I Panthéon-Sorbonne. UMR 7041ArScAn – Archéologies Environnementales.
Tutrice
Christophe CUPILLARD Ingénieur d’études, Service Régional d’Archéologie de Bourgogne-Franche-Comté. UMR 6249 – Laboratoire Chronoenvironnement.
Examinateur
Grégor MARCHAND Directeur de recherche au CNRS, UMR 6566 – CREAAH – Laboratoire Archéosciences, Université de Rennes I.
Rapporteur
Arturo MORALES Professeur Facultad de Ciencias Francisco Tomás y Valiente, Madrid, Espagne
Rapporteur
2 Université de Paris I Panthéon-Sorbonne
UFR 03 Histoire de l’Art et Archéologie – Ecole doctorale d’Archéologie
Economie de pêche au Mésolithique et diversité piscicole à
l'Holocène ancien dans le bassin hydrographique du
Doubs
Thèse présentée et soutenue publiquement pour obtenir le grade de Docteur de l’Université Paris I Panthéon-Sorbonne en Archéologie Environnementale, spécialité Archéozoologie
Par Déborah FRONTIN
Sous la direction de Christophe PETIT, Professeur à l’Université Paris I Panthéon-Sorbonne
Le 23 mars 2017
Jury
Christophe PETIT Professeur, Université Paris I Panthéon-Sorbonne. UMR 7041ArScAn – Archéologies Environnementales.
Directeur
Anne BRIDAULT Chargée de recherche au CNRS, HDR.
UMR 7041ArScAn – Archéologies Environnementales.
Co-directrice
Marie-Christine MARINVAL PRAG, Dr. Université Paris I Panthéon-Sorbonne. UMR 7041ArScAn – Archéologies Environnementales.
Tutrice
Christophe CUPILLARD Ingénieur d’études, Service Régional d’Archéologie de Bourgogne-Franche-Comté. UMR 6249 – Laboratoire Chronoenvironnement.
Examinateur
Grégor MARCHAND Directeur de recherche au CNRS, UMR 6566 – CREAAH – Laboratoire Archéosciences, Université de Rennes I.
Rapporteur
Arturo MORALES Professeur Facultad de Ciencias Francisco Tomás y Valiente, Madrid, Espagne.
Rapporteur
3
R
EMERCIEMENTSEn premier lieu, je souhaite remercier Christophe Petit, Professeur à l’Université de Paris I Panthéon-Sorbonne qui a accepté de diriger ce travail. Je le remercie pour ses relectures, ses conseils, ses encouragements et son intérêt pour mon travail depuis notre rencontre et tout au long de sa réalisation.
Merci également à Joëlle Burnouf, professeure à l’Université Paris I, Panthéon-Sorbonne pour avoir dirigé la première partie de ce travail avant son départ en retraite.
Mes plus sincères remerciements vont à Anne Bridault, chargée de recherche au CNRS et HDR, pour son encadrement et sa participation au jury de cette thèse. Ses relectures et ses conseils avisés m’ont permis de mener à bien ce travail. Je la remercie également pour son aide lors de la réalisation du projet de collection de référence de l’équipe Archéologies
Environnementales de la MAE.
Je remercie également Christophe Cupillard, ingénieur d’étude au SRA de Bourgogne-Franche-Comté, qui m’a permis de travailler sur les séries du Jura et en particulier sur l’ensemble de l’abri des Cabônes depuis mon Master. Qu’il trouve également ici toute ma reconnaissance pour son accueil chaleureux à Besançon lors de mes nombreuses missions sur le « terrain », pour ses relectures, sa disponibilité et ses conseils tout au long de la réalisation de ce travail et jusqu’à mes demandes de dernière minute. Merci enfin d’avoir accepté de juger ce travail.
Je remercie sincèrement Marie-Christine Marinval, professeure agrégée à l’Université de Paris I Panthéon-Sorbonne pour son encadrement, son soutien et sa présence bienveillante depuis mon arrivée à Paris en Master. Je la remercie également d’avoir accepté de participer au jury de ce travail.
Je remercie vivement Monsieur Grégor Marchand, directeur de recherche au CNRS, UMR 6566 – CREAAH – Laboratoire Archéosciences, Université de Rennes I, de me faire l’honneur d’être rapporteur de cette thèse et de juger ce travail. Qu’il reçoive ici toute ma gratitude.
Je remercie Monsieur Arturo Morales, Professeur à la Facultad de Ciencias Francisco Tomás y Valiente, Madrid, d’avoir accepté d’être rapporteur de ce travail et de participer au jury de cette thèse. Que lui soient exprimés ici mes plus sincères remerciements.
4 Merci également à toute l’équipe du Service Régional de l’Archéologie de Besançon pour leur accueil chaleureux et leur aide dans mes recherches, en particulier Marie-Hélène Chenevoy et Claire Jounin.
Merci à François Meunier, professeur émérite au Muséum National d’Histoire Naturelle, Laboratoire d’Ichtyologie Générale et Appliquée, pour ses conseils et sa formation lors de mon étude squelettochronologique.
Merci à Roland Libois de l’Université de Liège, Unité de Recherches Zoogéographiques, pour son aide lors de mon étude des agents accumulateurs et en particulier pour l’acquisition des pelotes de rejection de martin-pêcheur et des épreintes de loutre.
Merci à Dorothée G. Drucker, Université de Tübingen, pour les analyses isotopiques réalisées sur le matériel de Ranchot et pour son aide dans mes recherches bibliographiques.
Merci aux professionnels et particuliers ayant mis à ma disposition des données précieuses et des spécimens de poisson, en particulier au personnel du SD39 de l’ONEMA (Guy Durand, Patrick Gindre, Philippe Chanteloube, Stephane Forgeois DR2 ONEMA Bourg-Achard). Merci également à Mehdi El Bettah Chargé de Mission à la Fédération du Jura pour la Pêche et la Protection du Milieu Aquatique, à Philippe Falcy de l’association des pêcheurs sportifs de la vallée de la Joux et à Olivier Sommen pisciculteur et membre du programme saumon 2000 au sein de la Réserve Naturelle de la Petite Camargue Alsacienne.
Merci à Mickaël Bejean, responsable de l’aquarium de la Citadelle à Besançon avec une pensée émue pour Jean-Yves Robert, attaché de conservation en charge des collections de zoologie de la Citadelle à Besançon, trop tôt disparu.
Merci à Laure Fontana et Sophie Fornage-Bontemps pour leur aide dans mes recherches bibliographiques ou iconographiques.
Merci aux membres de l’Archaeological Fish Resource (Universités de Bradford, Cambridge, Bournemouth et York) de m’avoir permis de participer aux stages d’archéo-ichtyologie en Angleterre et pour leur aide lors de mes recherches en particulier Andrew Jones, Hannah Russ, Shelia Hamilton-Dyer et Rebecca Nicholson.
Merci aux étudiants et doctorants de la MAE et d’ailleurs avec qui j’ai partagé un bout de chemin au cours de ces années de thèse et en particulier Sarah Beslagic (j’espère qu’un jour on ira élever des chèvres dans les montagnes), Aurélien Creuzieux (la bande à Picsou), Christophe Wardius (au taquet ! et merci pour ton soutien et ton aide technique), Romain
5 Malgarini (merci pour les transports !), Annelise Binois (Merci pour tout, tout, tout, on en discute plus tard parce que c’est trop long), Charlotte Leduc (pour nos discussions, tes encouragements, la bibliographie), Aurélia Borvon (ton accueil à Nantes, la formation que tu m’as dispensé, ton aide, tes relectures, tes conseils, j’en oublie sûrement). Les petits nouveaux du bureau que j’ai connus plus tard mais qui sont parfois devenus grands avant moi : Alain, Jérôme, Lydie, Clémence, Jimena, à qui je souhaite bon courage et bonne continuation.
Merci à ceux qui m’ont aidé lors de la constitution de la collection de référence malgré les odeurs et les visions d’horreur : Anne Guillard, Aude Boyenval, Marie-Pierre Buscail, Cyrille, Virginie Labonne, Emilie Guillaud, Samantha, Sarah Beslagic (hé oui encore toi !).
Merci à Philippe Béarez et Emilie Guillaud du MNHN pour leur aide et les discussions que nous avons partagées.
Merci à toute l’équipe du collège Georges Brassens de Narbonne et en particulier à Mme Cané, principale du collège, pour son soutien et son intérêt pour ma recherche dès notre rencontre. Merci à elle pour la mise à disposition d’un bureau où j’ai pu travailler dans une ambiance de sérieux. Merci à l’équipe vie scolaire pour leur bonne humeur et leur soutien dans mes recherches en particulier Marianne, Sabrina, Blandine, Hélène et Jeremy. Merci aux personnels (enseignants, CPE, administratifs et autres) avec qui j’ai pu discuter et me détendre au cours de ces deux années passées au collège : M. Asseline, M. Sauniere, Mme Dressayre, Gilles Pittatore, Maria Gomez-Latorre, Didier Aribaud, Fabienne Rouffia, Wladimir Badia, Emeline, Delphine Ruquet.
Merci à mes amis français, marocains et sénégalais pour leur aide et leur soutien : Zineb, Freddy, Emeline Fievez, Philippe, Rajaa et sa famille, Lina, Hafida, Hanane, Myriam, Hady. Merci à toute l’équipe de Beincall : Simohamed, Mustapha, Samir, Khalid, Ghali pour leur soutien, leur bonne humeur et la mise à disposition d’un bureau avec ambiance de travail garantie «viiiiiite patate». Merci à mon équipe de RCDLB : Zeid, Makhtar, Diouf.
Enfin, ces remerciements ne seraient pas complets sans une pensée particulière pour ma famille. Mes parents et ma sœur que je remercie pour leur soutien sans faille et leur présence. Mes grands parents encore présents ou disparus en particulier Mamine pour sa passion. La famille de cœur : Florian et sa famille, mon « oncle et ma tante » Patrice et Dominique, Liliane, Zakaria.
6
S
OMMAIRE Remerciements ... 3 Sommaire ... 6 TOME I ... 8 Introduction ... 9I Contexte général et problématique ... 12
I.1 Cadre et définition du Tardiglaciaire-Holocène ... 12
I.2 Historique et problématique de recherche sur les ichtyofaunes en France pour le Tardiglaciaire-Holocène ... 19
I.3 Choix des sources d’information et détermination des outils d’analyse ... 30
I.4 Synthèse et caractérisation du corpus d’étude ... 41
II Présentation des séries ichtyofauniques dans leur contexte archéologique ... 44
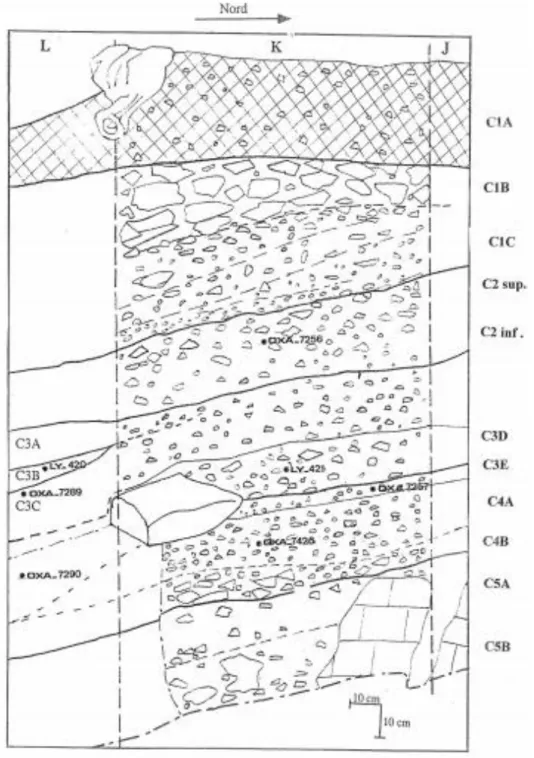
II.1 Le site de l’abri des Cabônes à Ranchot ... 46
II.2 Les autres sites étudiés ... 55
II.3 Synthèse ... 78
III Méthodologie ... 80
III.1 Tri des vestiges et constitution des ensembles étudiés ... 80
III.2 La diagnose des espèces ... 83
III.3 La quantification ... 109
III.4 La reconstitution des tailles et des poids de chair ... 112
III.5 Méthodologie appliquée à l’analyse taphonomique ... 115
III.6 Méthodologie appliquée à l’analyse économique ... 146
IV Résultats et interprétations ... 159
IV.1 Les spectres ichtyofauniques ... 159
IV.2 Origine de l’accumulation des vestiges ... 210
IV.3 Analyse économique ... 244
Conclusion générale ... 273
Bibliographie ... 276
Liste des tableaux ... 296
Liste des figures ... 303
7 Table des matières ... 311 TOME II ... 317 ANNEXES ... 318
8
9
I
NTRODUCTIONLa transition climatique Tardiglaciaire-Holocène a profondément bouleversé l’ensemble des composantes de l’environnement, depuis le milieu physique jusqu'à ses dimensions biologiques, végétales et animales (Bravard, Magny 2002, Bridault 1998, Bridault 2001, Bridault, Chaix 2002, Bridault, Chaix 2008, Bridault et al. 2000, Bridault et al. 2004, Brugal et al. 2004, Cupillard 2010, Magny 1993, Magny 1995, Magny 1998, Magny 2004, Magny et al. 2000, Rotillon 2000, Wall et al. 2010). Ces changements se manifestent entre autres par la transformation des paysages - disparition des glaciers, évolution des cours d’eau - et par le développement d’un milieu forestier de plus en plus fermé, qui s'accompagne d'une recomposition des cortèges de faune. Il semblerait donc inévitable que les sociétés humaines confrontées à ce réchauffement climatique aient dû adapter leurs économies de subsistance à ces nouvelles conditions environnementales (Binford 1968, Clark 1967, Flannery 1969, Zvelebil 1990).
Pourtant, le mode de vie des populations qui se succèdent depuis le Paléolithique final jusqu'au Mésolithique semble en apparence constant, avec des groupes humains mobiles vivant de la chasse, de la pêche et de la collecte. La recherche archéologique et plus particulièrement archéozoologique s'est donc de longue date attachée à éclairer cette apparente contradiction, notamment via l’étude des stratégies de subsistance et en particulier de l’acquisition et de l’exploitation des denrées alimentaires. La compréhension du mode d'exploitation des ressources animales (espèces ciblées, chasse spécialisée ou diversifiée) permet en effet d’approcher l’économie des groupes humains par la compréhension de l'utilisation des territoires, de la fonction des sites, de la gestion de la saisonnalité des ressources, facteurs constituant autant de critères discriminants utiles à la définition des cultures humaines et de leur évolution au cours de la Préhistoire. Le matériel osseux, témoin direct de ces pratiques, est donc un objet d’étude privilégié pour répondre à la question d’éventuelles modifications se produisant au sein de ces économies de subsistance.
Si la question de la chasse est aujourd’hui assez bien documentée (Bridault 1993, Bridault 1997a, Bridault, Fontana 2003, Costamagno, Laroulandie 2004, Fontana 1999, Fontana, Brochier 2009), il n’en va pas de même pour la pêche qui semble pourtant pratiquée depuis le début du Paléolithique (Cleyet-Merle 1990, Le Gall 1984a, Le Gall 1984b, Le Gall 1991, Le Gall 1992, Le Gall 1993, Le Gall 1998, Le Gall 2003a). Malgré un développement récent de l'archéo-ichtyologie ces dernières décennies, le rôle qu'a pu jouer la pêche dans les économies de subsistance à la Préhistoire ainsi que ses modalités pratiques demeurent des questions ouvertes. Son évolution sur le temps long de ce changement climatique reste également sujette à débat : en diminution constante selon les uns (Le Gall 1991, Le Gall 1992, Le Gall 1993, Le Gall 1998), en augmentation selon les autres (Binford 1968, Clark 1967, Flannery
10 1969, Zvelebil 1990), ou demeurant stable selon les troisièmes (Drucker, Bocherens 2001, Drucker et al. 2005, Schulting, Richards 2001, Valentin, Drucker 2009, Valentin, Drucker 2010). Ce travail cherche donc à apporter de nouveaux éléments concernant la pratique de la pêche et son évolution au cours de la transition Tardiglaciaire-Holocène par le biais de l'exploitation de restes d'ichtyofaune fossile.
Répondre à une telle problématique demande toutefois la mise en place en amont d’une méthodologie adaptée pour chacune des trois étapes de l'analyse : la récolte des données, l'exploitation des résultats et la comparaison avec des contextes pertinents. La bibliographie s'avère en effet insuffisante sur la détermination de l'origine des accumulations de vestiges, sur certains aspects de l'identification spécifique, tandis qu'aucun mode de quantification des restes ne fait l'unanimité parmi les chercheurs. Ce travail de recherche implique donc de faire un tri dans l'existant, de compléter les référentiels qui l'exigent et d'en développer de nouveaux le cas échéant. Ces étapes préliminaires réalisées, il devient alors possible par l'exploitation des données recueillies de déterminer tant les caractéristiques de chaque contexte (choix des espèces pêchées, des milieux de pêche, des techniques de pêches, etc.) que d'envisager une analyse plus globale de l’activité de pêche à différentes échelles de temps et d’espace. Un regard économique pourra également être porté sur ces résultats, par l'emploi des outils méthodologiques développés pour la chasse des mammifères et des oiseaux (Bridault 1993, Bridault 1997a, Bridault, Fontana 2003, Costamagno, Laroulandie 2004, Fontana 1999, Fontana, Brochier 2009).
Reconstituer les modalités d’évolution de la pratique de la pêche à travers le temps et tenter d’interpréter les éventuels changements repérés étant un objectif assez conséquent, il est également important de choisir judicieusement la zone d’étude et les séries à analyser. La logique de l'étude des ichtyofaunes impose un travail au niveau du bassin versant, unité hydrographique cohérente. Les séries retenues pour ce travail proviennent toutes du bassin du Doubs dans le massif jurassien, avec des sites localisés soit directement en bordure du Doubs, soit le long du Dessoubre, l’un de ses principaux affluents. Cette zone d’étude présente l’avantage de livrer des ensembles d’ichtyofaunes datés du début de l’Epipaléolithique jusqu’à la fin du Mésolithique, parmi lesquels une série inédite, celle de la couche 3 de l’abri des Cabônes à Ranchot (Jura), qui est, avec sa dizaine de milliers de restes, l'une des plus importantes du territoire français pour le Mésolithique. L’étude de cette série constitue la colonne vertébrale de ce travail ; la séquence temporelle et régionale est ensuite complétée par neuf autres séries en provenance de trois sites, Rochedane à Villars-sous-Dampjoux (5 séries de l'Epipaléolithique au Mésolithique), Gigot I à Gigot (2 séries mésolithiques) et la Roche-aux-Pêcheurs à Villers-le-Lac (2 séries mésolithiques).
Le plan de ce mémoire de thèse s'organise comme suit. Dans la première partie de ce travail, je poserai les bases du contexte de recherche, présentant le cadre temporel et géographique de
11 l’étude puis dressant un historique de la recherche en archéo-ichtyologie sur la pratique et l'évolution de la pêche au cours de la transition Tardiglaciaire-Holocène ancien. Je justifierai ensuite le choix des sources d’information retenues, des séries étudiées et des outils d’analyse utilisés. Une seconde partie sera consacrée à la présentation des ensembles fauniques dans leur contexte archéologique. Chaque site et chaque série feront l’objet d’une présentation détaillée permettant de comprendre les modalités de leur intégration au sein d’un corpus d'apparence hétérogène, notamment en ce qui concerne les nombres de restes. La troisième partie sera alors consacrée à la méthodologie. Les méthodes de base utilisées ainsi que les apports et compléments réalisés au cours de ce travail y seront détaillés pour chaque partie de l’analyse et chaque méthode. Enfin, les résultats et interprétations réalisés à l’aide des études archéo-ichtyologiques, taphonomiques et économiques seront présentés au sein de la dernière partie du travail avant de conclure.
12
I Contexte général et problématique
Aborder la question de la pratique de la pêche au cours de la transition Tardiglaciaire-Holocène demande de définir précisément le cadre paléoclimatique, paléoenvironnemental et chronoculturel de la période. En effet, il s’agit d’une fenêtre de temps au cours de laquelle se déroulent de profonds changements sur le plan climatique et environnemental mais également dans la structuration et le fonctionnement des sociétés humaines, en particulier en ce qui concerne les modes de subsistance. Afin de cerner la question de la pratique de la pêche et de son rôle dans l’alimentation humaine au cours d’une période de changements aussi importants, il sera essentiel de restituer le contexte de recherche. En effet, ce travail part de plusieurs hypothèses que nous présenterons au cours des prochains chapitres et qui servent de base aux réflexions engagées ici. Ainsi, au cours de cette première partie, après avoir défini le cadre paléoclimatique, paléoenvironnementalet chronoculturel de la période, nous présenterons la problématique archéo-ichtyologique dans son contexte de recherche avant de définir le choix du corpus d’étude. L’Est de la France et ses données seront plus particulièrement abordés, et nous reviendrons également sur le choix de cette région comme zone d’étude au cours de cette partie.
I.1 Cadre et définition du Tardiglaciaire-Holocène
La transition Tardiglaciaire-Holocène est une période très importante dans la compréhension de l’histoire de l’humanité puisqu’il s’agit du dernier réchauffement climatique majeur connu par l’Homme. L’étude des stratégies mises en place par les hommes en ce qui concerne l’occupation des territoires et les modalités de subsistance pour traverser cette période de grands changements n’en est donc que plus intéressante. Au cours de cette présentation, la chronologie globale a été assortie des repères chronologiques locaux (oscillations lacustres, nomenclature alpine). Dans la mesure du possible les dates indiquées sont présentées en datation calibrée exprimées en cal BC à partir du logiciel de calibration en ligne Calib 7.1.
I.1.1 Cadre chronologique et climatique
La transition Tardiglaciaire-Holocène est fixée par les travaux récents à la fin du Pleistocène et au début de l’Holocène vers 9700 cal BC (Cupillard et al. 2015). Il s’agit d’une période pendant laquelle se déroulent de nombreuses oscillations climatiques qui accompagnent la déglaciation würmienne ; puis l’adoucissement du climat qui caractérise l’Holocène. Elles correspondent dans l’hémisphère nord au recul des glaciers, à une élévation du niveau des mers et au développement progressif de la forêt. Grâce aux nombreux travaux réalisés sur les
13 dépôts fluviatiles, lacustres et palustres dans l’Est de la France et dans le nord de l’Europe (Bravard, Magny 2002, Magny 1993, Magny 1995, Magny 1998, Magny 2004, Magny et al. 2006, Magny et al. 2000), il s’agit d’une période bien connue de l’histoire climatique terrestre, ce qui permettra d’offrir un cadre solide à ce travail.
En France, le climat au cours du Tardiglaciaire-Holocène se caractérise par plusieurs phases d’amélioration et de détérioration qui ont pu être directement reliées à des évènements plus globaux (Fig. 1). Par exemple la « phase du Locle » identifiée dans l’est de la France correspond en fait à l’échelle mondiale à une longue détérioration climatique appelée « évènement de 8200 cal BP » (Magny 1995, Magny 2004). L’étude du contenu des séquences d’Europe du nord en particulier les données polliniques ont également permis la mise en place d’un cadre biostratigraphique pour la période. Ces études ont menées à l’établissement de zones polliniques aussi appelées biozones ou chronozones dont la définition et le calage chronologique pour la période sont les suivants :
Pour le Tardiglaciaire se succèdent le Dryas ancien (16 000-12 700 cal BC), le Bölling (12 700-12 100 cal BC), le Dryas moyen (12 100-11900 cal BC), l’Alleröd (11 900-11 000 cal BC) et le Dryas récent (10 900-9750 cal BC). Au cours de ces biozones, le climat est très instable (Bégeot et al. 2006, Cupillard 2010, Cupillard, Richard 1998, Magny et al. 2006, Peyron et al. 2005, Richard et al. 1998)
Pour le début de l’Holocène se succèdent le Préboréal (9750-8000 cal BC), le Boréal (8000-7000 cal BC) et l’Atlantique ancien ((8000-7000-4900 cal BC). Cet interstade tempéré dans lequel nous nous trouvons encore aujourd’hui correspond, à ses débuts, à une augmentation des températures et de l’humidité et à l’extension des forêts déjà initiée au cours de l’Alleröd (Bégeot et al. 2006, Cupillard 2010, Cupillard, Richard 1998, Magny et al. 2006, Peyron et al. 2005, Richard et al. 1998)
Afin de poser le cadre de cette étude, je présenterai les chronozones qui couvrent la fenêtre de temps qui nous intéresse soit la séquence qui s’étend de l’Alleröd (fin du Tardiglaciaire) à l’Atlantique ancien (dernières économies de subsistance de chasseurs-cueilleurs et passage aux économies de production).
L’Alleröd (11900-11000 cal BC) correspond à une phase tempérée de la fin du Tardiglaciaire caractérisée par l’installation d’un climat doux et humide. La remontée des températures au cours de la période permet l’expansion d’une forêt dominée par le pin et la recolonisation par la végétation de certaines zones occupées précédemment par les glaciers. L’Alleröd est également caractérisé par une augmentation des niveaux marins due à la fonte des glaciers (Magny 1995, Magny et al. 2006, Magny et al. 2000, Peyron et al. 2005, Wall et al. 2010). Le Dryas récent (10 900-9750 cal BC) correspond au dernier épisode froid du Tardiglaciaire. Au cours de cet épisode se produit un refroidissement brutal dû à la descente du front polaire
14 dans l’hémisphère nord et à une perturbation de la circulation thermohaline dans l’Atlantique nord (Cupillard 2010). En France, ce refroidissement entraîne un recul de la forêt et le retour des espèces steppiques. Cette période est divisée en trois phases : deux périodes froides qui encadrent une période plus favorable. Il s’agit d’une période peu propice au peuplement humain [Azilien et Epigravettien récent] (Fornage-Bontemps 2013), en particulier dans les zones d’altitude au cours des périodes froides. Dans le Jura franco-suisse par exemple, les occupations épipaléolithiques ont été repérées uniquement sur les marges du massif (Cupillard 2010, Cupillard et al. 2015).
Le Préboréal (9750-8000 cal BC) correspond à la première biozone de l’Holocène. Il s’agit d’une phase de réchauffement brutal dans l’hémisphère nord (Magny 1998, Magny et al. 2006, Magny et al. 2000, Peyron et al. 2005, Wall et al. 2010) caractérisée par une augmentation des températures hivernales et estivales, une baisse des précipitations et le développement de la forêt jusqu’à 1300 m d’altitude. Au cours de la période, les herbacées héliophiles disparaissent en raison du développement rapide de la forêt de pin. Comme pour le Dryas récent, le Préboréal est divisé en trois phases : deux phases tempérées entourent une phase de détérioration appelée dans le Jura « phase de Remoray ». Au cours de cette période, l’homme se réapproprie le territoire et en particulier les hauteurs. Les témoignages liés à la fréquentation des hauteurs par les chasseurs-cueilleurs du Mésolithique ancien sont présentes dans l’ensemble des massifs montagneux européens (Cupillard 2010).
A la suite du Préboréal, le Boréal (8000-7000 cal BC) correspond à une longue période de détérioration climatique classiquement définie comme l’époque du noisetier. Celle-ci caractérise surtout les deux premiers tiers du Boréal soit de 8020 à 7200 cal BC. Elle culmine entre 7600 et 7200 cal BC et, dans le Jura, elle est appelée « phase de Joux 2 » (Magny 1998, Magny et al. 2006, Magny et al. 2000, Peyron et al. 2005). La fin du Boréal est caractérisée par une amélioration climatique au cours de laquelle le noisetier cède la place à une forêt dense de chêne et d’orme accompagnés du tilleul, du frêne et de l’érable. Cette forêt annonce la chênaie mixte de l’Atlantique. Au cours de cette période, le peuplement humain (Mésolithique moyen) est bien développé et les premiers impacts de l’homme sur le milieu apparaissent, notamment par la mise en évidence dans certaines études polliniques de fluctuations anormales du noisetier (Bégeot 1998).
Enfin, l’Atlantique ancien (7000-4900 cal BC) correspond à la dernière période de la fenêtre de temps qui nous intéresse. Il s’agit de l’optimum climatique de l’Holocène au cours duquel se développe une chênaie mixte. Dans l’Est de la France, l’Atlantique ancien se divise en deux phases. La première, l’Atlantique ancien 1 durant laquelle se déroulent l’optimum climatique Holocène vers 6600 cal BC puis la crise climatique de 6200 cal BC. Elle est suivie d’une deuxième phase, l’Atlantique ancien 2 qui correspond à une longue période de détérioration climatique de 6000 à 4900 cal BC. Cette dernière phase est caractérisée par le
15 développement d’Abies dans le Jura et sur le plan humain par le développement des populations du Mésolithique récent puis par l’arrivée des premiers agriculteurs vers 5400-5300 cal BC [premières traces de pollens de plantes cultivées vers 5800-5700 cal BC] (Cupillard, Richard 1998).
I.1.2 Evolution du peuplement humain et de l’environnement animal
En ce qui concerne le peuplement humain au cours de cette période, l’Est de la France voit se succéder différents groupes possédant des cultures matérielles distinctes. La fin du Tardiglaciaire (fin du Paléolithique) est marquée par le passage des cultures du Magdalénien final aux cultures aziliennes et épigravetienne. L’Azilien se caractérise par une microlithisation croissante des industries lithiques mais les habitudes économiques et technologique restent relativement similaires à celles du Magdalénien (Cupillard 2010, Cupillard, Richard 1998, Cupillard, Richard 1999, David 1996, Thévenin 1981, Thévenin 1998, Thévenin 2000). Au cours de l’Azilien, une différenciation est faite entre l’Azilien ancien, contemporain de l’interstade Bölling-Alleröd, et l’Azilien récent qui se place durant le Dryas récent. Le début de l’Holocène voit, quant à lui, se succéder les populations du Mésolithique (ancien, moyen, récent/final) pendant la période qui s’étend du Préboréal à l’Atlantique. Le Mésolithique se caractérise par la microlithisation de l’outillage et par l’utilisation généralisée de l’arc. Ces changements sont liés, selon les chercheurs, à des changements de comportements d’acquisition des ressources alimentaires animales, à une adaptation à la transformation du milieu et en particulier au développement des forêts (Cupillard 2010, Cupillard, Richard 1998, Cupillard, Richard 1999, David 1996, Thévenin 1981, Thévenin 1998, Thévenin 2000)
Au cours de cette période, l’évolution de l’environnement animal est lui aussi bien connu grâce aux travaux archéozoologiques (Bridault 1998, Bridault 2001, Bridault, Chaix 2002, Bridault, Chaix 2008, Bridault et al. 2000, Bridault et al. 2004, Brugal et al. 2004). La recomposition des cortèges de faunes semble s’amorcer très tôt et s’opérer sur une assez longue durée. Elle s’accélère toutefois dès le Bölling en particulier avec la disparition ou la migration de la plupart des espèces associées à des conditions climatiques rigoureuses (le renne, le mammouth). Après 12 000 cal BC toutefois, certaines espèces comme le cheval sont encore attestées dans les spectres de faunes chassées par les populations aziliennes et sont associées à des espèces liées à des conditions plus tempérées comme le cerf, l’élan, le sanglier ou le chevreuil. Le cheval devient par la suite très rare dans les spectres de chasse du Mésolithique. Enfin, la fin de la période voit l’apparition de spectres mixtes mêlant espèces sauvages et premières espèces domestiques qui témoignent du phénomène de néolithisation (Ghesquière, Marchand 2010).
16 En ce qui concerne les ichtyofaunes, les glaciations du Quaternaire en particulier l’avancée et le retrait des glaciers et dans une moindre mesure l’abaissement des niveaux marins entrainent de nombreuses connexions inter-bassins hydrographiques qui ont permis aux espèces de coloniser de nouveaux cours d’eau (Persat, Keith 1997, Persat, Keith 2002). Ces connexions se trouvent à plusieurs niveaux : entre les grands cours d’eau, entre les petits ruisseaux mais également dans des seuils hydrographiques moins marqués. Un épisode ponctuel, unique ou très court, comme une crue, peut alors permettre le déplacement de certaines espèces. (Persat, Keith 1997). Avant le Tardiglaciaire, seules les espèces cryophiles et se reproduisant à basse température comme les Salmonidés, la lote, le chabot ou encore le brochet, et dans une moindre mesure, certains Cyprinidés rhéophiles comme la vandoise, semblent s’être maintenues (Le Gall 2010). En effet, les conditions sont rigoureuses et les espèces thermophiles comme le gardon disparaissent alors ou se déplacent vers des zones refuges méridionales localisées principalement dans le bassin de la mer noire et plus accessoirement dans le bassin du Golfe du Lion. Dès le Tardiglaciaire et en particulier au moment de la fonte des glaces, les opportunités de recolonisation sont nombreuses et c’est à ce moment là que les peuplements piscicoles connaissent leur maximum d’expansion en utilisant les voies naturelles formées par le Danube et le Rhône. Les espèces cryophiles se dispersent alors rapidement suivies par les espèces thermophiles (Le Gall 2010, Persat 2003, Persat, Keith 1997, Persat, Keith 2002).
I.1.3 Evolution de l’environnement physique
Au cours de la transition Tardiglaciaire-Holocène, l’impact du changement climatique n’est pas seulement perceptible sur les composantes biologiques de l’environnement mais également sur certaines de ses composantes physiques. Ainsi la disparition des glaciers dans les massifs montagneux remodèle le paysage. Les sols d’abord peu épais se développent, protégés de l’érosion par l’expansion de la végétation et en particulier de la forêt (Cupillard et al. 2015). De nombreuses cuvettes voient la création puis l’évolution de lacs plus ou moins étendus qui se comblent progressivement. Ceux-ci au départ assez étendus voient leur taille se réduire, certains disparaissent ou se transforment en tourbières (Cupillard 2010). Les cours d’eau évoluent également au cours de la période. Dans le Jura par exemple, une étude portant sur la Seille, affluent de la Saône, démontre que le changement climatique affecte très tôt la morphologie des cours d’eau qui passent progressivement de larges chenaux en tresse au cours du Dryas récent à des chenaux uniques à méandre au cours du Boréal. Ces changements en premier lieu liés à l’évolution du climat semblent également fortement dépendants du développement du couvert végétal. En effet, l’étude démontre que l’impact du changement climatique sur le cours d’eau est moins important lorsque la forêt est bien développée, cela malgré les nombreuses oscillations enregistrées au cours de la période (Rotillon 2000). Le
17 développement de la végétation, en particulier la végétation rivulaire aurait donc joué un rôle sur la fixation des cours d’eau au cours des périodes récentes.
I.1.4 Synthèse
La présentation réalisée au cours de ce chapitre démontre que ce travail s’inscrit dans un cadre complexe qui voit se dérouler au cours d’une période de temps restreinte des changements majeurs dans l’ensemble des composantes climatiques, physiques et biologiques de l’environnement. Afin de synthétiser de manière claire les modifications qui se déroulent au cours de cette fenêtre de temps et leur chronologie, un schéma de synthèse présentant l’ensemble de ces changements a été réalisé (figure 1 ci-dessous).
18 Figure 1 : Schéma de synthèse du cadre chronoculturel, environnemental et climatique de la transition Tardiglaciaire-Holocène dans le massif jurassien
19
I.2
Historique et problématique de recherche sur les ichtyofaunes en France pour leTardiglaciaire-Holocène
En France, les premières découvertes liées à la pêche dans les sites archéologiques remontent au XIXe siècle mais ce n’est qu’à partir de la deuxième moitié du XXe siècle, en particulier avec l’évolution des méthodes de fouille et de collecte, que l’archéo-ichtyologie va réellement prendre son essort (Cravinho 2009). Les études se limitent aux représentations iconographiques de poissons, aux ustensiles de pêche et aux plus gros restes osseux de poissons mis au jour. Toutefois, dès les années 1960, le développement de la fouille fine et du tamisage des sédiments augmente le nombre et la quantité des découvertes, en particulier pour les restes d’ichtyofaunes. L’archéo-ichtyologie et ses méthodes d’étude se développent alors afin d’exploiter le potentiel informatif de ces nouveaux vestiges. La discipline s’inspire des travaux réalisés sur les mammifères et évolue rapidement grâce aux recherches de nombreux précurseurs en France et ailleurs (Butler 1993, Butler 1996, Butler, Chatters 1994, Butler, Schroeder 1998, Carlson 1988, Casteel 1976, Colley 1984, Colley 1986, Cravinho 2009, Desse 1984, Desse, Desse 1976a, Desse, Desse 1976b, Desse, Desse 1983, Desse 1981, Desse 1980, Desse, Desse-Berset 1992, Desse, Desse-Berset 1998, Desse et al. 1987, Desse et al. 1989, Desse et al. 1990, Desse 1989, Desse 2002, Jones 1984, Jones 1986, Juan-Muns 1984, Lauren et al. 2008, Le Gall 1982, Le Gall 1984a, Le Gall 1984b, Le Gall 2003b, Le Gall 2008, Libois, Hallet-Libois 1988, Libois et al. 1987b, Meunier 1984, Morales, Rosenlund 1979, Nicholson 1992, Nicholson 1993a, Nicholson 1993b, Nicholson 1995, Nicholson 1996, Nicholson 1998, Nicholson 2000, Piquès 1998, Radu 2005, Sternberg 1995, Van Neer, Morales Muniz 1992, Wheeler, Jones 1989, Yee Cannon 1987). Dans un premier temps l’étude des ossements d’ichtyofaunes est menée conjointement à celle des autres types de vestiges (représentations artistiques et ustensiles de pêche). Les premières réflexions sur les économies de pêche s’intéressent tout particulièrement aux périodes préhistoriques et aux poissons de rivière, mais elles vont rapidement s’étendre à l’ensemble des ichtyofaunes et des périodes culturelles. Les principales problématiques abordées au départ sont de deux types :
- Méthodologiques : ayant pour but de développer la discipline et qui s’intéressent aux problématiques taphonomiques (origine des accumulations) aux problèmes d’identification des espèces, de quantification des restes, de reconstitution des tailles et poids des individus, de lecture des âges et des saisons de mort …
- Economiques : ayant pour but de développer la connaissance des économies de pêche et qui s’intéressent au fonctionnement de l’activité (milieux de pêche, techniques de pêche …) et à l’évaluation de son importance au sein des économies de subsistance.
20 Le gros de ces travaux est réalisé sur les restes d’ichtyofaunes qui sont d’ailleurs devenus aujourd’hui l’objet d’étude principal de la discipline, en grande partie grâce au nombre de découvertes qui dépasse largement celles des autres types de vestiges, rares, souvent périssables et qui s’accompagnent la plupart du temps, comme nous le verrons par la suite, de nombreuses problématiques et notamment de difficultés de détermination.
L’archéo-ichtyologie est donc une discipline relativement jeune et certaines des problématiques exposées ci-dessus sont encore aujourd’hui au cœur des réflexions qui l’animent. Ce travail ne fait pas exception puisqu’il apporte un éclairage aussi bien sur des questions méthodologiques qu’économiques et s’inscrit donc dans la continuité des travaux réalisés par les précurseurs français et internationaux.
Pour la période qui nous intéresse, la question de la pratique de la pêche par les sociétés de la fin des temps glaciaires et du début du Postglaciaire est déjà documentée. Toutefois, nous allons voir qu’elle n’en reste pas moins problématique aussi bien en France que dans le reste de l’Europe. Les études récentes qui traitent de cette problématique permettent de dresser trois modèles de pensée très différents qui ont constitué la base des réflexions engagées dans ce travail. Ces études que nous allons présenter en détail ci-après apportent des résultats plus ou moins détaillés et fiables. Toutefois, il est important de préciser qu’elles n’ont pas été réalisées dans les mêmes conditions ni avec les mêmes sources d’information. Certaines sont plus anciennes que d’autres, certaines ne traitent que d’une seule aire géographique ou d’une seule période de temps, certaines se basent exclusivement sur des données qualitatives quand d’autres, plus récentes, compilent des données quantitatives précises. Afin de dresser le contexte de recherche à partir duquel a été réalisé ce travail, je présenterai successivement ici les trois modèles de pensées qui animent la recherche récente en précisant pour chacun d’eux le contexte de recherche qui lui est associé. Une synthèse sera ensuite réalisée afin de préciser la problématique et la direction des recherches qui seront engagées dans la suite de ce travail.
I.2.1 Amplification de la pratique de la pêche ?
Cette première hypothèse est issue de travaux réalisés dans les années 1960-1970 par plusieurs chercheurs dont J.G.D.Clark, L.R. Binford, K. Flannery ou plus tard M. Zvelebil (Binford 1968, Clark 1967, Flannery 1969, Zvelebil 1990) à partir de l’étude de sites du Proche-Orient dont les hypothèses de recherche ont ensuite été transposées à l’Europe tempérée. Ces chercheurs et leurs successeurs ont émis puis discuté l’hypothèse d’un élargissement des ressources exploitées chez les sociétés du début du Postglaciaire : la théorie
21 de la « Broad Spectrum Revolution » (Flannery 1969) [page 74]. Depuis, les travaux se sont succédé, soutenant ou réfutant le bien-fondé de ce modèle de pensée qui avance l’idée que les changements environnementaux qui se sont opérés dès le début du réchauffement climatique holocène auraient entraîné des modifications profondes dans les modes de subsistance des populations humaines. Pendant cette période, vue comme un moment critique de l’histoire humaine, de nombreuses espèces végétales mais également animales peu caloriques auraient alors été exploitées par les hommes afin de parer la pénurie d’espèces habituellement consommées. Ces nouvelles ressources sont en effet vues comme ayant l’avantage d’apporter un complément nutritionnel non négligeable, d’être prévisibles, disponibles en grande quantité, stockables, permettant également de diminuer les risques d’échecs lors de la chasse/collecte et donc par extension les risques de famine.
Ce dernier mot peut paraître misérabiliste mais c’est en fait réellement dans cette optique qu’a été créée cette hypothèse de travail. En effet, au cœur de ce modèle, la recomposition de l’environnement au Tardiglaciaire-Holocène est perçue comme un phénomène négatif, créant une diminution des ressources consommables et ceci dès la fin des temps glaciaires. C’est justement à cause de cette « pénurie » que les populations humaines auraient été obligées de compenser la disparition des ressources habituellement exploitées, notamment en diversifiant la base de leur subsistance. Cette diversification aurait alors été orientée vers de nombreuses espèces habituellement délaissées. Outre les plantes, plusieurs petites espèces animales auraient été exploitées parfois en grande quantité parmi lesquelles les petits mammifères, les oiseaux, les mollusques (coquillages, escargots), les batraciens, les chéloniens ou encore les poissons. Dans ces travaux, ces espèces sont clairement décrites comme des ressources secondaires, de plus bas niveau trophique voire sub-optimales (moins intéressantes du point de vue nutritionnel), mais ayant le mérite d’avoir permis aux hommes de survivre pendant ces temps difficiles.
En France, cette problématique est encore bien présente dans les travaux des chercheurs travaillant sur les périodes qui couvrent la fin de la Préhistoire. En archéozoologie notamment, des travaux récents se sont attelés à étudier la validité de la théorie de la diversification des ressources exploitées à partir des spectres de faunes de sites localisés dans les régions du Jura, des Alpes du Nord et des Pyrénées françaises (Bridault 1993, Bridault 1994, Chaix 1988, Costamagno et al. 2009, Fontana, Brochier 2009). Concernant la chasse des grands mammifères, les travaux précédemment cités démontrent une exploitation plus large de l’environnement, parfois dès le Magdalénien supérieur. Toutefois, les techniques de prédation restent assez similaires et sont orientées vers quelques espèces de haut rang d’utilité économique et notamment vers le grand gibier. En ce qui concerne les petites espèces : mammifères, oiseaux, poissons … les données sont bien souvent plus problématiques. D’une part parce que la quantité de données est faible, d’autre part parce que l’origine anthropique des accumulations est souvent difficile à démontrer. Toutefois, certaines données
22 préliminaires permettent de proposer quelques tendances pour ces petites faunes et en particulier les poissons. Du Paléolithique final au Mésolithique final, une augmentation de l’occurrence des ichtyofaunes semble bien exister dès la fin du Paléolithique (Bridault 1994, Bridault 1997a, Costamagno et al. 2009, Fontana, Brochier 2009). Bien qu’aucune conclusion définitive ne puisse être émise sur ces données qui restent tout de même relativement partielles, il est intéressant de constater que ces observations vont dans le sens des hypothèses émises par L. R. Binford [1983, p. 211] (Binford 1983). Selon cet auteur, l’exploitation des ressources du milieu aquatique et notamment des poissons serait l’option de survie choisie de manière préférentielle par les populations d’Europe tempérée afin de survivre au changement climatique. Certains archéo-ichtyologues français constatent également dans leurs études un regain d’intérêt pour les activités halieutiques qui semble se manifester dès la transition climatique et qui serait lié à la raréfaction puis à la disparition du renne, gibier de prédilection des Magdaléniens (Desse, Desse 1976b) [T1, p. 697-702]. Ces observations iraient donc dans le sens des hypothèses émises dans le cadre du modèle de la « broad spectrum revolution».
I.2.2 Diminution de la pratique de la pêche ?
Cette deuxième hypothèse est issue des travaux d’O. Le Gall qui a réalisé plusieurs synthèses de données archéo-ichtyologiques en utilisant l’occurrence des espèces et le nombre de restes déterminés par espèce dans les spectres d’ichtyofaunes pêchées dans les sites français au cours de la transition Tardiglaciaire-Holocène. Ces synthèses concernent surtout le milieu continental, plus rarement marin et réunissent les données de près de 50 sites soit un peu plus de 70 ensembles répartis sur la période allant du Magdalénien ancien au Néolithique récent (Le Gall 1991, Le Gall 1992, Le Gall 1993, Le Gall 1998). Les études concernent en majorité les sites du sud de la France : le grand Sud-Ouest (vallées de la Garonne, du Lot, de l’Aveyron, le massif des Pyrénées…), la Méditerranée et la vallée du Rhône mais également quelques observations ponctuelles dans le nord de la France (bassin de la Seine, côte atlantique) et dans certains pays d’Europe (Allemagne, Danemark, Portugal).
Les résultats de ces travaux pour la fin du Paléolithique, pendant laquelle le changement climatique s’amorce, montrent un développement de la pratique de la pêche. Celle-ci est toutefois de courte durée. Rapidement, au cours des périodes suivantes le phénomène inverse apparaît. Dès la fin du Paléolithique supérieur, la pratique de la pêche diminue progressivement jusqu’à disparaître quasiment au cours du Mésolithique. Il faut alors attendre la fin du Mésolithique voire la transition Mésolithique/Néolithique pour constater une reprise modérée des activités halieutiques (Le Gall 1991, Le Gall 1992, Le Gall 1993, Le Gall 1998).
Ces tendances sont considérées comme générales à l’échelle de la France, voire de l’Europe, bien qu’il existe parfois quelques exceptions notamment dans des sites septentrionaux : c’est
23 le cas par exemple sur le site Mésolithique du Haut-des-Nachères à Noyen-sur-Seine (Seine-et-Marne). De plus, les périodes d’expansion et d’abandon semblent plutôt liées selon l’auteur à des phénomènes climatiques, environnementaux ou culturels (Le Gall 1993). Ainsi, l'expansion des pêches qui caractérise la période allant de la fin du Magdalénien jusqu’à l’Azilien, où un pic des activités de pêche est enregistré est mise en relation avec les changements engendrés par les évènements froids enregistrés au cours de cette fenêtre de temps (Cravinho 2009, Le Gall 1992, Le Gall 1993). Elle se caractérise par l’augmentation des captures d’espèces de plus bas niveau trophique comme les Cyprinidés qui vont alors rapidement dominer les assemblages de cette période.
Par la suite, une baisse progressive de la pratique de la pêche est enregistrée dès la fin de l'Epipaléolithique et le début du Mésolithique soit dès le Préboréal et jusqu’à la fin du Boréal (Le Gall 1991, Le Gall 1993). Au cours de cette période comme nous l’avons vu, certains sites mésolithiques semblent cependant faire exception à cette tendance générale comme Noyen-sur-Seine en Seine-et-Marne. Selon O. Le Gall, il ne s’agit toutefois que d’un site isolé et particulier. En effet, le site, attribuable au Mésolithique moyen/récent, contemporain de la transition Boréal/Atlantique, possède une position relativement « nordique » qui pourrait expliquer sa parenté avec les sites mésolithiques d’Europe du Nord à forte tradition halieutique. Il apparaît également localisé dans un environnement particulier (en bordure immédiate de la Seine), ce qui pourrait expliquer son caractère exceptionnel (Le Gall 1993). Hormis pour ces sites septentrionaux (Noyen-sur-Seine en France et sites d’Europe du Nord), cette tendance à la baisse est appuyée par des observations faites en Allemagne et au Portugal. Cette évolution négative de la pratique de la pêche semble donc répondre selon Le Gall à une tendance générale localisée en Europe tempérée mais dont l’auteur ne peut expliquer la raison.
Enfin, à la fin de la séquence chronologique, c'est-à-dire à l’extrême fin du Mésolithique voire à la transition du Mésolithique avec le Néolithique, une reprise des activités de pêche est observée par Le Gall 1993. Toutefois, cette augmentation n’atteindra jamais le niveau enregistré au cours de l’Azilien. Selon l’auteur, ce phénomène de reprise est peut-être à mettre en relation avec de possibles déboires expérimentaux ayant accompagné le développement de l’agriculture au Néolithique (Le Gall 1993). A l’image de la période d’amorce du changement climatique, la pêche a pu jouer un rôle de soutien similaire pour les populations néolithiques au cours de cette nouvelle période de changement. Cette hypothèse est soutenue par la quasi-absence de restes d’ichtyofaunes dans les sites dont les habitants ne semblent pas avoir rencontré de problèmes alimentaires particuliers ou dans ceux qui témoignent d’une agriculture bien installée. C'est-à-dire dont les spectres de faunes témoignent d’une forte tradition cynégétique ou encore dans ceux caractérisés par l’apparition des espèces domestiques notamment les plantes [légumineuses] (Le Gall 1993). Selon Le Gall, la pratique de la pêche au cours du Néolithique semble donc plutôt liée à d’éventuelles difficultés économiques qui accompagnent le changement climatique, l’arrivée de l’agriculture ou,
24 comme au Mésolithique, à l’occupation de certains biotopes ou de certaines aires géographiques particulières comme par exemple la vallée de la Seine avec le site de Noyen-sur-Seine ou la haute vallée de la Garonne avec les gisements de Buholoup (Montbéraud, Haute-Garonne) ou de la Balma Margineda (Aixovall, Andorre) (Le Gall 1991).
I.2.3 Stabilité de la pratique de la pêche ?
Ce troisième et dernier modèle est issu des résultats des travaux en biogéochimie isotopique réalisés depuis une quinzaine d’année sur les restes humains retrouvés dans les contextes archéologiques datés du Tardiglaciaire et de l’Holocène en France et en Europe (Danemark, Portugal, Suède, Angleterre, Ukraine, Belgique, Allemagne…). Ces études ont permis de reconstituer les anciens régimes alimentaires humains en se fondant sur l'analyse isotopique du collagène contenu dans les ossements archéologiques. Elles utilisent les teneurs en isotopes stables du carbone et de l’azote (13
C, 15N) dont l’analyse permet, par comparaison entre les valeurs obtenues pour certaines ressources alimentaires et les valeurs données par les ossements humains d'en déduire l’alimentation de ces derniers (Drucker, Henry-Gambier, Lenoir 2005, Dufour et al. 1999).
Dans ce modèle, la pratique de la pêche et la consommation du poisson par les populations humaines est attestée dès les périodes anciennes et se maintient tout au long de la séquence qui nous intéresse. Bien que relativement stable dans le temps, ce recours aux ressources aquatiques dans l’alimentation semble toutefois assez variable dans le détail, notamment en ce qui concerne les quantités de poissons consommées chez les populations étudiées (observations qualitatives).
En France, la consommation significative, voire majoritaire, de poisson est attestée dans le régime alimentaire humain dès le Paléolithique supérieur et final. Au cours du Magdalénien par exemple, les restes humains de l’abri Faustin en Gironde datés du Magdalénien supérieur final témoignent déjà d’une très grande dépendance aux ressources aquatiques au sein desquelles les Cyprinidés semblent jouer un rôle plus important que les Salmonidés (Drucker, Henry-Gambier, Lenoir 2005). Pour le Paléolithique final, la mandibule d’enfant datée de l’Azilien terminal à affinités laboriennes découverte dans la couche 2 du site de Pont d’Ambon en Dordogne témoigne également de la consommation de poisson (Cravinho 2009, Drucker, Bocherens 2001). Toutefois les individus analysés pour ces périodes ne consomment pas tous des ressources aquatiques comme l’illustre l’étude des sujets daté du Magdalénien indéterminé et du Magdalénien moyen des sites de Saint-Germain-la-Rivière et de Moulin-Neuf en Gironde (Drucker et al. 2005). Ainsi, bien que la consommation de protéines
25 d’origine aquatique soit avérée très tôt, il semble également exister une certaine variabilité dans le régime alimentaire des populations humaines au cours de ces périodes anciennes.
En ce qui concerne le Mésolithique, les données sont plus nombreuses et le même constat peut être fait. La consommation de protéines d’origine aquatique se poursuit mais les restes humains témoignent une fois de plus de la diversité des pratiques alimentaires. On distingue ainsi plusieurs tendances :
- La consommation d’une part significative de ressources aquatiques : les sujets de Hoëdic en Bretagne datés du Mésolithique récent par exemple témoignent d’une consommation massive de protéines d’origine marine avec comme ressource principale le poisson (Valentin, Drucker 2009). De même, les restes humains issus de la couche 9 datée de la fin du Mésolithique moyen du site du Haut-des-Nachères à Noyen-sur-Seine (Seine-et-Marne) témoignent d’une consommation significative de poissons d’eau douce. Dans ce site les informations vont même plus loin puisque la comparaison des données des ossements humains avec celles de restes de loutre issus du même contexte a permis de préciser la nature du poisson consommé. Les teneurs plus élevées en 13C et 15N de l’humain par rapport à la loutre pourraient indiquer la consommation de poissons de niveau trophique plus élevé (anguille, brochet) par l’être humain (Valentin, Drucker 2010). Ce constat va dans le sens des découvertes matérielles faites sur ce site où une nasse a été trouvée en association avec de nombreux restes de poissons en majorité attribués à l’anguille et au brochet, deux espèces de niveau trophique élevé (Dauphin 1987, Dauphin 1989).
- La consommation équilibrée de ressources d’origine aquatique et terrestre : les sujets du site de Téviec en Bretagne datés du Mésolithique récent par exemple semblent consommer des proportions assez similaires de protéines d’origine terrestre et aquatique (Valentin, Drucker 2009).
- La consommation d’une petite part de ressources aquatiques : les 9 individus mésolithiques de La Vergne en Charente-Maritime témoignent d’une consommation surtout tournée vers les protéines animales d’origine terrestre avec une contribution probable des protéines d’origine aquatique en faible quantité (Valentin, Drucker 2009).
- L’absence de consommation de ressources aquatiques : le sujet de Mareuil-lès-Meaux en Seine-et-Marne, attribué au Mésolithique moyen, présente un régime alimentaire essentiellement basé sur les protéines animales d’origine terrestre, en
26 particulier issues d’animaux vivant dans le milieu forestier dense (Valentin, Drucker 2010).
Au cours de ces études également, l’analyse approfondie de certaines données pour les sites de Téviec et Hoëdic a permis de préciser cette diversité des pratiques alimentaires au cours du Mésolithique (Schulting, Richards 2001). Dans ces deux sites funéraires qui ont livré 25 squelettes humains dont la chronologie s’étend sur une échelle de temps allant du Mésolithique ancien au Néolithique, la consommation des ressources d’origine marine est continue et attestée pour tous les individus. Dans le détail, les données de chaque sujet témoignent clairement de la complexité de l’alimentation humaine au cours de cette fenêtre de temps. Les critères d’âge et de sexe en particulier montrent quelques différences intéressantes : les femmes et notamment les jeunes femmes semblent moins consommer de poissons que les hommes, ce qui a permis d’émettre quelques hypothèses. La première pose la question de possibles interdits alimentaires auxquels auraient été soumises ces jeunes femmes, tandis que la deuxième s’intéresse aux mouvements de populations qui existaient à cette époque. Les jeunes femmes, en âge de se marier auraient ainsi pu venir s’installer à Hoëdic et Téviec depuis des sites où la consommation de protéines d’origine marine était beaucoup moins importante. Cette deuxième hypothèse est d’ailleurs appuyée par les données des femmes âgées dont la consommation de poisson rejoint celle des hommes (Schulting, Richards 2001). Ainsi, comme pour les périodes précédentes, si la consommation des ressources aquatiques est bien présente tout au long du Mésolithique, elle semble en revanche varier en fonction des groupes humains considérés. Notamment en fonction de l’environnement dans lequel étaient installés les sites, de l’accessibilité des ressources aquatiques voire en lien avec d’autres composantes sociales qu’il n’est pas toujours possible de cerner de manière précise.
Pour finir, le Néolithique ne fait pas exception et les travaux démontrent que la consommation des ressources aquatiques fait toujours partie du régime alimentaire humain. Toutefois, les populations néolithiques apparaissent souvent beaucoup moins dépendantes des ressources aquatiques que leurs prédécesseurs. Dans l’échantillon de Hoëdic et Téviec par exemple, l’individu le plus récent, daté du Néolithique consomme beaucoup moins de protéines d’origine marine que les sujets mésolithiques. Cette constatation trouve un écho dans les travaux réalisés au Portugal, en Suède, au Danemark, en Angleterre et en Irlande où une moindre dépendance aux protéines d’origine aquatique est également enregistrée. Ce passage vers un régime alimentaire plus « terrestre » n’est cependant pas brutal et semble s’opérer progressivement entre 7000 et 6000 cal BC dans le sud de l’Europe et entre 5500 et 3300 cal BC dans le nord de l’Europe où l’arrivée de l’agriculture est plus tardive, les prémices de ce nouveau régime étant d’ailleurs parfois visible dès la fin du Mésolithique (Bonsall et al. 2009, Dufour, Bocherens, Mariotti 1999, Lubell et al. 1994, Quill Smart 2003, Richards et al. 2003).
27 Toutefois, la majorité des échantillons analysés appartiennent à des individus issus de populations côtières ce qui peut représenter un biais. Pour le Néolithique il est donc nécessaire d’approfondir le travail pour les populations continentales avant de pousser plus avant ces constatations. D’autres exemples européens appuient d’ailleurs ces mises en garde puisque l’alimentation de leurs populations témoigne à l’inverse d’un concours encore important des protéines d’origine aquatique : en Belgique dans le bassin de la Meuse (Bocherens et al. 2007, Schulting, Richards 2001) et dans la région du Danube, du Dniepr, et de la mer Noire (Lillie, Richards 2000, Richards, Price, Koch 2003).
Pour en conclure avec ce dernier modèle, malgré les nombreux échantillons qui témoignent de la complexité du régime alimentaire humain au cours de ces périodes, il semble clairement exister une continuité dans la pratique de la pêche et de la consommation du poisson depuis les périodes anciennes jusqu’au Néolithique. Ces tendances semblent généralisées en France et en Europe où les témoignages concordent. Au cœur de ce modèle, ce sont plutôt les modalités des variations enregistrées au sein du régime alimentaire humain qui restent le plus souvent difficiles à cerner. Toutefois, la multiplication des études au cours de ces dernières années laisse entrevoir des résultats intéressants qui permettront sans doute de répondre à ces interrogations.
I.2.4 Synthèse : évolution ou stabilité de la pratique de la pêche au cours de la transition Tardiglaciaire-Holocène ?
Les trois modèles présentés ci-dessus démontrent la complexité de l’approche d’un sujet aussi précis que la question de l’évolution de la pratique de la pêche et de la consommation du poisson par les populations humaines au cours de la transition Tardiglaciaire-Holocène. En effet, au sein des trois hypothèses de recherches actuelles, les tendances apparaissent très différentes voire complètement opposées (figure 2 ci-dessous). S’ajoute à ces constatations la perception du statut de la pêche par les différents auteurs qui diffère également d’un modèle à l’autre. Dans les deux premiers modèles, la pêche est perçue comme une activité secondaire, de soutien alimentaire en période difficile. Dans le troisième modèle en revanche, les auteurs semblent plutôt attribuer la pratique de la pêche et la consommation du poisson à des raisons pragmatiques (sites côtiers) ou sociales. Outre la question de l’évolution de la pratique de la pêche au cours du temps, la question du statut de cette activité sera donc également intégrée à la problématique de recherche de ce travail.
Afin d’y répondre, il est important de constater d’ores et déjà que les informations relatives à cette activité semblent difficiles à obtenir et peuvent conduire à des conclusions diverses. En effet, l’information semble intégrée à différentes sources mais de manière partielle, chacune ne permettant alors de saisir qu’une partie du phénomène. De plus, la précision de ces
28 données est extrêmement variable, allant de simples observations et hypothèses de recherche à des analyses statistiques ou scientifiques poussées, complexifiant encore le problème notamment en ce qui concerne la fiabilité et les possibilités de comparaison des résultats obtenus.
Toutefois, au-delà de cet aspect complexe voire problématique de l’état de la recherche, je pense qu’il faut plutôt voir ici des aspects complémentaires, qui démontrent l’intérêt d’aborder cette activité et l’ensemble des données qui lui sont associées de manière globale. Cependant, reconstituer les modalités d’évolution de la pratique de la pêche de manière globale ne sous-entend pas d’utiliser l’ensemble des informations disponibles sans envisager un examen préalable détaillé de celles-ci. Ainsi pour répondre de la manière la plus fiable et la plus détaillée possible à la question assez vaste que se propose d’étudier ce travail, il est indispensable d’effectuer une première phase d’analyse sur les sources elles-mêmes, ceci afin de sélectionner les témoignages qui seront les plus à même d’apporter une base fiable et une réponse la plus complète possible à la problématique posée. Ainsi nous poursuivrons ce chapitre dans cette optique en explorant de manière critique les sources matérielles, les échelles de temps et d’espace mais également les méthodes les plus à même de fournir des informations de qualité.
29 Figure 2 : Schéma synthétique présentant les hypothèses d’évolution de la pratique de la pêche au cours de la transition Tardiglaciaire-Holocène.