HAL Id: hal-02823535
https://hal.inrae.fr/hal-02823535
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Christian Cocquempot, Ake Lindelöw
To cite this version:
Christian Cocquempot, Ake Lindelöw. Longhorn beetles (Coleoptera, Cerambycidae). Alien terrestrial arthropods of Europe, 4 (1), Pensoft Publishers, 2010, BioRisk, 978-954-642-554-6. �10.3897/biorisk.4.56�. �hal-02823535�
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 193
Longhorn beetles
(Coleoptera, Cerambycidae)
Chapter 8.1
Christian Cocquempot1, Åke Lindelöw2
1 INRA UMR Centre de Biologie et de Gestion des Populations, CBGP, (INRA/IRD/CIRAD/Montpellier
SupAgro), Campus international de Baillarguet, CS 30016, 34988 Montférrier-sur-Lez, France 2 Swedish university of agricultural sciences, Department of ecology. P.O. Box 7044, S-750 07 Uppsala, Sweden
Corresponding authors: Christian Cocquempot (cocquemp@supagro.inra.fr), Åke Lindelöw (
Ake.Linde-low@ekol.slu.se)
Academic editor: David Roy | Received 28 December 2009 | Accepted 21 May 2010 | Published 6 July 2010
Citation: Cocquempot C, Lindelöw Å (2010) Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1. In: Roques A et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4(1): 193–218. doi: 10.3897/biorisk.4.56
Abstract
A total of 19 alien longhorn beetle species have established in Europe where they presently account for ca. 2.8 % of the total cerambycid fauna. Most species belong to the subfamilies Cerambycinae and Laminae which are prevalent in the native fauna as well. Th e alien species mainly established during the period 1975–1999, arriving predominantly from Asia. France, Spain and Italy are by far the most invaded countries. All species have been introduced accidentally. Wood-derived products such as wood- packaging material and palettes, plants for planting, and bonsais constitute invasive pathways of increasing impor-tance. However, only few species have yet colonized natural habitats outside parks and gardens. Present ecological and economical impacts, and future trends are discussed.
Keywords
Cerambycidae, Europe, Introductions, Establishments, Biogeographical origins, Pathways, Impacts
8.1.1 Introduction
Th e coleopteran family Cerambycidae (longhorn beetles) is currently classifi ed in the superfamily Chrysomeloidea, along with the families Vesperidae and Distenidae (Hunt et al. 2007, Szeoke and Hegyi 2002). Cerambycidae is a large family comprising about BioRisk 4(1): 193–218 (2010)
doi: 10.3897/biorisk.4.56 www.pensoftonline.net/biorisk
Copyright C. Cocquempot, Å. Lindelöw. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
RESEARCH ARTICLE
BioRisk
40000 described species worldwide. Longhorn beetles are all phytophagous. Larvae may be found in conifer, deciduous and fruit trees, in bushes and herbaceous plants. Th ey are mainly xylophagous borers of living, decaying or dead wood. Some species also bore small twigs, roots or fruit endocarps. Th ey usually have a long period of lar-val development, some species being capable of developing in woody material a long time after the death of the tree. Th ey are thus very susceptible to transport with wood products, facilitating their introduction and establishment.
Th e oldest known introduction of a longhorn beetle from one continent to anoth-er was probably that of the house boranoth-er, Hylotrupes bajulus (L., 1758), which was fi rst described by Linnaeus from both Europe and ‘America septentrionali’ (von Linnaeus 1758). Since a study by Duff y in 1953 (Duff y 1953a) for Great Britain, there has been no further large synthesis of the alien cerambycid species introduced to Europe. Since 1999, the development of research interests in the Asian longhorn beetles, Anoplophora spp., in North America has raised awareness of the risks presented by cerambycid im-portation and provided a baseline for subsequent studies (Haack et al. 2000, Haack et al. 2010). Th ere is an urgent need for a comprehensive literature review of the alien cerambycids that have successfully established in Europe.
Th e exponential growth in the volume of international trade in both horticulture and forestry has allowed an increasing number of wood products and ornamental plants potentially containing cerambycids to arrive in Europe. More than 250 species have been introduced to Europe or moved within Europe since the middle of the 18th cen-tury (Cocquempot 2007) but most of them never established. We have identifi ed 19 species alien to Europe that have established in Europe but have not yet been eradicated.
8.1.2 Taxonomy of the Cerambycid species alien to Europe
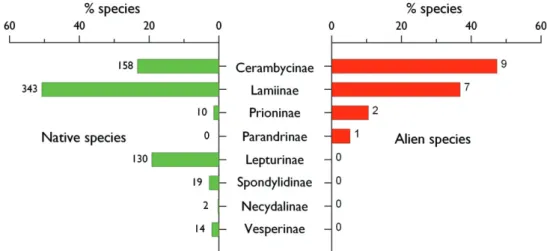
Taxonomy in Cerambycidae sensu lato is not well established (e.g., Hunt et al. 2007, Lawrence and Newton 1995, Napp 1994, Özdikmen 2008, Sýkorová 2008) but a gen-eral consensus exists about the presence in Europe of 7 subfamilies, namely Ceramby-cinae, Lamiinae, Lepturinae, Necydalinae, Prioninae, Spondylidinae, and Vesperinae (the latter being sometimes considered as a valid family). A total of 677 native species are known to occur in Europe (Althoff and Danilevsky 1997, Fauna Europaea), being largely dominated by 3 subfamilies (Lamiinae- 343 spp.; Cerambycinae- 158 spp.; Lepturinae- 130 spp.) which account for 93.2% of the total.
Th e 19 alien species established in Europe belong to only 3 of these subfamilies, Cerambycinae, Laminae and Prioninae (Table 8.1.1). Th e alien species are mostly rep-resented by the subfamily Cerambycinae, followed by Lamiinae but the relative propor-tion of aliens compared to the total cerambycid fauna is still limited (<6%) in these two subfamilies. By contrast, the proportion of aliens is much more important in Prioninae with 2 species adding to 10 native ones (Fig. 8.1.1.). In addition, Parandrinae, a sub-family which is not represented in the native European entomofauna, is represented by Parandra brunnea, a North American species introduced in Germany (Nüssler 1961).
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 195
Two more alien species have been introduced and established in Israel, Batocera
rufomaculata (DeGeer, 1775) (Bytinski-Salz 1956, Chikatunov et al. 1999, Sama et
al. 2010) and Xystrocera globosa (Olivier, 1795) (Chikatunov et al. 2006, Sama et al. 2010), but they have not yet spread to Europe and were not considered in Table 8.1.1.
Table 8.1.2 gives a list of species of European origin introduced through human activity in another part of Europe (aliens in Europe). Th ese species are mostly of Medi-terranean origin introduced in more northern areas and species from Continental Eu-rope introduced to the Atlantic islands.
8.1.3 Major biological characteristics of the cerambycid species alien to Europe
Lepturinae but also Prioninae and Parandrinae share some biological characteristics that reduce their probability of introduction. Larvae in these subfamilies develop in decaying wood and are rarely imported with wood products or living plants. Intercep-tions have shown that they are mainly introduced through accidental importation in industrial packages or in stocks of perishable vegetables. Only a few species of Lepturi-nae (Tribe Rhagiini, and some Lepturinii) developing on recently felled trees are likely to be successfully introduced through the wood trade. Th e importation of living potted plants is also a potential new pathway for Prioninae.
Cerambycinae and Lamiinae seem more predisposed to introduction. Most species develope in living plants and several Cerambycinae undertake their entire life-cycle in dead wood, e.g. the cosmopolitan tribe Hesperophanini and the species Hylotrupes
bajulus and Gracilia minuta. Th us, Cerambycinae and Lamiinae can easily survive Figure 8.1.1. Relative importance of the subfamilies of Cerambycidae in the alien and native entomo-fauna in Europe. Subfamilies are presented in a decreasing order based on the number of alien species. Species alien to Europe include cryptogenic species. Th e number over each bar indicates the number of species observed per family.
throughout the importation process of living plants including bonsai (e.g.
Anoplo-phora chinensis (Cocquempot 2007, EPPO 2006, van Rossem et al. 1981, Schmidt
and Schmidt 1990)), recently felled logs and other non-aged wood products (e.g.
Ano-plophora glabripennis (Cocquempot et al. 2003, Haack et al. 2000), Monochamus spp.
(Cocquempot 2007, Cocquempot (Unpubl.), Duff y 1953a), Chlorophorus annularis (Cocquempot 2007) and Phoracantha spp. (Cocquempot and Debreuil 2006)). Spe-cies in the genera Hesperophanes, Trichoferus, and Stromatium can emerge from wood products even several years after importation (Duff y 1953a).
Once a population is introduced, the capability for natural dispersal constitutes an important factor for establishment success. Although our knowledge about the disper-sal behaviour of alien longhorn beetles is still rather limited and mostly concerns only a few species of recent invaders such as Anoplophora glabripennis (Smith et aol. 2001) and A. chinensis (Adachi 1990, Komazaki and Sakagami 1989), this variable is impor-tant when designing an eradication attempt (MacLeod et al. 2002).
8.1.4 Temporal trends of introduction in Europe of alien Cerambycids
Figure 8.1.2 presents the temporal changes in the records of Cerambycid species alien
to Europe from 1492 to 2007. Cerambycids have tracked trade routes since the
begin-ning of overseas communications. Th e fi rst species to have moved are those which live in dry wood and undergo a long stage of larval development. Th ese species have be-come cosmopolitan (e.g. Hylotrupes bajulus) or nearly so (e.g. Stromatium spp.). With the increased speed of international transport from 1850 to 1925, species with shorter life cycles were able to reach Europe alive and become established, e.g. Neoclytus
acu-minatus (Reineck 1919, Sama 2002, Tassi 1969). Later, only two species were
intro-duced from North America to Europe via the US eff ort to supply extra furniture and increase military material after the 1st World War (i.e., Parandra brunnea, Neoclytus
acuminatus). Subsequently, 50 years passed until a second wave of introduction
ar-rived alongside with the rapid development of international exchange of goods and transportation after the 2nd World War. During the recent period, two further species have been detected in the wild - Anoplophora chinensis in 2000 in Italy (Colombo and Limonta 2001) and A. glabripennis in 2001 in Austria (Dauber and Mitter 2001).
Th e number of interceptions of Cerambycids is still increasing throughout Eu-rope. However, more eff ective control at borders is like to have reduced establishments following interception or introductions. Th e importation of exotic plants also off ers opportunities for introduction but also constraints the establishment of some alien species. For example, Phoracantha spp. could not have been introduced without the importation and mass cultivation of its host plants, Eucalyptus spp. in the Mediter-ranean basin. In south-eastern France, an Australian cerambycid, Bardistus cibarius (Newman, 1841) could survive only on its original host plant, an introduced grass tree (Xanthorrhoea sp., Xanthorrhoeaceae); the beetle population disappeared immediately after the infested host plants were removed (Cocquempot 2007). Th e case of Batocera
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 197
rufomaculata (DeGeer, 1775) found in Munster’s Zoo (Germany) is similar
(Coc-quempot 2007) although this tropical species has established in Israel since at least 1948 (Bahillo de la Puebla and Iturrondobeitia-Bilbao 1995, Plavilstshtikov 1934, Sama et al. 2010). Th e combination of importation of longhorn beetle species with their specifi c host plant or groups of plants followed by establishment is rare. However the establishment of A. chinensis is an exception. Other species are frequent intercepted at border controls, e.g. Mimectatina meridiana (Matsushita, 1933) with Cycas fruits from Japan (Cocquempot 2007) or Trichoferus campestris (Faldermann 1835) with
Sa-lix timber from China (Cocquempot 2007).
Th e degree of polyphagy is also an important factor in the likelihood of establish-ment. Polyphagous species appear to have a higher potential to establish than oligopha-gous and monophaoligopha-gous species. Th e large number of hosts utilised by Anoplophora spp. (Cocquempot et al. 2003, Hérard and Roques 2009, Maspero et al. 2007a) is a main factor in the diffi culty in eradicating this species for example. Th ese diffi culties appear much less important for oligophagous species such as Callidiellum rufi penne (Bahillo and Iturrondobeitia-Bilbao 1995, Campadelli and Sama 1988, Plavilstshtikov 1934) Figure 8.1.2. Temporal changes in the mean number of new records per year of Cerambycid species alien to Europe from 1492 to 2007. Th e number over each bar indicates the absolute number of species newly recorded per time period.
or Phoracanthine species. It is also the case for the North American wood borer Saperda
candida (Fabricius, 1787), which was introduced in Germany in 2008 but apparently
did not established yet (EPPO 2008, Nolte Krieger 2008). By contrast, Monochamus species have a regime close to polyphagy, including a large number of conifer species, and may spread throughout Europe. Th ere is no example of establishment in Europe of a strictly monophagous exotic long-horned beetle. Species with a limited host range do not seem to be capable of going beyond the interception or introduction stage, e.g.
Bardistus cibarius (Cocquempot 2007).
8.1.5 Biogeographic patterns of the cerambycid species alien to Europe
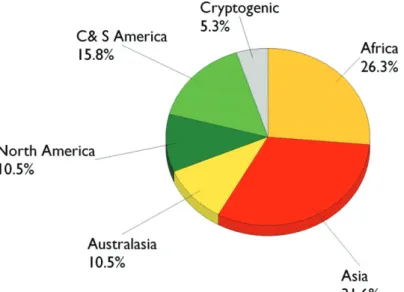
Alien species established in Europe mostly originated from Asia, followed by Africa (Figure 8.1.3). Th e region of origin appears to depend on the major trade routes de-veloped by each country. Some North African species have colonized Mediterranean countries such as Spain, France, and Malta for example. Other African species have often been intercepted but only Phryneta leprosa has established in Malta where the climate is favourable for development (Mifsud and Dandria 2002). Long-established trade routes between Iberian countries and South American countries have resulted in some historic, isolated establishments in the Spanish and Portuguese Atlantic Is-lands but with a limited risk of further expansion (Lemos-Perreira 1978, Méquignon 1935). With the numerous interceptions in the U.K (Duff y 1953a) together with the colonial trade routes with African and Asiatic countries, it is surprising that only
Tri-nophylum cribratum has established to date (Gilmour 1948); the incompatible climate
may negate the development of tropical and subtropical species. Two species native to North America, Parandra brunnea and Neoclytus acuminatus, also colonized Europe at the beginning of the last century. Th e fi rst species is well established but restricted to Dresden (Germany) (Nüssler 1961). Th e second is widely established in the Mediter-ranean area but its populations appear to be declining (Brustel et al. 2002). Beside these two species, there have been no further establishments originating from North America; the pathway of transported material is mainly in the reverse direction, from Europe to America.
Some Australian species have reached Europe but only those using Eucalyptus
(Pho-racantha spp.) have successfully established (Cocquempot and Sama 2004) and only in
areas newly planted with these fast-growing tree species. Th e large diff erences in spe-cies composition between the fl oras of Australia and Europe probably accounts for the failure of Australasian longhorn beetles such as in Bardistus cibarius on Xanthorrhoea sp. (Cocquempot 2007) to establish.
Recent increases in commercial traffi c from Asia (especially China) to Europe has accounted for the introduction of a number of new species of cerambycids. Striking examples are Callidiellum rufi penne which has recently established in Spain (Bahillo de la Puebla and Iturrondobeitia-Bilbao 1995) and Italy (Campadelli and Sama 1988),
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 199
not eradicated in several countries (Hérard and Roques 2009, Maspero et al. 2007a),
Psacothea hilaris (Pascoe, 1857) under eradication in Italy (Cocquempot 2007, Jucker
et al. 2006), and Monochamus alternatus Hope, 1842 intercepted a number of times in Germany (Cocquempot 2007) and France (Cocquempot Unpubl.) but not yet estab-lished. A fi nal case, Xylotrechus stebbingi, is less clear. It is believed that an initial intro-duction from its native area of central Asia to Asia Minor was followed by a step-wise expansion into southern Europe and North Africa (Cocquempot and Debreuil 2006, Sama 2002, Šefrová and Laštůvka 2005).
Alien cerambycid species are not evenly distributed throughout Europe. Large dif-ferences in the number of aliens are apparent between countries, France, Italy and Spain being by far the most invaded (Figure 8.1.4).
8.1.6 Main pathways of introduction to Europe of alien cerambycid spe-cies
All alien longhorn beetles established in Europe have been introduced accidentally; there are no examples of a successful, deliberate introduction. Th e principal pathways of arrival have been identifi ed and presented by Frank 2002 and each relates to the import of immature stages that subsequently emerge as adults. Th ere are relatively few records of living adults imported with vegetables or fruits although Eucalyptus beetles,
Phoracantha recurva, were found in a cluster of bananas (Bosmans 2006).
Th e longest established pathway is timber importation for house construction
(Hy-lotrupes bajulus) or building furniture (e.g. Trichoferus spp., Stromatium spp. and Chlo-rophorus annularis arriving with bamboo- made objects (Cocquempot 2007)). Species
introduced through this pathway have traditionally required a long life cycle but more rapid travel now enables the introduction of species with a one year life cycle. Th e second pathway is via the importation of timber for pulp (e.g., for Phoracantha spp.). A third, more recent, pathway concerns wood packages, palettes and other wood-de-rived products (e.g., for Anoplophora glabripennis) (Hérard and Roques 2009). Th e fi nal pathway is the importation of plants for planting in nurseries, including the bon-sai industry, which has resulted in the arrival of species such as Anopolophra chinensis (Cocquempot 2007, EPPO 2006, van Rossem et al. 1981, Schembri and Sama 1986),
Callidiellum rufi penne and Bardistus cibarius.
All pathways are still prevalent but they vary in importance. Most recent inter-ceptions (from the end of the 20th Century) have related to wood-manufactured products (e.g. Chlorophorus annularis and Trichoferus campestris). Importation of
Eu-calyptus wood for pulp has also resulted in the introduction of a second species of Phoracantha, P. recurva (Miquel 2008). If such importations continues a number of
Figure 8.1.4. Comparative colonization of continental European countries and islands by Cerambycidae species alien to Europe. Archipelago: 1 Azores 2 Madeira 3 Canary islands.
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 201
additional species of this genus, which are mainly related to Eucalyptus(Wang 1995), are expected to arrive.
Since their fi rst usage, wood packaging and palettes have constituted an impor-tant introduction pathway. Th e source material spends suffi cient time as logs without sanitary controls to be colonized by longhorn beetles. When the wood is turned into packages or palettes, infestation occurs mainly as unnoticed early stages (eggs or fi rst- instar larva). Development continues in the woody material during importation and emergence of adults occurs often unnoticed in warehouses, weeks or months after ar-rival. Th is is the case for A. glabripennis, P. hilaris and M. alternatus which may already complete their entire lifecycle before the source wood is processed or destroyed. Wood package is often produced using low quality timber often colonized by longhorn beetle species, which is increasing its potential as a vector.
Other, less signifi cant, introduction pathways have also been identifi ed, yet they typically only transported one or a few individuals which fail to establish. Th e intro-duction route is unknown for other species such as Acanthoderes jaspideus (Méqui-gnon 1935), Oxymerus aculeatus (Alluaud 1935), Deroplia albida, and Phryneta leprosa (Mifsud and Dandria 2002) but they may be related to the uncontrolled importation of wild plants. Natural range expansion cannot be ruled out for a few species which have a nearby native range, e.g. Lucasianus levaillantii (Mayet 1905, Pellegrin and Cocquempot 2001) and Xylotrechus stebbingi (Šefrová and Laštůvka 2005) originating from North Africa and the Middle East, respectively.
8.1.7 Ecosystems and habitats invaded in Europe by alien cerambycid species
Although all natural or artifi cial terrestrial ecosystems and anthropogenic areas which contain trees, bushes or wood products are potentially occupied by alien longhorn bee-tles, establishment in Europe is concentrated in man-made habitats to date, especially in parks and gardens (Figure 8.1.5). To date, only the two clytine beetles, Neoclytus
acumi-natus and Xylotrechus stebbingi, have colonized natural habitats. X. stebbingi is very
com-mon on Eucalyptus cut wood in Crete (Sama 2002) for example and may be related to the polyphagous nature of these two species. Other polyphagous species such as Anoplophora spp. also have the potential to live in urban areas, in cultivated lanes (e.g. planted with poplars) as well as in natural forests where potential host plants occur. However, disper-sal from man-made habitats to natural forests appears to be a slow process. For the fi rst twenty-two years since its arrival in North America, A. glabripennis has been restricted to trees in urban areas until 2008 when it was found in natural forests dominated by Acer trees (Haack et al. 2010). Although such a process has not yet been observed in Europe, there is a strong risk that Anoplophora spp. will spread to naturally-forested landscapes, if the ongoing eradication attempts in Austria, Germany, France and Italy are unsuccessful.
Th e expansion of oligophagous species is inevitably more dependant on the pres-ence of suitable host plants. Th ose using largely- planted trees can spread more easily.
Th us, Phoracantha spp. that live only in eucalypt trees have colonized ornamental tree plantations in urban areas as well as old plantations such as those found on the Medi-terranean islands and in neighbouring countries, and industrial plantations created for paper pulp. Other established species mostly have a distribution restricted to Mediter-ranean and Atlantic islands. In these areas, anthropogenic ecosystems are mainly colo-nized. A species of considerable concern with conifer forests is Monochamus alternatus, which could potentially become established in coniferous plantations and forests and subsequently transfer the pine wood nematode (Bursaphelenchus xylophilus Steiner & Buhrer, 1934).
8.1.8 Ecological and economic impact of alien cerambycid species
Although there is concern about the potential ecological impact of the invasive long-horn beetles N. acuminatus and X. stebbingi, there is no measure of their impact on trees or any estimation of possible competitive displacement of the native fauna. Th e ecological impact of Anoplophora species may also be important if they establish in European forests. Anoplophora could compete with other arthropods occupying the same niche, but they also create niches for other arthropods that live in tunnels in decaying wood or compete with other saproxylic beetles. Th e joint introduction Figure 8.1.5. Main European habitats colonized by the established alien longhorn beetles. Th e number over each bar indicates the absolute number of alien longhorn beetles recorded per habitat. Note that a species may have colonized several habitats.
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 203
and establishment of the Citrus longhorn beetle, A. chinensis, and its parasitoid,
Aprostocetus anoplophorae Delvare, 2004, exemplifi es the potential risk of adaptation
of imported parasitoids which themselves might not specialise on the native fauna (Delvare et al. 2004).
Although the ecological niche occupied by an alien species may be vacant there remains a risk of secondary infection resulting from their damage. For example, sec-ondary infestation by the pine wood nematode vectored by Monochamus spp. (Evans et al. 2008, Kawai Miho et al. 2006) may cause serious impacts to coniferous trees in all landscapes. M. alternatus has only been intercepted in Germany and France (Cocquempot 2007, Cocquempot (Unpubl.)); yet the pine wood nematode which it vectors was recorded from Portugal in 1999 (Mota et al. 1999). After having been contained for several years in a limited area, the nematode has spread throughout Por-tugal, as well as being eradicated following incursions into Spain in 2008 and Madeira in 2009. A novel association with the native species, M. galloprovincialis (Villiers 1967) has also been reported. Th e expansion as well as new introductions of the pine wood nematode could potentially have a substantial level of economic impact in all areas of coniferous cultivation in Europe.
Other economic impacts are mainly associated with ornamental trees in urban areas, cultivated trees such as poplars and eucalypts and nurseries, including these for bonsai production. Studies of Anoplophora glabripennis in North America and A.
chinensis in China indicate the possible scale of economic damage following
estab-lishment of these species in a new country or in a plantation, especially of poplar or Citrus trees (Cocquempot et al. 2003, Haack et al. 2010, MacLeod et al. 2002). As a control measure, ornamental trees colonized by invasive longhorns must be elimi-nated without consideration of their aesthetic value. Eradication measures entail high costs to be borne by local communities or private owners. Special attention is paid to A. chinensis necessitating complete removal of trees, including the rootstock (Haack et al. 2010).
Poplars or eucalypt plantations can be highly aff ected as has already been the case in China (A. glabripennis on poplars) and in Spain (Phoracantha spp.), where infested trees become unsuitable for pulp and wood exploitation. Th e Citrus longhorn beetle is also considered as an important risk for all Citrus fruit production in the Mediter-ranean area and its islands.
Th e nursery industry is already concerned. Th ere are several examples of introduc-tions or establishments of potentially invasive species such as Callidiellum rufi penne and Anoplophora chinensis, with the imports of nursery plants. Nurseries can them-selves be vectors of aliens when they dispatch their products.
Th e eradication process established for quarantine species aims to limit introduc-tions although only a few eradicaintroduc-tions have been offi cially reported in Europe, e.g. as for Anoplophora chinensis in France (Hérard et al. 2006, Hérard and Roques 2009). Phytosanitary interceptions at borders are likely to have prevented a number of intro-ductions and further establishments (e.g., Monochamus alternatus, Trichoferus
(Cocquem-Figure 8.1.6. Adults of some alien longhorn beetle species. a Phoracantha semipunctata b Phoracantha recurva c Mimectatina meridiana (Credit: Christian Cocquempot) d Xylotrechus stebbingi (Credit: Vítěslav Maňák) e Bardistes cibarius (Credit: Christian Cocquempot) f Psacothea hilaris g Parandra brunnea (a, b, e, f, g: Credit: Henri-Pierre Aberlenc).
pot 2007) whilst at the same time, several non-quarantine species not submitted to importation controls have become established (e.g., Xylotrechus stebbingi, Phoracantha
semipunctata, Neoclytus acuminatus). Th is illustrates the importance of quarantine spe-cies lists, which should be preventive and not only curative to be most eff ective.
Human-mediated dispersal should also be tightly controlled during the eradica-tion process. Without due respect for control obligaeradica-tions, eradicaeradica-tion can fail. For example, the long delay by Italian authorities in applying control measures and strong management measures against Anoplophora chinensis (EPPO 2009, Jucker et al. 2007) or inadvertent movement of untreated wood material for A. glabripennis in New-York (Haack et al. 1997) are examples of ineff ective eradication effi cacy.
a b f
e g
c
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 205
8.1.9 Expected trends
Th e combination of increasing volumes of trade, the increased speed of import of potential vectors, the diversity of sources and sites for introduction is likely to result in increasing invasion risk (Cocquempot 2007). All recently established species alien
to Europe have been intercepted too late after their introduction and have been
out-side offi cial institutional controls. Th ese factors make it increasingly diffi cult for rapid eradication after initial arrival. Eff ective monitoring of each point of possible entry is unfeasible when the key pathways identifi ed here have diff erent vectors and locations of arrival (e.g. airports, harbours, stations, lorry parks), and there are major diff er-ence in the quality of phytosanitary controls between European countries, particularly following the enlargement of the EU. Th e risk depends on volume and diversity of vector material imported, and subsequently there is greatest risk in countries such as the UK, France, Spain, Italy, Netherlands, Belgium and Germany. Th e case of
Anopolo-phora glabripennis in North America and Europe clearly demonstrates the possibility of
spread in our continent; such detailed assessment is required for all potentially invasive longhorn beetles (MacLeod et al. 2002).
According to Worner (2002), progress in the knowledge of invasion processes and associated preventive measures have not been followed by actions since the late 1980’s. Preventive methods are still routinely applied, e.g. the application of ISPM 15 (Interna-tional Standard for Phytosanitary Measures No.15), which set standards for heat treat-ment and fumigation of wood product materials used in international trade is likely to limit the arrival of longhorn beetles related to these materials although a few have been found to survive (Haack et al. 2010). However, this method is not uniformly applied to all imported living trees, shrubs plants for planting or bonsais. Th us, a high number of imported bonsais or other nursery trees infested with Anoplophora chinensis are still discovered (Hérard and Roques 2009). Although importation controls could be im-proved, they will never off er full protection. Further, controls which reduce the risk of introduction are mainly restricted to quarantine species. Post-interception or controls at importation points should be extended to all the potential pests posing risk and not be restricted to quarantine species already intercepted, introduced or established.
References
Adachi I (1990) Population studies of Anoplophora malasiaca adults (Coleoptera: Cerambyci-dae) in a Citrus grove. Research Population Ecology 32: 15–32.
Adlbauer K (2006) personnal communication.
Allenspach V (1973) Insecta Helvetica 3 – Coleoptera Cerambycidae. Züric, Switzerland: Fotoro-tar AG. 216 pp.
Alluaud C (1935) Les Coléoptères des îles Salvages. Revue française d’Entomologie 2: 35–44. Althoff J, Danilevsky ML (1997) A Check-List of Longicorn Beetles (Coleoptera, Cerambycoidea)
Bahillo P, Iturrondobeitia-Bilbao JC (1995) Primera cita de Callidiellum rufi penne (Motschuls-ky, 1860) para la Península Ibérica (Coleoptera: Cerambycidae). Boletín de la Asociación
española de Entomología 19: 204.
Bercedo P, Bahillo P (1998) Phoracantha recurva (Coleoptera: Cerambycidae): Una nueva plaga en los eucaliptales españoles. Boletín de la Sociedad Entomológica Aragonesa 23: 52. Bercedo P, Bahillo P (1999) Primera cita para Europa de Phoracantha recurva Newman, 1840
(Coleoptera, Cerambycidae). Estudios del Museo de Ciencias Naturales de Alava 14: 169– 174.
Berger P (1992) Présence de Phoracantha semipunctata F. dans le sud de la France : une menace pour les eucalyptus (Coleoptera Cerambycidae). Bulletin mensuel de la Société Linnéenne de
Lyon 61: 301–304.
Bertolini S (1872) Catalogo sinonimico e topografi co dei Coleotteri d’Italia. Firenze : Tipografi a Cenniniana, 263 pp.
Bijaoui R (1980) Notes de chasses et observations diverses : Neoclytus acuminatus (Col. Ceram-bycidae) dans le Tarn. L’Entomologiste 36: 268.
Borges PAV, Cunha R, Gabriel R, Frias Martins A, Silva L and Vieira V (eds) A list of the
terres-trial fauna (Mollusca and Arthropoda) and fl ora (bryophyta, Pteridophyta and Spermatophyta) from the Azores. Direccao Regional do Ambiente and Universidade das Açores, 317 pp.
Bosmans B (2006) Phoracantha recurva (Coleoptera: Cerambycidae) found in a cluster of ba-nanas Phegea 34: 105.
Brustel H, Berger P, Cocquempot C (2002) Catalogue des Vesperidae et des Cerambycidae de la faune de France (Coleoptera). Annales de la Société entomologique de France 38: 443–461. Bytinski-Salz H (1956) Th e Cerambycidae of Israel. Bulletin of the Research Council of Israel
5B: 207–226.
Cadahia D (1980) Proximidad de dos nuevos enemigos de los Eucalyptus en España. Boletín
del Servicio de Plagas forestales 6: 165–192.
Campadelli G, Sama G (1988) Prima segnalazione per l’Italia di cerambicide giapponese:
Cal-lidiellum rufi penne Motchulsky. Bollettino dell’Istituto di Entomologia “Guido Grandi” della Universita degli Studi di Bologna 43: 69–73.
Carter M, Smith M, Harrison R (2009) Genetic analyses of the Asian longhorned beetle (Co-leoptera, Cerambycidae, Anoplophora glabripennis), in North America, Europe and Asia.
Biological Invasions. doi 10.1007/s10530-009-9538-9
Cavalcaselle B (1983) Osservazioni sulla diff usione di Phoracantha semipunctata F. in Sardegna, Sicilia e nell’Italia centro-meridionale. Atti XIII Congresso Nazionale Italiano di
Entomolo-gia, Sestriere-Torino, 455–464.
Černý Z (2002) A contribution to the understanding of Beetles of the family Coleoptera: Cer-ambycidae in Greece. Studie a Zprávy Okresniho Muzea Praha-Vychod 14: 3–26.
Chikatunov V, Kravchenko, Dorchin Y, Müller G (2006) Th e xylophagous beetles (Bupresti-dae, Lycti(Bupresti-dae, Bostrichi(Bupresti-dae, Anobii(Bupresti-dae, Cerambyci(Bupresti-dae, Scolyti(Bupresti-dae, Platypodidae) collected in the Israeli light trap survey and their association with the major phyto-geographical zones of Israel. Esperiana. Buchreiche zur Entomologie 12: 313–320.
Chikatunov V, Pavlicek T, Nevo E (1999) Coleptera of “Evolution Canyon”. Lower Nahal Oren, Mount Carmel, Israel. Pensoft Series faunistica 14: 1–174.
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 207
Cocquempot C (1993) Notes de chasse et Observations diverses – Nouvelle extension pour
Phoracantha semipunctata (F.) (Col. Cerambycidae). L’Entomologiste 49: 37.
Cocquempot C (2007) Alien longhorned beetles (Coleoptera Cerambycidae): Original inter-ceptions and introductions in Europe, mainly in France, and notes about recently impor-ted species. Redia 89: 35–50.
Cocquempot C, Debreuil M (2006) Xylotrechus stebbingi Gahan 1906 et Phoracantha
semipun-ctata (Fabricius, 1775): historique de l’expansion française de deux envahisseurs
(Coleop-tera Cerambycidae). Rutilans 9: 85–89.
Cocquempot C, Grousset C, Pellegrin D (2007) Distribution française de Lucasianus levaillantii (Lucas, 1846) et des sous-espèces d'Icosium tomentosum (Lucas, 1854) (Coleoptera
Ceramby-cidae). Rutilans 10: 35–39.
Cocquempot C, Hérard F, Reynaud P (2003) Les longicornes asiatiques Anoplophora
glabripen-nis et Anoplophora chinensis, une menace sérieuse pour l’arboriculture fruitière, les plantes
d’ornement et les forêts françaises. Phytoma – La Défense des Végétaux 561: 24–28. Cocquempot C, Sama G (2004) L’expansion circumméditerranéenne de Phoracantha recurva
(Newman, 1840) (Coleoptera, Cerambycidae). L’ Entomologiste 59: 209–214.
Colombo M, Limonta L (2001) Anoplophora malasiaca Th omson (Coleoptera Cerambycidae Lamiinae Lamiini) in Europe. Bollettino di Zoologia agraria e di Bachicoltura II Ser 33: 65–68.
Dauber D, Mitter H (2001) Das erstmalige Auftreten von Anoplophora glabripennis Motschul-sky 1853 auf dem europaïschen Festland (Coleoptera: Cerambycidae: Lamiinae). Beiträge
zur Naturkunde Oberösterreichs 10: 503–508.
De Fluiter HJ (1950) Monochamus (Monohammus) galloprovincialis Oliv. (Col.Cerambicidae).
Entomologische Berichten 13: 135–136.
Delvare G, Bon M-C, Hérard F, Cocquempot C, Maspero M, Colombo M (2004) Descrip-tion of Aprostocetus anoplophorae n. sp. (Hymenoptera: Eulophidae), a new egg parasitoid of the invasive pest Anoplophora chinensis (Förster) (Coleoptera: Cerambycidae). Annales de
la Société entomologique de France 40: 227–233.
Demelt C (1974) Zusammenfassung und revision der cerambycidenfauna der Canarischen Inseln. Nouvelle Revue d’Entomologie 4: 227–236.
Dioli P, Vigano C (1990) Presenza in Valtellina di un cerambice nuovo per la Fauna italiana: Xylotrechus stebbingi Gahan, 1906 (Insecta, Coleoptera, Cerambycidae). Il Naturalista
Valtellinese 1: 7–10.
Duff y EAJ (1953a) A Monograph of the immature stages of British and imported Timber Beetles (Cerambycidae). London: British Museum (Natural History). 350 pp.
Duff y EAJ (1953b) Trinophylum cribratum Bates (Col., Cerambycidae). A new Indian pest of British timber yards. Entomologist’s Gazette 4: 254–264.
EPPO (2004). First report of Anoplophora glabripennis in Germany. EPPO Reporting Services 5: 2004/072.
EPPO (2006). Recent interceptions of Anoplophora species in United Kingdom. EPPO
Repor-ting Services 5: 2006/098.
EPPO (2008) Premier signalement de Saperda candida en Allemagne: addition à la Liste d’Alerte de l’OEPP. EPPO Reporting services 7: 2008/139.
EPPO (2009a) De plus amples détails sur le foyer d’Anoplophora chinensis à Roma (IT). EPPO
Reporting Services 3: 2009/048.
EPPO (2009b) Anoplophora chinensis found on Cornus and Crataegus in the Netherlands.
EPPO Reporting Services 9: 2009/173
Evans S, Evans HF, Ikegami M (2008) Modeling PWN-Induced Wilt Expression: A Mecha-nistic Approach. In: Mota M, Vieira P (Eds) Pine Wilt Disease: A Worldwide Th reat to
For-est Ecosystems. Th e Netherlands: Springer Science+Business Media B.V., 259–278.
EPPO (2010) Anoplophora chinensis found again in the Netherlands. EPPO Reporting Services 2 : 2010/025
Fauna Europaea. http://www.faunaeur.org.
Fauvel (1884) Les Longicornes Gallo-Rhénans (suite) - Catalogue. Revue d’Entomologie 3: 317–390.
Frank H (2002) Pathways of arrival. In: Hallman GJ, Schwalbe CP Invasive Arthropods in
Agri-culture: Problems and Solutions. Enfi eld, UK: Sciences Publishers Inc., 71–86.
Friedman ALL, Rittner O, Chikatunov VI ( 2008) Five new invasive species of longhorn beetles (Coleoptera: Cerambycidae) in Israel. Phytoparasitica 36: 242–246
Gilmour EF (1948) Trinophylum cribratum Bates (Col., Cerambycidae) new to Britain.Th e
Entomologist’s monthly Magazine, 84: 12–16.
Haack RA, Poland TM, Gao RT (2000) Th e United States experience with the exotic Cer-ambycid Anoplophora glabripennis: Detection, Quarantine, and Control. In: Quarantine
pests, risks for the forestry sector and their eff ects on foreign trade. Proceedings on CD-Room of Silvotecna Conference 14, 27–28 June 2000. Concepcion, Chile: CORMA. 12 pp.
Haack RA, Law KR, Mastro VC, Ossenbruggen SH, Raimo BJ (1997) New York’s battle with the Asian Long-Horned Beetle. Journal of Forestry 95: 11–15.
Haack RA, Hérard F, Sun JH, Turgeon JJ (2010) Managing invasive populations of Asian longhorned beetle and Citrus longhorn beetle: A worldwide perspective. Annual Review of
Entomology 55: 521–546.
Hérard F, Ciampitti M, Maspero M, Krehan H, Benker U, Boegel C, Schrage R, Bouhot-Delduc L, Bialooki P (2006) Anoplophora species in Europe: infestations and management processes. Bulletin OEPP/EPPO Bulletin 36: 470–474.
Hérard F, Maspero M, Ramualde N, Jucker C, Colombo M, Ciampitti M, Cavagna B (2009)
Anoplophora glabripennis - Eradication programme in Italy (April 2009). http://www.eppo.
org/QUARANTINE/ anoplophora_glabripennis/ANOLGL_IT.htm.
Hérard F, Roques A (2009) Current status of Anoplophora spp. in Europe and an update on suppression eff orts. In: Gottschalk KW (Ed) Proceedings of the 20th U.S. 20th USDA
Interagency Research Forum on Invasive Species Annapolis, Maryland, 13- 16 January 2009. Gen. Tech. Rep. NE-337. Morgantown, USA: USDA Forest Service, Northeastern Research Station.
Heyrovský L (1930) Entomologické Prírucky XV. Catalogus Coleopterum Cechosloveniae – 1. Cerambycidae. Casopis Ceskoslovenské spolecnosti Entomologické 26–27: 1–61.
Heyrovský L (1951) Catalogus Coleopterum Cechosloveniae. Supplementum. Cerambycidae (Tesaríci). Casopis Cekqslovenské spolecnosti Entomologické, 48: 43–53.
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 209
Heyrovský L, Sláma G (1992) Tesaríkovití – Coleoptera, Cerambycidae. Zlín, Czech Republic: Kabourek. 367pp.
Horion A (1974) Faunistik der mitteleuropäischen Käfer 12: Cerambycidae – Bockkäfer. Überlin-gen-Bodensee. 228 pp.
Hunt T, Bergsten J, Levkanicova Z, Papadopoulou A, Saint-John O, Wild R, Hammond P M, Ahrens D, Balke M, Caterino M S, Gómez-Zurita J, Ribera I, Barraclough TG, Bocakova M, Bocak L, Vogler AP (2007) A Comprehensive Phylogeny of Beetles Reveals the Evolu-tionary Origins of a Super-radiation. Science, 318: 1913–1916.
Ilić N (2005) [Longhorn beetles (Coleoptera, Cerambycidae) from Serbia. Faunistic review] (in Serbian) Belgrade: SZGR “Joksimović”.180pp.
Jucker C, Tantardini A, Colombo M (2006) First record of Psacothea hilaris (Pascoe) in Europe (Coleoptera Cerambycidae lamiinae Lamiini). Bollettino di Zoologia agraria e di
Bachicol-tura 38: 187–191.
Jucker C, Maspero M, Colombo M (2007) Principali fi tofagi in ambiente urbano e loro conte-nimento. Informatore fi topatologico: 22–25.
Kawai Miho, Etsuko Shoda-Kagaya, Tadashi Maehara T, Zhou ZH, Lian CL, Iwata RT, Ya-mane A, Hogetsu T (2006). Genetic structure of pine sawyer Monochamus alternatus (Co-leoptera: Cerambycidae) populations in Northeast Asia: consequences of the spread of Pine Wilt Disease. Environmental Entomology 35, 569–579.
Köhler F (2000) Erster Nachtrag zum „Verzeichnis der Käfer Deutschlands“. Entomologische
Nachrichten und Berichte 44: 60–84.
Komazaki S, Sakagami Y (1989) Capture-Recapture Study on the Adult Population of the White Spotted Longicorn Beetle, Anoplophora malasiaca (Th omson) (Coleoptera: Ceram-bycidae) in a Citrus Orchard. Applied Entomology and Zoology, 24: 78–84.
Korcynski I (1985) Th e Biology of Nathrius brevipennis (Muls.) (Coleoptera, Cerambycidae) in Poland. Polskie Pismo Entomologiczne 55: 73–78.
Lawrence JF, Newton AF (1995) Families and subfamilies of Coleoptera (with selected genera, notes, references and data of family-group names. In: Pakaluk J, Ślipiński SA (Eds)
Biol-ogy, Phylogeny, and Classifi cation of Coleoptera. Papers celebrating the 80th Birthday of Roy
Crowson. Warzawa: Muzeum i Instytut Zoologii PAN, 779–1006.
Lemos-Perreira AB (1978) Sobre o aparecimento em Portugal de um Coleóptero Longicórnio exótico, do género Acrocinus. Publicações do Instituto de Zoologia «Dr. Augusto Nobre»,
Fac-uldade de Ciências do Porto 138: 1–13.
Linnaeus C von (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera,
species, cum characteribus, diff erentiis, synonymis, locis. 10th Edition. Holmiæ, Impensis
di-rect. Laurentii Salvii (Salvius)
Lucht WH (1987) Die Käfer Mitteleuropas. Katalog. Krefeld, Germany: Goecke & Evers Ver-lag. 342pp.
MacLeod A, Evans HF, Baker RHA (2002) An analysis of pest risk from an Asian longhorn beetle (Anoplophora glabripennis) to hard wood trees in the European community. Crop
Protection, 21: 635–645.
Maspero M, Cavalieri G, D’Angelo G, Jucker C, Valentini M, Colombo M, Hérard F, Lopez J, Ramualde N, Ciampitti M, Caremi G, Cavagna B (2007a) Anoplophora chinensis -
Eradi-cation programme in Lombardia (Italy) http://www.eppo.org/QUARANTINE/anoplo-phora_chinensis/chinensis_IT_2007.htm.
Maspero M, Jucker C, Colombo M (2007b) First record of Anoplophora glabripennis (Mots-chulsky) (Coleoptera Cerambycidae Lamiinae Lamiini) in Italy. Bollettino di Zoologia
agra-ria e di Bachicoltura, Serie II 39: 161–164.
Mayet V (1905) Le Lucasianus levaillanti Lucas (Col.), sa découverte en France et ses mœurs.
Bulletin de la Société entomologique de France: 277–279.
Mazzeo G, Siscaro G (2007) Presenza di Phoracantha recurva su eucalipto in Sicilia. Informatore
fi topatologico: 35–37.
Mendizábal M (1944) Cerambícidos de interés agricola. Boletín de Patología vegetal y Ento-mología agrícola 11: 463–476.
Méquignon A (1935) Voyage de MM. L. Chopard et A. Méquignon aux Açores (aoùt- sep-tembre 1930) – VIII. Coléoptères américains acclimatés aux Açores. Bulletin de la Société
entomologique de France 13: 203–204.
Mifsud D (2002) Longhorns beetles of the Maltese Islands (Coleoptera, Cerambycidae). Th e
Central Mediterranean Naturalist 3: 161–168.
Mifsud D, Booth R (1997) Further contributions to the knowledge of the Longhorn Beetles (Coleoptera: Cerambycidae) of the Maltese Islands. Th e Central Mediterranean Naturalist 2: 35–36.
Mifsud D, Dandria D (2002) Introduction and establishment of Phryneta leprosa (Fabricius) (Coleoptera, Cerambycidae) in Malta.Th e Central Mediterranean Naturalist 3: 207–210. Miquel ME (2008) Th e presence of Phoracantha recurva (Newman) (Coleoptera:
Cerambyci-dae) in France and how to diff erentiate if from P. semipunctata. British Journal of Natural
History 21: 193–194.
Mota MM, Braasch H, Bravo MA, Penas AC, Burgermeister W, Metge K and Sousa E (1999) First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1: 727– 734.
Napp DS (1994) Phylogenetic relationships among the subfamilies of Cerambycidae (Coleop-tera – Chrysomeloidea). Revista Brasileira de Entomologia 38: 265–419.
Nolte O, Krieger D (2008) Nachweis von Saperda candida Fabricius 1787 auf Fehmarn – eine weitere, bereits in Ansiedlung befi ndliche, eingeschleppte Käferart in Mitteleuropa.
DgaaE-Nachrichten 22: 133–136.
Nüssler H (1961) Ein beständiges Vorkommen des nearktischen Bockkäfers Parandra brunnea Fabr. (Col. Cerambycidae) in Deutschland. Abhandlungen und Berichte aus dem Staatliches
Museum für Tierkunde in Dresden 26: 125–130.
Orousset J (1984) Phoracantha semipunctata Fabr., un ravageur des eucalyptus présent en Corse (Col. Cerambycidae). Nouvelle Revue d’Entomologie 1: 322.
Orousset J (1991) Présence aux Îles Canaries du ravageur des Eucalyptus Phoracantha
semi-punctata (Col. Cerambycidae). Nouvelle Revue d‘Entomologie 8: 76.
Orousset J (2000) Phoracantha recurva Newman sera-t-il à ajouter à la faune de France ? (Co-leoptera, Cerambycidae). Le Coléoptériste 39: 129–131.
Özdikmen H (2008) A nomenclatural act: some nomenclatural changes on palaearctic long-horned beetles (Coleoptera: Cerambycidae). Munis Entomology and Zoology 3: 707–715.
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 211
Palmeri V, Campolo O (2006) Sulla presenza di Phoracantha recurva e Phoracantha
semipunc-tata F. (Coleoptera Cerambycidae) in Calabria. Bollettino di Zoologia agraria e di Bachicol-tura 38: 251–254.
Pellegrin D (1990) Icosium tomentosum Lucas, espèce nouvelle pour la France continentale (Col. Cerambycidae Cerambycinae). L’Entomologiste, t. 46 (4): 167–169.
Pellegrin D, Cocquempot C (2001) Réhabilitation de la présence en France de Lucasianus
le-vaillanti (Lucas, 1849) et nouvelles données géographiques. Biocosme mésogéen 18: 27–31.
Pérez Moreno I (2001) Plagas introducidas em España peninsular em la segunda mitad del siglo
XX. ARACNET 4 http://entomologia.rediris.es/aracnet/num4/entomap/index.htm.
Picard F (1937) Sur les Cérambycides de Madère et des îles voisines. Compte rendu sommaire
des Séances de la Société de Biogéographie 13: 70–72.
Pil N, Stojanović D (2005) Some rare longhorn beetles (Coleoptera: Cerambycidae) without Protection on the national level found on Mt. Fruška Gora, Serbia. Archives of Biological
Science Belgrade 57: 137–142.
Plavilstshtikov NN (1934) Bestimmungs-Tabellen der europaïschen Coleopteren. 112 Heft.
Cer-ambycidae III. Teil. Cerambycinae: Cerambycini II1 (Callichromina, Rosaliina, Callidiina),
Troppau, Germany: Verlag Edmund Reitter. 230pp.
Plaza Lama J (1990) Primera cita para la fauna ibérica de Lucasianus levaillanti (Lucas, 1849) (Col. Cer.). Boletín del Grupo Entomológico de Madrid 5: 73–75.
Reineck G (1919) Die Insekten der Mark Brandenburg. 2. Coleoptera. Cerambycidae. Beiheft
der Deutsche Entomologische Zeitschrift 63: 1–92.
Rossem G. van, van de Bund CF, Burger HC, de Goff au LJW (1981) Bijzondere aantastingen door insekten in 1980. Entomologische Berichten 41: 84–87.
Ruiz JL, Barranco P (1998) Phoracantha recurva Newman, 1840, nueva especie plaga para la Región Mediterránea (Coleoptera: Cerambycidae). Boletín de la Asociación Española de
Entomología 22: 227–228.
Sama G (1984) Fauna d’Italia Vol. XXV. Coleoptera Cerambycidae, Catalogo topographico e
sino-nimico. Bologna, Italia: Edizioni Calderini. 216 pp.
Sama G (2002) Atlas of the Cerambycidae of Europe and Mediterranean area. Vol 1: Northern,
Western, Central and Eastern Europe. Zlín, Czech Republic : Kabourek. 173 pp.
Sama G (2006a) Notes on Taeniotes scalaris (Fabricius, 1781) and T. cayennensis (Th omson, 1859) (Coleoptera: Cerambycidae) in the Azorean Archipelago. Boletín de la Sociedad
En-tomológica Aragonesa 39: 231–234.
Sama G (2006b) Taxonomical remarks on Xylotrechus smei (Castelnau & Gory, 1841) and X.
stebbingi Gahan, 1906 and their distribution in Western Palaearctic region. Doriana 7: 1–6.
Sama G, Bocchini R (2003) Phoracantha recurva (Newman, 1840) (Insecta Coleoptera Ce-rambycidae), prima segnalazione per la fauna italiana.Quaderno di Studi e Notizie di Storia
naturale della Romagna 18: 168.
Sama G, Buse J, Orbach E, Friedman ALL, Rittner O, Chikatunov V (2010) A new catalo-gue of the Cerambycidae (Coleoptera) of Israel with notes on their distribution and host plants. Munis Entomology and Zoology 5: 1–51.
Šefrová H, Laštůvka Z (2005) Catalogue of alien animal species in the Czech Republic. Acta
Sama G, Cocquempot C (1995) Note sur l’extension européenne de Xylotrechus stebbingi Ga-han, 1906 (Coleoptera, Cerambycidae, Clytini). L’Entomologiste 51: 71–75.
Schembri S, Sama G (1986) Th e Cerambycidae of the Maltese Islands (Coleoptera). Bollettino
della Società entomologica italiana 118: 93–100.
Schmidt G, Schmidt HU (1990) Ein ungewöhnlicher “Gast” aus einem Bonsai- Bäumchen:
Anoplophora malasiaca Th oms. (Coleoptera: Cerambycidae). Mitteilungen des
Internation-alen Entomologischen Vereins 15: 69–75.
Serrano ARM (1982) Contribuciao para o conhecimato do poro amerto, distribuciao e oviger dos Coleopteros do Archipelago dos Açores. Boletim do Museu Municipal do Funchal 34: 67–104.
Sliwinski Z (1958) Nathrius brevipennis (Muls.) Bosq. a new species of beetle (Coleoptera, Cerambycidae) for fauna Polands. Polskie Pismo Entomologiczne 28: 129–131.
Smith MT, Bancroft J, Li G, Gao R, Teale S (2001) Dispersal of Anoplophora glabripennis (Cerambycidae). Environmental Entomology 30: 1036–1040
Speight MCD (1988) Th e Irish Cerambycid fauna (Coleoptera : Cerambycidae). Bulletin of the
Irish biogeographical Society 11: 41–76.
Sýkorová M (2008) Molecular phylogeny of subfamilies Spondylidinae and Lepturinae based on
mitochondrial 16S rDNA, BCc. PhD Th esis. České Budĕjovice, Czech Republic: Faculty of
Science, University of South Bohemia. 34pp.
Szeoke K, Hegyi T (2002) A szolo uj kartevoje az amerikai darazscincer ( Neoclytus acuminatus Olivier). [A new grapevine pest: redheaded ash borer ( Neoclytus acuminatus Olivier)].
No-venyvedelem Majus 38: 268–269.
Tassi F (1969) Un longicorne australiano per la prima volta in Italia (Col. Cerambycidae).
Bolletino dell’ Associazione Romana di Entomologia 24: 69–71.
Teunissen (2002) Waarnemingen van Xylotrechus stebbingi in Griekenland. Een Aziatische wes-penboktor die zich in korte tijd heeft verbreid in het Middellandsee-Zeegebied.
Entomolo-gische Berichten (Amsterdam) 62: 57–58.
Tomiczek C, Hoyer-Tomiczek U (2008). Biology Studies Relevant to the Vector Role of
Mono-chamus Species for PineWood Nematode. In: Mota M, Vieira P (2008) Pine Wilt Disease:
A Worldwide Th reat to Forest Ecosystems. Dordrecht, Th e Netherlands: Springer, 215–221.
Verdugo Páez AV (2004) Los cerambícidos de Andalucía (Coleoptera: Cerambycidae). Cór-doba, Spain: Sociedad Andaluza de Entomología. Monográfi co 1. 148 pp.
Villiers A (1967) Coléoptères Cérambycides de l’île de Chypre. L’Entomologiste 23: 63–64. Villiers A (1979) Un nouveau Clytini en France ? L’Entomologiste 35: 144.
Vincent R (2007) Catalogue des Coléoptères du département de la Saône-et-Loire (F-71). Volume
I: Cerambycidae. Terre Vive, Supplément Hors-Série: 183 pp.
Vives E (1995) Notas sobre longicornios ibéricos (V). Cerambícidos importados o aclimatados en la Península Ibérica (Coleoptera, Cerambycidae). Zapateri, Revista aragonese de
Ento-mologia 5: 165–174.
Wang Q (1995) A Taxonomic Revision of the Australian Genus Phoracantha Newman (Cole-optera: Cerambycidae). Invertebrate Taxonomy, 9: 865–958.
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 213
Weidner H (1973) Insekten als Schädlinge an Drogen in Apotheken un Drogerien. Anzeiger
für Schädlingskunde und Pfl anzen- und Umweltschutz vereinigt mit Schädlingsbekämpfung
46: 177–180.
Weyers M (1875) Communication sur Leptidea brevipennis à Bruxelles. Annales de la Société
entomologique de Belgique, Comptes rendus des Séances 18: 86–87.
Weyers M (1876) Communication à propos de Longicornes nouveaux pour la faune belge.
Annales de la Société entomologique de Belgique, Comptes rendus des Séances 19: 65.
Wiel P van der (1956) Bijdrage tot de kennis der Nederlandse kevers, IV. Tijdschrift voor Entomologie, t. 99 (1–2): 1–21.
Winkler A (1932) Catalogus Coleopterum regionis palaearcticae. Wien: Albert Winkler Verlag: 1135–1226.
Wittenberg R (Ed) (2005) An inventory of alien species and their threat to biodiversity and economy in Switzerland. Délemont, Switzerland: CABI Bioscience Switzerland Centre, Report to Swiss Agency for Environment, Forest and Landscape. 417 pp.
Wollaston TV (1863) On the Canarian Longicorns. Journal of Entomology 8: 99–110. Worner SP (2002) Predicting the invasive potential of exotic insects. In: Hallman GJ, Schwalbe
CP Invasive Arthropods in Agriculture: Problems and Solutions. Enfi eld, UK: Sciences Pub-lishers Inc., 119–137.
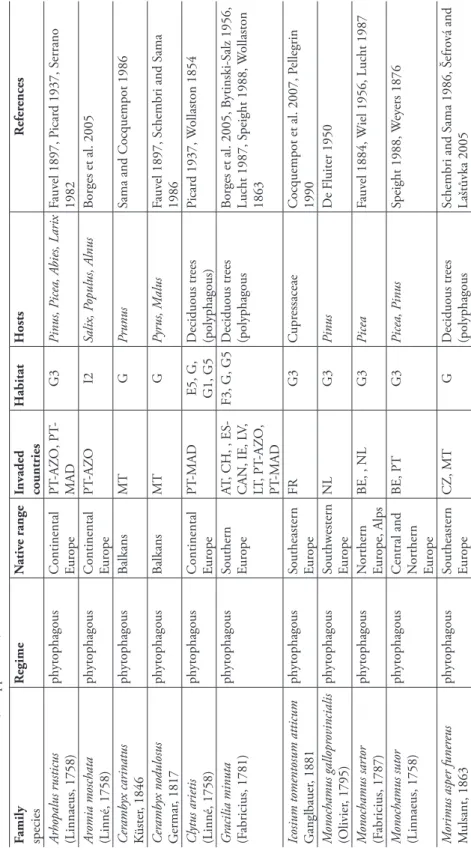
F amily Species Status R egime N ativ e range 1st r ecor d in E u rope In vaded countries H abitat H osts R efer ences A canthoder es jaspidea G ermar , 1824 A phyto -phagous B razil 1880, PT -AZ O P T -AZ O I2 A cacia, Albizzia
Borges et al. 2005, Méquignon 1935, S
errano 1982 A cr ocinus longimanus (Linnaeus, 1758) A phyto- phagous B razil 1977, PT PT , PT -MAD I2 M oraceae, A pocynaceae Lemos-P err eira 1978 , V iv es 1995 A noplophor a chinensis (F örster , 1848) (= A . malasiaca Th ompson, 1865) A phyto- phagous China S outh-Central 2000, IT IT , NL FB, F A, I2, G A cer , B etula, C arpinus C itr us, Cor ylus, R osa and deciduous shr ubs (polyphagous)
Cocquempot 2007, Colombo and Limonta 2001, 2009a, EPPO 2009b
, E vans et al. 2008, H érar d et al. 2006 A noplophor a glabripennis (M otschulsky , 1853) A phyto- phagous China S outh-Central 2001, A T A T , DE, FR, IT FB, F A, I A cer , A esculus, B etula, Carpinus, Fagus, P opulus, Salix Car ter et al. 2009,
Cocquempot 2007, Cocquempot et al. 2003, Dauber and M
itter 2001, EPPO 2004, H érar d et al. 2006, 2009 C allidiellum ru fi penne (M otschulsky , 1860) A phyto- phagous Eastern Asia, Japan
1906, FR ES, FR, IT FA, FB, G1, G5, J4 C upr essaceae (C upr essus macr ocarpa ) B ahillo and I turr ondobeitia 1995, Campadelli and S ama 1988, Cocquempot 2007 Chlor ophor us annularis (F abricius, 1787) A phyto- phagous Asia- Temperate 1991, ES ES G B amboo V iv es 1995 C yr thognathus forfi catus (F abricius, 1792) A phyto- phagous Africa 1872, MT MT U U nkno wn B er tolini 1872 D er olus mauritanicus B uquet, 1840 A phyto- phagous N or thern Africa 1884, FR ES ?, FR ? E7, F5, F8, FB, I2, X11 N erium oleander B rustel et al. 2002, F auv el 1884, M endizábal 1944, V er dugo 2004 T a b le 8.1.1.
List and characteristics of the Ceramb
ycidae species alien to E
ur
ope. S
tatus: A: Alien to E
ur
ope; C: cr
yptogenic species. Coun
tr y codes abbr eviations refer to ISO 3166 (s ee appendix I). H abitat abbr eviations r
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 215 F amily Species Status R egime N ativ e range 1st r ecor d in E u rope In vaded countries H abitat H osts R efer ences D er oplia albida (B rullé, 1838) A phyto- phagous Canar y Islands 1988, ES ES E7, F6, FB, G5 Pelargonium V iv es 1995 L ucasianus lev aillantii (L ucas, 1846) A phyto- phagous N or thern Africa 1905, FR ES, FR, PT FA, G, FB C upr essus B rustel et al. 2002, Cocquempot et al. 2007, May et 1905, P ellegrin and Cocquempot 2001, P laza Lama 1990, V iv es 1995 N eoclytus acuminatus (F abricius, 1775) A phyto- phagous South- Central U.S.A.
1908, IT
CH, CZ, DE, FR, HR, HU, IT, ME, PT
-MAD, RS, SI FB, G, G1, G5, I2, X11 U lmus, Fr axinus, Juglans B ijaoui 1980, B rustel et al. 2002, Cocquempot 2007, H eyr ovský 1951, I lić 2005, P icar d 1937, P il and Stojano vić 2005, R eineck 1919, S ama 1984, T assi 1969, V illiers 1979, W inkler 1932, W ittenberg 2005 O xymer us aculeatus lebasi D upont, 1838 C phyto- phagous U nkno wn U nkno wn ES-CAN U C alophyllum Alluaud 1935 Pa randr a br unnea (F abricius, 1789) A phyto- phagous N or th America 1916, DE DE G, J1 T ilia, P opulus , deciduous trees G rämer 1961, N üssler 1961 P hor acantha r ecur va N ewman, 1840 A phyto- phagous A ustralia 1992, IT ES, GR, IL, IT , IT -SAR, IT -SIC, MT , PT G1 E ucalyptus B er cedo and B ahillo 1998, B er cedo and B ahillo 1999,
Černý 2002, Cocquempot 2007, Cocquempot and S
ama 2004, F riedman et al. 2008, M azz eo and S iscar o 2007, M ifsud 2002, M iquel 2008, O rousset 2000, P almeri and Campolo 2006, P ér ez M or eno 2001, R uiz and B arranco 1998,
Sama and Bocchini 2003, Sama et al. 2010,
W
F amily Species Status R egime N ativ e range 1st r ecor d in E u rope In vaded countries H abitat H osts R efer ences P hor acantha semipunctata (F abricius, 1775) A phyto- phagous A ustralia 1948, IL CY , FR,
FR-COR, ES, ES-CAN, GR, IL, IT
, IT -SAR, IT -SIC, MT , PT , PT -MAD FB, G, G1, G5, I2, X11 E ucalyptus B erger 1992, B rustel et
al. 2002, Cadahia 1980, Cav
alcaselle 1983, Černý
2002, Cocquempot 1993, Cocquempot 2007, Cocquempot and S
ama 2004,
M
ifsud and Booth 1997,
O rousset 1984, O rousset 1991, S ama et al. 2010, T eunissen 2002, V iv es 1995, W ang 1995 P hr yneta lepr osa (F abricius, 1775) A phyto- phagous South Tropical Africa
1997, FR FR, MT G M or us nigr a M ifsud and D andria 2002, V incent 2007 Taeniotes cay ennensis Th omson, 1859 A phyto- phagous Central America 1858, PT PT -AZ O U T ropical tr ees Sama 2006a T rinophylum cribr atum (B ates, 1878) A phyto- phagous India U nkno wn GB I2 D eciduous tr ees, Larix , P inus (polyphagous) Du ff y 1953b , G ilmour 1948 X ylotr echus stebbingi G ahan, 1906 A phyto- phagous Central Asia 1990, IT CH, CY ,
DE, FR, GR, GR-CRE, GR-NEG, GR- SEG, IL, IT
, IT -SAR FB, G, G1, G5, I2, X11 A lnus, F icus, M or us, P opulus
Cocquempot 2007, Cocquempot and D
ebr euil 2006, D ioli and V igano 1990, K öhler 2000, S ama 2006b , Sama et al. 2010, Š efr ov á and Laštůvka 2005, T omicz ek and H oy er-T omicz ek 2008, W ittenberg 2005
Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 217 F amily species R egime N ativ e range In vaded countries H abitat H osts R efer ences A rhopalus rusticus (Linnaeus, 1758) phytophagous Continental Eur ope PT -AZ O, PT -MAD G3 P inus, P icea, A bies, Larix Fauv el 1897, P icar d 1937, S errano 1982 A romia moschata (Linné, 1758) phytophagous Continental Eur ope PT -AZ O I2 Salix, P opulus, A lnus Borges et al. 2005 C er amb yx carinatus K üster , 1846 phytophagous B alkans MT G P runus
Sama and Cocquempot 1986
C er amb yx nodulosus G ermar , 1817 phytophagous B alkans MT G P yr us, M alus Fauv el 1897, Schembri and S ama 1986 Clytus arietis (Linné, 1758) phytophagous Continental Eur ope PT -MAD E5, G, G1, G5 D eciduous tr ees (polyphagous) P icar d 1937, W ollaston 1854 G racilia minuta (F abricius, 1781) phytophagous Southern Eur ope A T , CH, , ES-CAN, IE, L V , LT , PT -AZ O, PT -MAD F3, G, G5 D eciduous tr ees (polyphagous Borges et al. 2005, B ytinski-S alz 1956, L ucht 1987, S peight 1988, W ollaston 1863 Icosium tomentosum atticum G anglbauer , 1881 phytophagous Southeastern Eur ope FR G3 C upr essaceae Cocquempot et al. 2007, P ellegrin 1990 M onochamus gallopr ovincialis (O livier , 1795) phytophagous Southw estern E ur ope NL G3 P inus D e F luiter 1950 M onochamus sar tor (F abricius, 1787) phytophagous N or thern E ur ope, Alps BE, , NL G3 P icea Fauv el 1884, W iel 1956, L ucht 1987 M onochamus sutor (Linnaeus, 1758) phytophagous Central and Nor thern E ur ope BE, PT G3 P icea, P inus Speight 1988, W ey ers 1876 M orimus asper funer eus M ulsant, 1863 phytophagous Southeastern Eur ope CZ, MT G D eciduous tr ees (polyphagous Schembri and S ama 1986, Š efr ov á and Laštůvka 2005 T a b le 8.1.2.
List and characteristics of the Ceramb
ycidae species alien
in E ur ope. Countr y codes abbr eviations r
efer to ISO 3166 (see appendix I). H
abitat abbr
e-viations r
F amily species R egime N ativ e range In vaded countries H abitat H osts R efer ences N athrius br evipennis (M ulsant, 1839) phytophagous Southw estern E ur ope A T , BE, CH, CZ, DE, GB, IE, L U, L V , PL, PT -AZ O F3 D
eciduous and conifer
tr
ees (polyphagous)
A
dlbauer 2006, Borges et al. 2005,
Du ff y 1953a, H eyr ovský 1930, K or cynski 1985, L ucht 1987, S liwinski 1958, S peight 1988, W eidner 1973, W ey ers 1875 P hymatodes testaceus (Linné, 1758) phytophagous Continental Eur ope PT -AZ O G D eciduous and fr uit tr ees, pr eferably on Q uer cus Fauv el 1897, P icar d 1937, W ollaston 1854 Poecilium lividum (R ossi, 1794) phytophagous Southeastern Eur ope BE, CH, CZ, DE, L U, NL G,J1 Q uer cus, C astanea L ucht 1987, H eyr ovský and S láma 1992, H orion 1974, Š efr ov á and Laštůvka 2005, W ittenberg 2005 Rhagium inquisitor (Linné, 1758) phytophagous Continental Eur ope IE G3 Conifers (P inus, P icea, A bies, Larix); deciduous tr ees ( B etula, F agus, Q uer cus ) Speight 1988 R osalia alpina (Linné, 1758) phytophagous Central Eur ope, Alps MT G, I2, J1 Fagus , and other deciduous tr ees H
orion 1974, Schembri and S
ama 1986 Stictoleptur a rubr a (Linné, 1758) phytophagous Central Eur ope PT -AZ O G 3 Conifers ( P inus, P icea, A bies, Larix ) Borges et al. 2005 St romatium unicolor (O livier , 1795) phytophagous Southeastern Eur ope PT -MAD G D eciduous tr ees
(mostly) and conifers (polyphagous)
Fauv el 1897, P icar d 1937 T richofer us fasciculatus (F aldermann, 1837) phytophagous Southeastern Eur ope CH, PT -MAD G D eciduous tr ees (polyphagous) Allenspach 1973, P icar d 1937 T richofer us griseus (F abricius, 1792) phytophagous Southeastern Eur ope CZ G F icus, P istacia, R osa Šefr ov á and Laštůvka 2005 X ylotr echus ar vicola (O livier , 1795) phytophagous Southeastern Eur ope SP-CAN G D eciduous tr ees (polyphagous) D emelt 1974