Global mismatch between fishing dependency and larval supply from marine reserves
Texte intégral
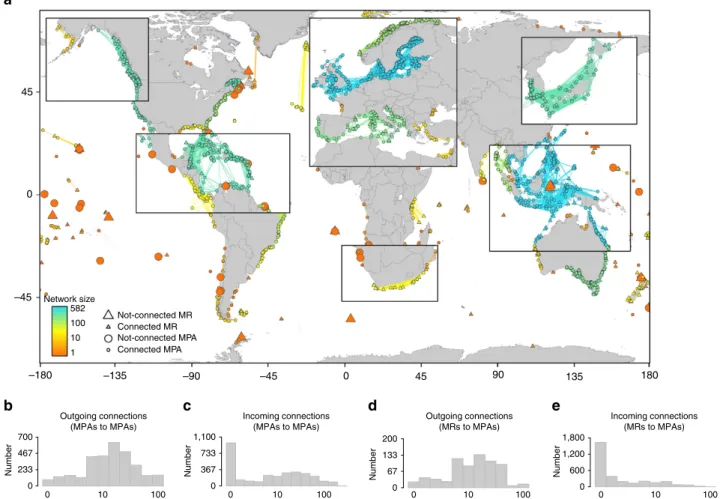
Figure



Documents relatifs
The New Caledonian archipelago offers a unique opportunity to explore these relationships since (i) human population density is very heterogeneous along a marked gradient from
In this section, as a supplement to the main theorems stated in Sec- tion 1, we give a yet another result of the failure of stable rationality of smooth weighted hypersurfaces,
Therefore, this will not affect its price, nor the number of issues sent to subscribers, nor the publication of a second issue next autumn.. There are two reasons
US-Canada Pacific Salmon Treaty signed in 2008 resulting in 50% loss of fish for west coast Vancouver Island trollers. Comprehensive plan by AMB to deal with treaty impacts
In the context of larval dispersal and population connectivity, the two strategies consisting in (1) laying few, large, demersal eggs, giving birth to larvae which develop quickly
A means to prove this conjecture is to remark that the APN property is equivalent to the fact that the rational points of a certain algebraic surface X in a 3-dimensional space
[6], fishing is allowed in the so called artisanal zone (corresponding to the protected area in our case) and the objective to be maximised is somewhat different (it takes into
We consider priced timed games with one clock and arbitrary (positive and negative) weights and show that, for an important subclass of theirs (the so-called simple priced timed
