Oxygen deprivation stress in a changing environment
15
0
0
Texte intégral
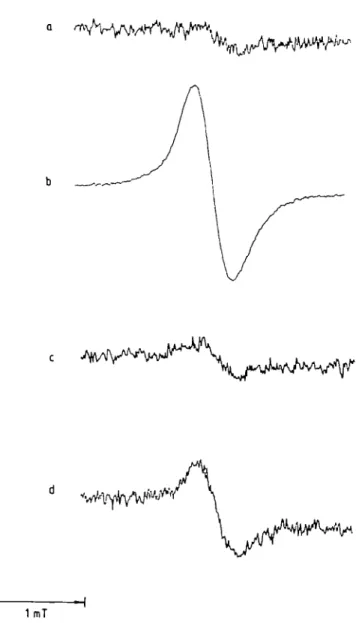
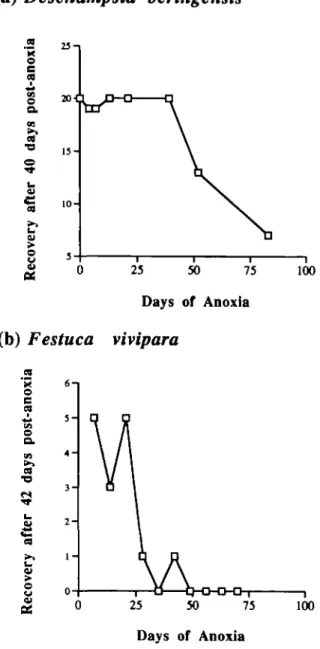
Figure



+4


Documents relatifs