HAL Id: tel-01222760
https://tel.archives-ouvertes.fr/tel-01222760
Submitted on 30 Oct 2015HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Role of human gamma-delta T lymphocytes in the
instruction of the adaptive immune response against
Plasmodium falciparum infection.
Jennifer Ruth Howard
To cite this version:
Jennifer Ruth Howard. Role of human gamma-delta T lymphocytes in the instruction of the adaptive immune response against Plasmodium falciparum infection.. Immunology. Université de Bordeaux, 2015. English. �NNT : 2015BORD0110�. �tel-01222760�
THÈSE PRÉSENTÉE POUR OBTENIR LE GRADE DE
DOCTEUR DE
L’UNIVERSITÉ DE BORDEAUX
ÉCOLE DOCTORALE : Sciences de la Vie et de la Santé
SPÉCIALITÉ : Microbiologie-Immunologie
Par Jennifer HOWARD
Role of human gamma-delta T lymphocytes in
the instruction of the adaptive immune response
against Plasmodium falciparum infection.
Soutenue publiquement le 16/07/15
!
Membres du jury :
Pr. Jean-Francois MOREAU
Président
Pr. Lars HVIID
Rapporteur
Dr. Matthias EBERL
Rapporteur
Pr. Marita TROYE-BLOMBERG
Examinateur
Dr. Odile MERCEREAU-PUIJALON
Examinateur
Dr. Julie DECHANET-MERVILLE
Directeur
!
! ! 2! !
! ! 3! ! ! ! ! ! ! ! ! ! ! ! !
In!memory!of!Charlotte.
!
!
! ! 4!
Résumé&en&Français&:&
Rôle& des& lymphocytes& T& gamma& delta& dans& l’induction& de& la&
réponse&immunitaire&adaptative&dans&un&contexte&d’infection&
par&Plasmodium*falciparum.&
Introduction:&
Le!paludisme,!aussi!appelé!malaria,!est!la!maladie!parasitaire!la!plus!répandue!au!monde! et!surtout!dans!les!pays!en!voie!de!développement.!Malgré!les!traitements!qui!ont!permis! une! baisse! de! 42%! de! la! mortalité! en! 2000! (OMS! 2014),! 627! 000! personnes! (surtout! des! enfants)! meurent! du! paludisme! chaque! année.! Avec! la! dissémination! progressive! de! moustiques!résistants!aux!insecticides!et!de!parasites!résistants!aux!drogues,!de!nouveaux! remèdes! et! vaccins! sont! nécessaires.! Cependant,! pour! trouver! des! vaccins! plus! spécifiquement! dirigés! contre! ces! parasites,! il! est! indispensable! de! bien! comprendre! le! mécanisme!de!réponse!immunitaire!à!l’infection!par!Plasmodium.!!
Plusieurs!études!ont!montré!que!des!lymphocytes!T!(LT)!γδ!humains!aussi!appelées!LT! non! conventionnels,! sont! impliqués! dans! la! réponse! immune! précoce! à! l’infection! par! les! stades!érythrocytaires!Plasmodium!falciparum.!Il!existe!plusieurs!sousSpopulations!de!LT!γδ! dont!une,!caractéristique!des!primates,!exprime!un!récepteur!à!l’antigène!(TCR)!Vγ9Vδ2.!Ces! LT!Vγ9Vδ2!sont!activés!par!des!phosphoantigènes!(PhSAg)!qui,!a!priori,!ne!nécessitent!pas! d’être!apprêtées!sur!les!molécules!de!CMH!I!et!II!classiques,!contrairement!aux!LTαβ.!Un!des! PhSAg! prototypiques,! le! HMBPP! (1ShydroxyS2SmethylS2Sbutenyl! 4! diphosphate)! est! produit! par!la!voie!de!synthèse!des!isoprénoïdes!dite!«!non!mévalonate!»ou!DOXSP&et!utilisée!par!de! nombreux! microSorganismes,! y! compris! les! parasites! Apicomplexa! tels! que! P.! falciparum! (pour!revue!:!Morita!et!al.!2007).!Il!a!été!montré!que!les!cellules!T!Vγ9Vδ2!sont!activées!par! des!PhSAg!produits&par&des!globules!rouges!parasités!(GRP)!au!stade!schizonte!(Goerlich!et!al! 1991;! Jones! et! al,! 1996;.! Costa! et! al! 2011).! L’activation! des! LT! Vγ9Vδ2! se! traduit! par! une! sécrétion! de! cytokines! (Goodier! et! al.! 1995;! Pichyangkul! et! al.! 1997;! Scragg! et! al.! 1999;! D’Ombrain! et! al.! 2007),! une! prolifération! (Behr! &! Dubois! 1992)! et! aussi! par! une! dégranulation!qui!inhibe!la!forme!!extracellulaire!du!parasite,!appelée!mérozoïte!(Farouk!et! al.!2004;!Costa!et!al.!2011),!Des!études!antérieures!ont!démontré!une!activation!des!cellules! T! Vγ9Vδ2! par! des! extraits! de! schizontes! et! par! des! lysats! de! GRP! (Behr! &! Dubois! 1992;! Goodier!et!al.!1995;!Behr!et!al.!1996;!Waterfall!et!al.!1998;!P.!Jagannathan!et!al.!2014),!ainsi! que!par!des!surnageants!de!culture!(Goodier!et!al.!1995;!Jones!et!al.!1996;!Lindberg!et!al.! 2013)! et/ou! par! la! coSculture! des! LT! Vγ9Vδ2! avec! des! GRP! intacts! (Waterfall! et! al.! 1998;! Costa!et!al.!2011;!Ribot!et!al.!2012).!Ainsi,!les!LT!Vγ9Vδ2!semblent!jouer!un!rôle!effecteur! important! à! étudier! et! à! exploiter.! En! effet! de! nombreux! aspects! fonctionnels! restent! ambigus,! en! particulier! les! mécanismes! conduisant! à! leur! activation! par! les! stades! érythrocytaires!du!parasite!et!leur!impact!sur!la!réponse!immunitaire!adaptative.!
Premièrement,!les!mécanismes!de!production,!de!libération!et!de!présentation!des!PhS Ag!par!des!!GRP!aux!LT!Vγ9Vδ2!sont!peu!connus.!Il!a!été!récemment!montré!que!les!PhSAg! produits! par! les! cellules! tumorales! sont! présentés! au! TCR! Vγ9Vδ2! via! la! Butyrophiline3A1! (BTN3A1)!(Harly!et!al.!2012;!Sandstrom!et!al.!2014;!Vavassori!et!al.!2013;!Wang!et!al.!2013).! Par!contre,!l’expression!de!BTN3A1!à!la!surface!des!globules!rouges!n’avait!pas!encore!été! rapportée.!Récemment,!des!vésicules!de!type!exosomes!dérivés!de!GRP!ont!été!impliquées! dans!la!communication!interSparasitaire!et!dans!l’activation!du!système!immunitaire!(RegevS Rudzki!et!al.!2013;!Mantel!et!al.!2013).!On!ne!sait!pas!si!les!PhSAg!sont!présentés!via!des! vésicules!«!exosomes!like!».!
! !
5! Deuxièmement! la! stimulation! des! LT! Vγ9Vδ2! in! vitro! par! les! PhSAg! induit! à! la! fois! l’expression! de! molécules! présentatrices! et! coSstimulatrices! associées! à! la! fonction! des! cellules! présentatrices! d’antigènes! (CPA).! Ces! LT! γδSCPA! sont! capables! de! capturer,! de! processer!et!de!présenter!des!antigènes!bactériens!et!viraux!à!des!LT!αβ!naïfs!CD4+!et!CD8+! (Brandes!et!al.!2005;!Brandes!et!al.!2009;!Landmeier!et!al.!2009;!Altvater!et!al.!2012;!Khan,! Curbishley,!et!al.!2014).!Cependant!l’implication!in!vivo!des!LT!γδSCPA!en!réponse!à!un!agent! infectieux!n’a!pas!encore!été!démontrée.!Comme!il!a!été!montré!que!les!PhSAg!produits!par! les!GRP!sont!capables!d’activer!les!LT!Vγ9Vδ2!T!il!est!possible!que!les!LT!Vγ9Vδ2!acquièrent! un!phénotype!et!une!fonction!de!CPA!dans!le!contexte!de!l’infection!à!P.!falciparum.! La!possibilité!d’une!fonction!CPA!des!LT!Vγ9Vδ2!au!cours!de!l’infection!par!P.!falciparum! est! d’autant! plus! intéressante! que! les! parasites! utilisent! de! nombreuses! stratégies! afin! d’inhiber! et! d’échapper! aux! réponses! immunitaires! de! l’hôte,! contribuant! à! l’acquisition! lente!d’une!immunité!instable!et!non!stérile!dans!les!régions!où!la!malaria!est!endémique.! Une!façon!particulière!de!faire!cela!est!d’inhiber!la!maturation!des!CPA!professionnelles,!les! cellules!dendritiques!(CD),!diminuant/empêchant!ainsi!l’activation!des!LT!αβ!CD4+!and!CD8+,! et!l’aide!subséquente!des!lymphocytes!B!(Urban!et!al.!1999;!Millington!et!al.!2006;!Elliott!et! al.!2007;!Bettiol!et!al.!2010;!PinzonSCharry!et!al.!2013).!Les!LT!Vγ9Vδ2SCPA!pourraient!ainsi! représenter!un!mécanisme!pour!contourner!l’inhibition!des!CD,!et!pourraient!être!utilisés!en! thérapeutique.! L’objet!de!cette!thèse!a!été!de!déterminer!le!mécanisme!d’activation!des!LT!Vγ9Vδ2!par! des!GRP!intacts!et!d’étudier!l’impact!de!cette!activation!sur!le!rôle!potentiel!de!CPA!des!LT! Vγ9Vδ2!au!cours!de!l’infection!par!P.!falciparum.!Ce!travail!a!donné!lieu!à!deux!publications,! résumées!ciSdessous.!
Résultats:&
Guenot,!M.*,!Loizon,!S.*,!Howard,&J.,!Costa,!G.,!Baker,!D.A.,!Mohabeer,!S.Y.,!TroyeS
Blomberg,!M.,!Moreau,!JSF.,!DéchanetS!Merville,!J.,!MercereauSPuijalon,!O.,!MamaniS
Matsuda,! M.
!
and! Behr,! C.! Phosphoantigen& burst& upon& Plasmodium* falciparum&
schizont& rupture& can& distantly& activate& Vgamma9Vdelta2& T–cells.& Infect.! Immun.,!
2015!Oct;83(10):3816S24.!
*ces!auteurs!ont!contribué!de!façon!équivalente!à!ce!travail! Dans!cet!article!nous!montrons!que!:!!
A. Les! globules! rouges! infectés! par! Plasmodium! falciparum! (GRP)! n’expriment! pas! BTN3A1!et!activent!les!LT!Vγ9Vδ2en!absence!de!contact.!!
Un!anticorps!antiSBTN3A!a!été!utilisé!pour!rechercher!l’expression!de!BTN3A!sur!les!GRP.! L’utilisation!d’inserts!de!type!transwell!empêchant!le!contact!entre!lymphocytes!et!GRP!n’a! pas!modifié!l’activation!des!LT!Vγ9Vδ2!par!les!GRP.!
B. Des! molécules! libérées! par! les! GRP! dans! le! surnageant! activent! les! LT! Vγ9Vδ2! de! façon!dépendante!du!TCR!et!ont!les!caractéristiques!de!phosphoantigènes.!!
Le! surnageant! (SN)! de! culture! de! GRP! est! capable! d’activer! les! LT! Vγ9Vδ2.! Un! traitement! des!SN!par!l’Appyrase!bloque!l’activité!de!ces!SN,!tout!comme!l’incubation!des!LT!Vγ9Vδ2! avec!un!anticorps!bloquant!le!TCR.!
C. Les! molécules! activatrices! libérées! par! les! GRP! sont! des! intermédiaires! de! la! voie! DOXSP.!!
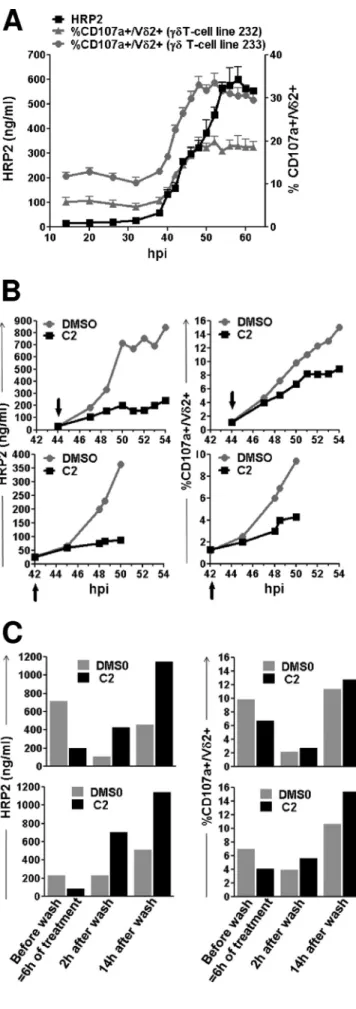
! ! 6! Le!traitement!des!GRP!par!de!la!Fosmidomycine!bloque!la!voie!DOXSP.!Les!SN!de!culture!des! GRP!ainsi!traités!ne!sont!pas!capables!d’activer!les!LT!!Vγ9Vδ2.! D. Les!molécules!stimulatrices!des!LT!Vγ9Vδ2!sont!libérées!dans!le!SN!de!culture!lors!de! la!rupture!des!schizontes,!sous!forme!libre!et!non!liée!à!des!exosomes.!! Des!GRP!sont!mis!en!culture!au!stade!40S44!heures!jusqu’à!la!rupture!et!le!SN!de!culture! correspondant!est!collecté!à!intervalles!réguliers.!La!rupture!est!monitorée!par!le!dosage!de! la!protéine!parasitaire!HRP2!dans!le!SN!et!la!présence!de!phosphoantigènes,!par!la!mesure! de!l’activation!des!LT!Vγ9Vδ2!par!les!SN.!La!cinétique!de!libération!de!phosphoantigènes!est! concomitante! avec! celle! de! la! rupture.! Le! blocage! de! celleSci! inhibe! la! libération! de! phosphoantigènes!dans!le!SN.!L’ultracentrifugation!des!SN!de!culture,!qui!élimine!les!microS vésicules,!n’empêche!pas!l’activation!des!LT!Vγ9Vδ2!par!les!SN.!
E. La! bioactivité! des! phosphoantigènes! de! P.! falciparum! est! estimée! à! 0,003S0,006! fg/GRP! et! quelques! milliers! de! GRP! sont! suffisants! pour! activer! des! LT! Vγ9Vδ2! in! vitro.!!
La! sensibilité! des! LT! Vγ9Vδ2! est! estimée! par! comparaison! de! leur! activation! doseS dépendante!par!le!HMBPP!et!par!du!SN!de!GRP.!!
!
Au! vu! de! l’ensemble! de! ces! résultats! nous! proposons! un! modèle! d’activation! des! LT! Vγ9Vδ2! dans! lequel! les! phosphoantigènes! de! la! voie! DOXSP! sont! libérés! dans! le! milieu! environnant!lors!de!la!rupture!du!parasite.!Ils!sont!alors!capables!d’activer!à!distance!les!LT! Vγ9Vδ2!via!BTN3A!exprimée!sur!les!cellules!voisines.!!
Par! la! suite! nous! nous! sommes! intéressés! aux! conséquences! de! l’activation! des! LT! Vγ9Vδ2!par!les!GRP!et!notamment!à!l’éventuelle!fonction!CPA!de!ces!lymphocytes.!!
Howard,& J.,!Loizon,!S.,!Duluc,!D.,!Pitard,!V.,!Khan,!M.W.A.,!Moser,!B.,!Mechain,!M.,!
Duvignaud,!A.,!Malvy.!D.,!TroyeSBlomberg,!M.,!Moreau,!JSF.,!DéchanetS!Merville,!J.,!
MercereauSPuijalon,!O.,!MamaniSMatsuda,!M.!and!Behr,!C.&The&antigen&presenting&
potential&of&human&Vgamma9Vdelta2&T&cells&in&response&to&P.*falciparum&infection.&
Dans!cet!article,!nous!montrons!que!:! A. Les!LTVγ9Vδ2!d’individus!infectés!par!P.!falciparum!ont!un!phénotype!de!CPA!partiel.! La!cytométrie!de!flux!a!été!utilisée!afin!d’évaluer!l’expression!de!molécules!de!présentation! et! de! coSstimulation! à! la! surface! des! LTVγ9Vδ2! de! patients! infectés! par! P.! falciparum.! En! comparaison!aux!donneurs!sains,!les!LTVγ9Vδ2!expriment!plus!de!HLASDR!et!de!CD86.!B. Les!LTVγ9Vδ2!acquièrent!un!phénotype!comparable!aux!CPA!après!une!stimulation! par!la!forme!sanguine!de!P.!falciparum!in!vitro.!!
Les!cellules!mononucléées!du!sang!périphérique!de!donneurs!sains!ont!été!stimulées!avec! des! GRP! de! 36S44! heures! purifiés! et! cultivés! pendant! 12! jours! avec! de! l’ILS2! et! de! l’ILS15.! L’expression! de! molécules! de! présentation! et! de! coSstimulation! a! été! mesurée! par! cytométrie!de!flux!tous!les!3!jours.!
C. Les! lignées! cellulaires! T! Vγ9Vδ2! stimulées! avec! les! GRP! sont! capables! de! crossS présenter!un!antigène!dérivé!de!la!protéine!M1!à!des!LT!αβ!CD8+!spécifiques.! Les!lignées!cellulaires!T!Vγ9Vδ2!stimulées!avec!les!GRP!ont!été!chargées!avec!la!protéine!M1! du!virus!de!l’influenza!sur!la!nuit,!lavées,!et!mises!en!coSculture!avec!des!LTCD8+!spécifiques! du!peptide!M1pp58S66.!L’activation!a!été!mesurée!par!la!production!d’IFNγ!des!LT!CD8+.! !
! ! 7! ! Ces!données!nous!permettent!de!conclure!qu’il!est!physiologiquement!plausible!que! les!LTVγ9Vδ2!se!comportent!comme!des!CPA!pendant!l’infection!par!P.!falciparum,!et!que!ce! champ!nécessite!des!études!complémentaires!afin!d’examiner!entièrement!leur!importance! et!leur!utilisation!thérapeutique.!
Discussion:&
A! la! lumière! de! l’ensemble! de! ces! résultats,! nous! pouvons! proposer! un! modèle! de! réponse!des!LT!Vγ9Vδ2!à!l’infection!par!P.!falciparum!:!les!stades!schizonte!de!P.!falciparum! adhèrent!à!l’endothélium!des!microvaisseaux!sanguins,!où!ils!libèrent!des!phosphoantigènes! lors! de! leur! rupture.! Cela! initie! une! inflammation! locale,! attirant! de! nombreuses! cellules! immunocompétentes! dont! les! LT! Vγ9Vδ2! qui,! tout! comme! les! cellules! endothéliales,! expriment! BTN3A! et! peuvent! donc! présenter! aux! LT! Vγ9Vδ2! les! phosphoantigènes! libérés! par! les! GRP.! Les! LT! Vγ9Vδ2! ainsi! activés! dégranulent,! ce! qui! inhibe! le! développement! parasitaire.!Les!conditions!inflammatoires!ainsi!que!les!PhSAg!présents!dans!l’environnement,! stimulent!alors!de!façon!appropriée!les!LT!Vγ9Vδ2!qui!acquièrent!un!phénotype!CPA.!Il!a!été! montré!que!les!LT!Vγ9Vδ2!ainsi!stimulés!peuvent!exprimer!le!récepteur!CCR7,!leur!conférant! une!capacité!de!homing!vers!les!organes!lymphoïdes!secondaires.!Les!LT!Vγ9Vδ2SCPA!ayant! phagocyté!des!antigènes!de!Plasmodium!présents!dans!le!milieu!peuvent!ainsi!migrer!vers! les!ganglions!où!ils!interagissent!et!activent!les!LT!CD4+!et!CD8+.! Dans!des!expériences!à!venir,!il!sera!important!de!vérifier!l’ensemble!de!fonctions!CPA! des!LT!Vγ9Vδ2!dans!un!contexte!d’infection!à!P.!falciparum.!La!capacité!à!activer!des!LT!αβ! sera!mesurée!par!des!expériences!de!réaction!lymphocytaire!mixte,!test!de!référence!pour! la! mesure! de! la! présentation! antigénique.! L’évaluation! de! la! capacité! des! LT! Vγ9Vδ2! à! processer!et!à!présenter!des!antigènes!de!P.!falciparum!nécessite!des!LT!CD8+!spécifiques.! Pour!cela,!une!lignée!LT!CD8+!spécifique!de!l’antigène!MSPS1!(merozoite!surface!protein!1!)! de!P.!falciparum!a!été!générée!au!cours!de!cette!thèse!et!sera!utilisée!pour!tester!la!capacité! des!LT!Vγ9Vδ2!à!présenter!la!protéine!MSPS1.!
L’étape! suivante! sera! de! démontrer! in! vivo! la! contribution! des! LT! Vγ9Vδ2SCPA! à! la! réponse! immune! lors! de! l’infection! par! Plasmodium.! On! pourra! alors! s’interroger! sur! des! éventuelles!applications!vaccinales!des!LT!Vγ9Vδ2.!Il!sera!notamment!important!d’identifier! le! meilleur! stimulant,! capable! de! favoriser! la! mise! en! place! des! fonctions! CPA& plutôt! que! cytotoxiques!des!LT!Vγ9Vδ2.!Ce!stimulant,!sous!réserve!qu’il!puisse!être!inoculé!sans!danger,! pourra! alors! être! utilisé! soit! pour! aider! l’expansion! des! LT! Vγ9Vδ2,! soit! comme! adjuvant!
! ! 8! ! ! ! ! !
I!would!like!to!warmly!thank!Pr.!Lars!Hviid!and!Dr.!Matthias!Eberl!for!
taking! the! responsibility! of! being! the! reporters! for! this! thesis.! I! would!
also! like! to! sincerely! thank! Pr.! Marita! TroyeSBlomberg! and! Dr.! Odile!
MercereauSPuijalon!for!accepting!to!be!examiners,!and!Pr.!JeanSFrancois!
Moreau!for!accepting!to!be!the!president!of!the!jury!for!this!thesis.!
! !
9!
Acknowledgements&
No! thesis! student! is! an! island,! and! the! work! here! presented! would! never! have! manifested! were! it! not! for! the! talented! and! kind! people! I! have! come! to! know! here! in! Bordeaux!and!in!the!EVIMalaR!community.! First!I!wish!to!thank!Charlotte!Behr,!who!took!a!chance!on!a!young!biochemistry!BSc,!to! commence!a!PhD!in!malaria!and!immunology.!ThankSyou!for!welcoming!me!into!your!home! and!family,!and!for!mentoring!me!in!my!first!steps!into!research.!And!thankSyou!for!bringing! me!to!Bordeaux,!where!I!have!learnt!so!much,!and!met!so!many!wonderful!people.!You!were! right!–!I!am!not!the!same!Jenny!that!I!was!when!I!started!!I!am!very!sad!and!sorry!that!our! time!was!cut!short,!and!I!did!not!get!to!know!you!better.!This!thesis!is!for!you!–!you!were! (again)!right!–!I!could!do!it.!! To!Julie,!Odile,!JeanSFrancois!and!Marita!who!stepped!in!to!supervise!and!guide!in!sad! circumstances,!my!heartfelt!thanks.!ThankSyou!Julie!for!taking!on!the!extra!responsibilities,! and! for! your! supervision! and! wise! guidance,! especially! in! these! last! months.! It’s! been! a! pleasure!to!work!with!you.!ThankSyou!Odile!for!making!the!many!trips!to!Bordeaux!to!meet! with! us! and! for! your! invaluable! advices! and! direction.! ThankSyou! JeanSFrancois! for! your! discussions!and!the!impartation!of!knowledge!and!wisdoms!that!enriched!this!work.!ThankS you! Marita! for! your! support! and! encouragement,! and! welcoming! me! in! my! visit! to! Stockholm.!
‘L’Equip! Behr’! –! what! would! I! have! done! without! you!! Séverine,! Maria,! Cecile,! Julian,! Marianne,! Dorothée! –! you! have! housed! me! when! I! was! homeless,! taught! me! when! I! was! clueless,!encouraged!me!when!I!was!downhearted,!pushed!me!when!I!was!dragging!my!feet! and!helped!me!in!hundreds!of!other!little,!and!not!so!little!ways.!A!million!ThankSyous.!I’m! going!to!miss!you.!And!to!all!the!students!who!passed!through!our!group,!especially!Kavish! and!Ines!–!thankSyou!for!discussion!had,!experiments!made!together!and!patience!while!I! tried!to!explain!in!my!nascent!French…! A!tous!les!CIRIDSiens,!merci!pour!votre!chaleureux!accueil!de!cette!fille!100%!anglaise.! Merci!pour!votre!patience!avec!mes!questions!toutes!bêtes!sur!des!choses!administratives,! des! protocoles,! des! correspondances,! et! les! choses! pratiques! concernant! la! vie! francais,!! pour!les!discussions!scientifiques!bilingues,!les!fous!rires!dans!le!couloir,!des!pauses!café!au! soleil! et! tous! les! moments! sympas,! dans! le! labo! et! en! dehors,! que! nous! avons! passé! ensemble.!Et!merci!en!plus!pour!m’avoir!appris!le!Francais!!!(et!à!apprécier!l’expresso).!!
(NB.!Ce!paragraphe!a!été!corrigé!par!C.K!et!L.M.)!
Many! thanks! to! Pr.! Bernhard! Moser! and! his! lab! at! Cardiff! university! for! having! me! to! stay!and!learn!the!techniques!that!became!the!bedrock!of!‘Project2’,!particularly!Dr.!Wajid! Khan!for!your!kind!teaching!and!friendship.!Also!thanks!to!Mr!and!Mrs!Davies!for!giving!me!a! home!during!my!stay.! ThankSyou!to!the!doctors!of!the!tropical!medicine!unit!of!Bordeaux!University!Hospital,! Dr!Denis!Malvy,!Dr!Mattheiu!Mechain!and!Dr!Alexandre!Duvignaud!and!the!technicians!at! the!Hospital!Immunology!lab!for!your!collaborations!on!the!patient!studies.! ThankSyou!all!the!blood!donors!to!EFS!Acquitaine!and!the!staff!who!work!there,!without! whose!generosity!this!project!would!not!have!been!possible.! This!PhD!was!funded!by!the!EVIMalaR!network.!Many!Thanks!to!the!organisers!of!the! EVIMalaR!PhD!School!for!this!great!opportunity!and!invaluable!trainings!during!the!program.!!
! ! 10!
ThankSyou! to! the! members! of! my! thesis! advisory! committee! not! already! mentioned,! Pr.! Robert! Sauerwein! and! Dr! Olukemi! Amodu,! for! yearly! discussions! and! advices.! To! all! the! EVIMalaR! PhD! students! S! ThankSyou! for! your! friendship,! mutual! support,! and! visits.! It! has! been!brilliant!to!get!to!know!you!all!and!share!this!experience!with!you.!Look!how!far!we’ve! come!since!that!first!trip!to!Heidelberg!!
To! my! friends! in! Bordeaux,! particularly! France,! Ashley,! Melissa,! Bernard,! Lilli,! Savitha,! Bode,! Yejide,! and! Tife! S! ThankSyou! for! your! encouragement! and! care,! and! for! keeping! me! sane!and!fed,!especially!during!the!writing!of!this!manuscript.!Thanks!also!to!the!wonderful! people!at!the!Bordeaux!English!church!for!your!prayers!and!boundless!kindness.!
To!my!friends!across!the!stream!–!thankSyou!for!visits,!parcels!and!Skype,!and!for!always! being!available!when!I!was!in!town.!
Finally! to! my! family! –! thankSyou! for! or! your! constant! love! and! care,! and! seemingly!
! ! 11! ! ! ! ! ! The!first!ray!of!light!which!illumines!the!gloom,!and!converts!into!a!dazzling!brilliancy!that! obscurity!in!which!the!role!of!the!Vγ9Vδ2!T!cells!in!the!instruction!of!the!adaptive!immune! response!to!P.!falciparum!infection!would!appear!to!be!under,!is!derived!from!the!sum!work! of!4!years!study,!which!the!author!of!these!papers!feels!the!highest!pleasure!in!laying!before! her! readers,! as! a! proof! of! the! careful! attention,! indefatigable! assiduity,! and! nice! discrimination,!with!which!her!search!among!the!multifarious!documents!and!experimental! data!confided!to!her!has!been!conducted.!
! ! 12!
Table&of&Contents&
1! &Introduction&...&19!
1.1!
Malaria&...&21!
1.1.1!
P.!falciparum!lifecycle!...!23!
1.2!
Immune&responses&to&P.*falciparum&infection&...&23!
1.2.1!
PreSerythrocytic!immunity!...!25!
1.2.1.1!
Humoral!immunity!...!25!
1.2.1.2!
Cellular!immunity!...!25!
1.2.2!
Erythrocytic!immunity!...!26!
1.2.2.1!
Humoral!immunity!...!26!
1.2.2.2!
Cellular!immunity!...!29!
1.2.3!
Antigen!presenting!cell!inhibition.!...!30!
1.2.3.1!
Kupffer!cell!inhibition!(preSerythrocytic)!...!31!
1.2.3.2!
Dendritic!Cell!inhibition!(erythrocytic)!...!31!
1.2.4!
Vaccination!...!32!
1.2.4.1!
The!importance!of!antigen!presentation!...!32!
1.2.4.2!
Spotlight!on!MSP1!...!32!
1.3!
Human&Vγ9Vδ2&T&cells&...&33!
1.3.1!
yδ!T!cell!development!...!36!
1.3.2!
Vγ9Vδ2!TCR!and!ligand!recognition!...!36!
1.3.2.1!
PSAg!and!the!DOXSP!isoprenoid!biosynthesis!pathway!...!36!
1.3.2.2!
PSAg!presentation!S!Butyrophilin!...!38!
1.3.2.3!
Modulating!Vγ9Vδ2!T!cell!activation:!CoSreceptors!and!their!ligands!...!41!
1.3.2.3.1!
Adhesion!partner!molecules!...!41!
1.3.2.3.2!
Natural!Killer!receptors!(NKR)!...!41!
1.3.2.3.3!
TollSlike!receptors!(TLR)!...!41!
1.3.2.3.4!
Fc!receptor!...!41!
1.3.3!
Vγ9Vδ2!T!cell!role!antiSinfection!...!41!
1.3.3.1!
Expansion!and!differentiation!...!43!
1.3.3.2!
Effector!functions!...!43!
1.3.3.2.1!
Cytokine!secretion!...!43!
1.3.3.2.2!
Cytotoxicity!...!43!
1.3.3.2.3!
T!Regulatory!functions!...!44!
1.3.3.2.4!
Interactions!with!other!immune!cells!...!44!
1.4!
Vγ9Vδ2&T&cells&as&professional&antigen&presenting&cells&...&45!
1.4.1!
What!defines!a!professional!antigen!presenting!cell?!...!45!
1.4.2!
A!precedent!in!other!mammals!S!APCSlike!function!of!γδ!T!cells!in!cows,!pigs! and!mice.!...!50!
1.4.3!
A!suggestive!Chemokine!receptor!profile!S!evidence!of!Vγ9Vδ2!!T!cells!being! in!the!right!place!at!the!right!time.!...!51!
1.4.4!
Ability!for!effective!αβ!T!cell!activation.!...!51!
1.4.4.1!
‘DCSlike’!surface!molecule!expression.!...!51!
1.4.4.2!
Ability!to!present!to!and!effectively!activate!naïve!CD4!T!cells.!...!52!
1.4.4.3!
Ability!to!present!to!and!effectively!activate!naïve!CD8!T!cells.!...!53!
1.4.4.4!
Gold!Standard!Ag!presentation!test:!MixedSlymphocyte!reaction!(MLR).!...!53!
1.4.5!
Ability!to!acquire!and!process!Ag.!...!54!
1.4.5.1!
Acquisition!and!processing!of!endogenous!Ag!for!class!I!presentation.!...!54!
1.4.5.2!
Acquisition!and!processing!of!exogenous!Ag!for!class!II!presentation.!...!54!
1.4.5.3!
Processing!of!exogenous!Ag!for!class!I!presentation!/!crossS!presentation.!...!55!
1.4.6!
How!when!and!where!might!this!happen!in!vivo?!...!56!
1.4.7!
Potential!for!Vγ9Vδ2!TSAPCs!in!immunotherapy!...!58!
! !
13! 1.5
!
Vγ9Vδ2+&T&cells&in&the&immune&response&to&P.&falciparum&infection&...&59!
1.5.1
!
Vγ9Vδ2!T!cell!activation!in!response!to!P.!falciparum!exposure!...!59!
1.5.1.1
!
Early! Evidence! that! Vγ9Vδ2! T! cells! play! a! role! in! the! immune! response! to! P.! falciparum!S!Activation!of!Vγ9Vδ2!T!cells!in!primary!infections!...!59!
1.5.1.2!
Mechanism!of!activation!...!60!
1.5.1.3!
Difference!between!primary!and!secondary!immune!responses!...!60!
1.5.2!
How!does!Vγ9Vδ2!T!cell!activation!affect!disease!outcome?!...!61!
1.5.2.1!
Lessons!from!mouse!studies.!...!61!
1.5.2.2!
Vγ9Vδ2!T!cells!release!inflammatory!cytokines!...!64!
1.5.2.2.1!
Cytokine!production!is!associated!with!protection:!...!64!
1.5.2.2.2!
Cytokine!production!is!associated!with!pathogenesis:!...!65!
1.5.2.3!
Direct!antiSparasitic!mechanism!in!vitro!...!65!
1.5.3!
Discussion!...!65!
2! &Results&...&67!
2.1!
Project&Rationale&...&69!
2.2!
Project&1.&Vγ9Vδ2&T&cell&activation&by&intact&P.&falciparum&infected&red&blood& cells.&...&70!
2.3!
Project& 2.& Vγ9Vδ2& T& cell& APC& potential& in& response& to& P.* falciparum& iRBC& stimulation.&...&83!
2.3.1!
Project!2.!Additional!Work!...!102!
2.3.1.1
!
Can! Vγ9Vδ2! T! cells! activate! αβ! T! cells! in! the! context! of! a! mixedSlymphocyte! reaction!(MLR)?!...!102!
2.3.1.1.1!
Methods!...!102!
2.3.1.1.2!
Results!and!discussion!...!104!
2.3.1.2!
Can!Vγ9Vδ2!TSAPCs!process!and!present!a!P.!falciparum!blood!stage!Ag?!...!104!
2.3.1.2.1!
Methods!...!105!
2.3.1.2.2!
Results!and!discussion!...!105!
2.3.1.3!
Are!Vγ9Vδ2!TSAPCs!susceptible!to!adhesionSmediated!iRBC!inhibition?!...!106!
2.3.1.3.1!
Methods!...!106!
2.3.2!
Project!2!Conclusions!...!108!
3! &Discussion&...&109!
3.1!
The& Cytotoxic& and& APC]like& functions& of& Vγ9Vδ2& T& cells& in& response& to& P.* falciparum&activation:&...&111!
3.2!
Could& Vγ9Vδ2& T& cells& compensate& for& DC& inhibition& during& P.* falciparum& infection?&...&113!
3.3!
Cytotoxic&and&antigen&presenting&Vγ9Vδ2&T&cell&activation&and&impact&during&P.* falciparum&infection.&...&114!
3.4!
Potential&therapeutic&applications&of&Vγ9Vδ2&T]APCs&in&P.*falciparum&infection. &...&115!
3.5!
Perspectives&...&116!
4! &References&...&119!
5! Annex:&Trainings&and&Communication&of&Results.&...&152!
! ! !! ! 14!
List&of&Illustrations&
Figures:&
Figure!1!:!Countries!with!ongoing!transmission!of!malaria!2013!(WHO!2014).!...!22!
Figure! 2! :! Countries! reporting! insecticide! resistance! since! 2010,! by! number! of! insecticide! classes!(WHO!2014).!...!22!
Figure! 3! :! Areas,! or! tiers,! of! differing! risks! of! artemisinin! resistance,! SouthSEast! Asia,! December!2014!(WHO!2014).!...!22!
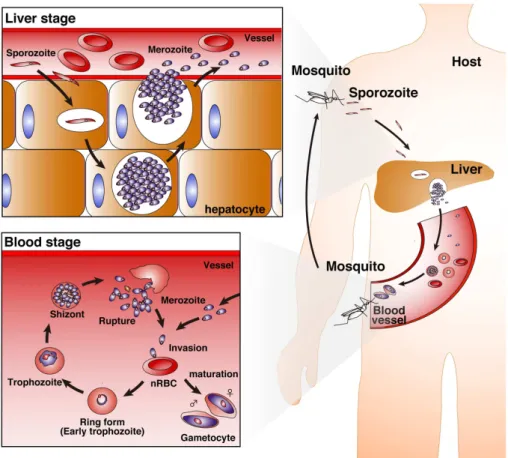
Figure!4!:!Life!cycle!of!Plasmodium!falciparum!parasites!(Inoue!et!al.!2013).!...!24!
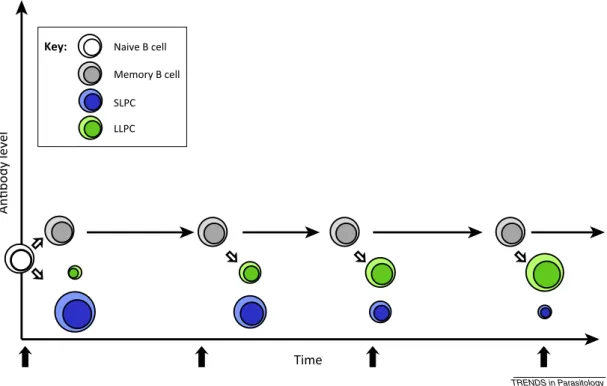
Figure!5!:!Proposed!time/exposureSdependent!changes!in!antibody!longevity!and!plasma!cell! composition!(Hviid!et!al.!2015).!...!28!
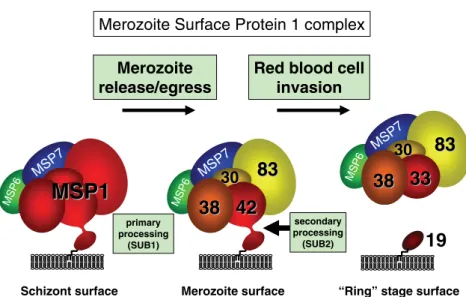
Figure!6!:!The!assembly!and!processing!of!the!MSP1!complex!(Holder!2009).!...!34!
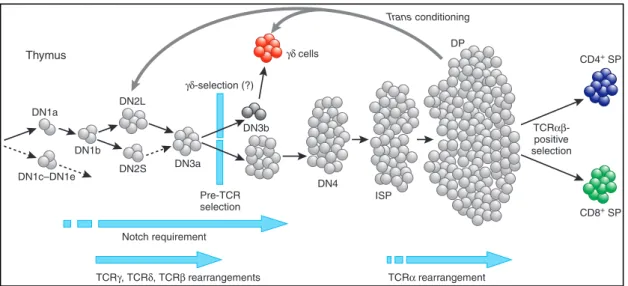
Figure!7!:!Stages!of!T!cell!differentiation!(Hayday!&!Pennington!2007).!...!35!
Figure! 8! :! Genetic! organization! and! putative! variability! of! the! γδ! TCR! adapted! from! (Konigshofer!&!Chien!2006;!Bonilla!&!Oettgen!2010).!...!35!
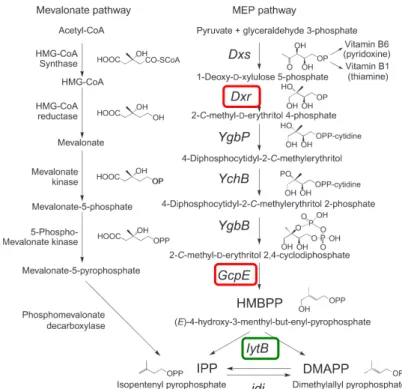
Figure!9!:!MEP!(DOXSP)!and!mevalonate!pathways!for!isoprenoid!biosynthesis.!(Morita!et!al.! 2007)!...!37!
Figure!10!:!Structure!and!activity!of!prenyl!pyrophosphate!antigens!and!analogs.!(Morita!et! al.!2007)!...!37!
Figure!11!:!Simplified!schematics!depicting!the!allosteric!model!and!the!antigenSpresenting! model!of!BNT3A1!PSAg!presentation!(Karunakaran!&!Herrmann!2014).!...!40!
Figure!12!:!γδ!TScell!receptors!(Rey!et!al.!2009).!...!40!
Figure! 13! :! APC! and! αβ! T! cell! interaction! at! the! synaptic! cleft! (Huppa! &! Davis! 2003;! Kambayashi!&!Laufer!2014).!...!46!
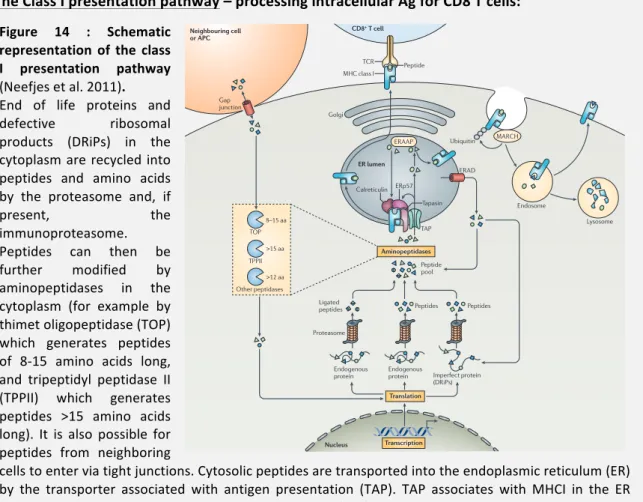
Figure! 14! :! Schematic! representation! of! the! class! I! presentation! pathway! (Neefjes! et! al.! 2011).!...!48!
Figure! 15! :! Schematic! representation! of! the! class! II! presentation! pathway! (Neefjes! et! al.! 2011)!...!49!
Figure!16!:!Schematic!representation!of!the!crossSpresentation!pathway!(Joffre!et!al.!2012). !...!49!
Figure!17:!Possible!CD4/CD8!T!cell!activation!by!iRBCSstimulated!Vγ9Vδ2!T!cell!lines!in!MLR. !...!103!
Figure!18!:!Experiment!outline!to!test!MSP1!crossSpresentation!by!Vγ9Vδ2!T!cells.!...!105!
Figure!19!:!iRBCSstimulated!Vγ9Vδ2!T!cells!express!CD54!and!transiently!express!CD36.!...!107!
! !! ! 15!
Tables:
! Table!1!:!Activating!ligands!for!γδ!T!cells!(Vantourout!&!Hayday!2013).!...!34!
Table!2!:!T!cell!memory!subsets!and!their!characteristics&(Battistini!et!al.!2005).!...!42!
Table!3!:!CoSstimulatory!molecules,!their!receptors!and!functions.!...!47!
Table!4!:!CD4!T!helper!cell!subtypes,!their!stimulus!and!characteristics!(based!on!(Geginat!et! al.!2014)).!...!47!
Table! 5! :! Summary! of! mouse! studies! outlining! γδ! T! cell! requirement! for! Plasmodium! infection!clearance.!...!62!
Table! 6! :! Summary! of! mouse! studies! outlining! γδ! T! cell! characteristics! in! the! immune! response!to!Plasmodium!infection.!...!63!
Boxes:
!Box!1!:!1,!2,!3!Go!–!naïve!αβ!T!cell!activation.!...!46
!
Box!2!:!The!intracellular!journey!to!Ag!presentation.!...!48!
! ! 16!
List&of&Abbreviations&
A&
Ab!:!Antibody! ADCC!:!AbSdependent!CellSMediated! Cytotoxicity! ADCI!:!Antibody!Dependent!Cellular! Inhibition! Ag!:!Antigen! APC!:!Antigen!Presenting!Cell! AS!:!Adjuvant!Systems!B&
BCR!:!B!Cell!Receptor! BrHPP!:!Bromohydrin!Pyrophosphate! BRSV!:!Bovine!Respiratory!Syncytial!Virus! BTN!:!Butyrophilin!C&
CCR!:!CSC!Chemokine!Receptor! cDC!!:classical!DCs! CDR!:!Complementary!Determining!Region! C/EBPα!:!CCAAT/EnhancerSBinding!Protein! α! CFSE!:!Carboxyfluorescein!Succinimidyl! Ester! ChAd63!:!ChimpanzeeSderived!Simian! Adenovirus!63! CHMI!:!Controlled!Human!Malaria! Infection! CLIP!:!Class!IISassociated!Ii! CM!:!CerebralSMalaria! CSA!:!Chondroitin!Sulphate!A! CSP!:!Circumsporozoite!Protein! CTL!:!Cytotoxic!T!Lymphocytes!! CTLA!:!Cytotoxic!T!lymphocyteSassociated! Antigen!D&
D!:!Diversity!segment! DAMP!:!Danger!Associated!Molecular! Pattern! DC!:!Dendritic!Cell! DDT!:!!dichlorodiphenyltrichloroethane! DMAPP!:!Dimethylallyl!Pyrophosphate! DN!:!Double!Negative!T!cells! DOXSP!:!1SdeoxySDSxylulose!5Sphosphate!! DP!:!Double!Positive!T!cells! DRiPs!:!Defective!Ribosomal!Products!E&
EBV!:!Epstein!Barr!Virus! ER!:!Endoplasmic!Reticulum! ERAD!:!Endoplasmic!ReticulumSAssociated! protein!degradation! ERAP!:!ERSresident!Peptidase! ERK!:!ExtracellularSsignal!Regulated!Kinase!F&
FasSL!:!Fas!ligand! Fc!:!Fragment,!Constant! FCS!:!Foetal!Calf!Serum! FDA!Food!and!Drug!Administration! FPPS!:!Farnesyl!Pyrophosphate!Synthase!G&
GAP!:!Glidosome!Associated!Protein! GMSCSF!:!GranulocyteSMacrophage! ColonySStimulating!Factor! GPA!:!Glycophorin!A! GPI&:&Glycosyl!phosphatidyl!Inositol&&H&
Hb!:!Haemoglobin! HBsAg!:!Hepatitis!B!surface!Ag! HIV!:!Human!ImmuneSdeficiency!Virus! HLASDR!:!Human!Leukocyte!Antigen!D! locusSRelated!Protein! HMBPP!:!(E)S4SHydroxyS3SmethylSbutS2S enyl!Pyrophosphate!I&
ICAM!:!Intercellular!Adhesion!Molecule!! ICOS!:!Inducible!CoSstimulator! IFNγ!:!InterferonSγ!!! ! 17! Ig!:!Immunoglobulin!! IL!:!Interleukin! ILT2!:!ImmunoglobulinSLike!Transcript!2! IPP!:!Isopentenylpyrophosphate! IRAP!:!InsulinSRegulated!Amino!Peptidase! iRBC!:!infected!Red!Blood!Cell! ISP!:!Immune!Single!Positive! ITN!:!Insecticide!Treated!Net! iTreg!:!induced!T!regulatory!cell!
J&
J!:!Iunction!segment!K&
KO:&Knock!Out!L&
LFA!:!Lymphocyte!FunctionSassociated! Antigen! Ii!:!Invariant!chain! LLPC!:!LongSLived!Plasma!Cell! LMP:!EBV!Latent!Membrane!Protein! LPS!:!Lipopolysaccharide!M&
M.!:!Mycobacterium! MAPK!:!MitogenSActivated!Protein!Kinase! MARCH1!:!MembraneSAssociated!Ring! Finger!(C3HC4)!1! MBC!:!Memory!B!cell! MCP!:!Macrophage!Chemoattractant! Protein! MEP!:!2SCSmethylSDSerythritol!4Sphosphate! MHC!:!Major!Histocompatibility!Complex! MIC!:!MHC!classSI!Related!Proteins! MIF!:!Macrophage!Migration!Inhibitory! Factor! MLR!:!Mixed!Lymphocyte!Reaction! moDCs!:!monocyte!derived!DCs! MSP!:!Merozoite!Surface!Protein!! MSP1pp!:!MSP1!peptide!pool! MVA!:!Modified!Virus!Ankara!N&
NAI!:!Naturally!Acquired!Immunity! NK!:!Natural!Killer! NKG!:!Natural!Killer!Group! NKR!:!Natural!Killer!Receptors! NK!T!cells!:!Natural!Killer!T!cells! NLR!:!NodSLike!Receptor! NO!:!Nitric!Oxide!O&
OVA!:!Ovalbumin!P&
P.!:!Plasmodium!! PSAg!!:!Phosphoantigen! PAM!:!Pregnancy!Associated!Malaria! PAMP!:!Pathogen!Associated!Molecular! Pattern! PBMC!:!Peripheral!Blood!Mononuclear!Cell! pDCs!:!plasmocytoid!DCs! PLC!:!Peptide!Loading!Complex! PMA!!:!Phorbol!12Smyristate!13Sacetate!! PPD!:!Mycobacteria!tuberculosis!Purified! Protein!Derivative! PRRs!:!Pattern!Recognition!Receptors! PS!:!Phosphatidylserine!Q&
R&
RAS!:!Radiation!Attenuated!Sporozoite! RBCs!:!Red!Blood!Cells! RTS,S!:!CSP!based!malaria!vaccine!S&
SCID:!Severe!Combined!Immunodeficiency! SEB!:!Staphylococcus!Enterotoxin!B! SLE!:!Systemic!Lupus!Erythematous!! SLPCs!:!Short!Lived!Plasma!Cells! SMA!:!Severe!Malarial!Anaemia! SN!:!Supernatant! SP!:!Single!Positive!T!cells! Spp!:!Species! SUB!:!SubtilisinSlike!protease!T&
TAP!:!Transporter!Associated!with!Antigen!! Presentation!! ! 18! Tcm!:!Central!Memory!T!cell! TCR!:!T!Cell!Receptor! Tem!:!Effector!Memory!T!cell! Temra!:!Terminally!Differentiated!Effector! Memory!T!cell! Tfh!:!Follicular!Helper!T!!cell! TGF!:!Transforming!Growth!Factor! Th!:!Helper!T!cell! TLR!:!TollSlike!Receptor!! Tnaïve!:!Naïve!T!cells! TNF!:!Tumour!Necrosis!Factor! TOP!:!Thimet!Oligopeptidase! TPPII!:!Tripeptidyl!Peptidase!II! TRAP!:!ThrombospondinSRelated!Adhesion!!!!! Protein! Treg!:!Regulatory!T!cell! TSST!:!Toxic!Shock!Syndrome!Toxin!! TT!:!single!chain!Tetanus!Toxoid!
U&
uiRBC!:!uninfected!Red!Blood!Cell! ULBP!:!UL16SBinding!Proteins!V&
V!:!Variable!segment! VSA!:!Variant!Surface!Ags!W&
X&
Y&
Z&
! ! !! !
19!
1
&
Introduction&&
! ! 20!
! !
21!
1.1 Malaria!
Malaria!is!caused!by!infection!with!the!Plasmodium!species!(spp)!parasite.!There!are!5! species! of! Plasmodium! parasite! that! infect! humans:! P.! falciparum,! P.vivax,! P.! ovale,! P.! malariae!and!P.!knowelsi.!They!are!transmitted!during!the!blood!meal!of!a!female!Anopheles! mosquito.! Malaria! is! prevalent! in! subStropical! regions,! with! the! greatest! density! in! SubS Saharan!Africa!(See!Figure!1).!In!2013!there!was!an!estimated!584,000!malaria!deaths!worldS wide:!90%!were!in!SubSSaharan!Africa,!and!78%!were!children!under!5!years!(WHO!2014).!P.! falciparum! is! the! most! prevalent! species! in! SubSSaharan! Africa! and! the! most! deadly.! Symptoms!are!characterized!by!cyclical!episodes!of!fever!paroxysms!and!include!vomiting,! join! pains! and! jaundice.! Severe! malaria! can! be! fatal! and! presents! an! array! of! severe! symptoms!including!severe!anemia,!respiratory!distress!caused!by!severe!metabolic!acidosis,! cerebralSmalaria! (CM),! multiSorgan! failure! and,! in! pregnant! women,! placental! malaria! (Murray!et!al.!2012).!! The!relationship!between!Plasmodium!parasites!and!humans!is!thought!to!be!as!ancient! as!the!evolutionary!divergence!of!humans!and!African!great!apes.!Stable!transmission!was! established!with!the!advent!of!agriculture!around!10,000!years!ago!(Carter!&!Mendis!2002).! It!is!not!surprising!therefore!that!malaria!has!had!a!significant!impact!on!both!our!history! and!our!biology.!High!prevalence!of!thalassemia,!glucoseS6Sphosphatase!deficiency!(Tishkoff! et! al.! 2001),! sickle! cell! anaemia! and! other! haemoglobinopathies! (Liddell! et! al.! 2014)! in! malaria!endemic!countries!has!been!attributed!to!genetic!mutations!conferring!protection! against! severe! malaria! (reviewed! (Wellems! et! al.! 2009;! Mangano! &! Modiano! 2014)).! Heterozygous! expression! of! haemoglobin! (Hb)! S! grants! high! protection! to! severe! disease,! but! homozygous! expression! results! in! sickle! cell! anaemia.! Haemoglobin! C! and! E! are! also! associated!with!some!protection,!but!not!pathology!(Taylor!et!al.!2012).!The!low!levels!of!P.! vivax! in! West! and! Central! Africa! can! be! explained! by! a! genetic! mutation! in! indigenous! populations! that! suppresses! Duffy! antigen! (Ag)! expression! in! the! erythrocyte! lineage! and! protects!red!blood!cells!(RBCs)!from!invasion!(Hamblin!&!Di!Rienzo!2000;!Tournamille!et!al.! 1995;!Miller!et!al.!1975).!Genes!implicated!in!autoSimmunity!are!also!associated!with!malaria! selection,! such! as! the! FGR2B! locus,! which! codes! an! inhibitory! Fc! (fragment,! constant)! receptor.! Homozygosity! for! this! gene! results! in! decreased! immune! inhibition,! which! has! been!associated!with!systemic!lupus!erythematous!(SLE),!but!also!protection!against!severe! malaria!(Willcocks!et!al.!2010).!! At!the!peak!of!malaria!spread!in!the!19th!century,!over!half!the!world's!population!was!at! risk.!Economic!and!social!development,!combined!with!an!"eradication"!campaign!involving! massive!use!of!antimalarials!(in!particular!the!recently!developed!chloroquine)!and!intense! use!of!DDT!(dichlorodiphenyltrichloroethane)!insecticides!led!to!great!reduction!of!malaria! prevalence!during!the!1960's,!and!malaria!was!eliminated!from!North!America!and!European! Mediterranean! areas.! However! levels,! particularly! in! Africa,! began! to! rise! again! from! the! 1970’s,! partly! due! to! the! development! of! chloroquineSresistant! P.! falciparum! strains! and! DDT!resistance!mosquitoes!(Carter!&!Mendis!2002;!Wellems!et!al.!2009).!
Reduction!of!malaria!has!again!been!the!focus!of!global!attention!with!its!inclusion!in!the! United!Nations!millennium!development!goals!(target!6.C!S!Have!halted!by!2015!and!begun! to! reverse! the! incidence! of! malaria! and! other! major! diseases).! Current! control! strategies! involve!the!distribution!of!insecticide!treated!nets!(ITNs);!vector!clearance!by!indoor!residual! spraying!and!larvae!control;!the!use!of!rapid!diagnostic!tests!for!better!case!management! and!blood!stage!preventative!chemoprevention,!particularly!in!pregnant!women!and!infants.! These!have!led!to!a!30%!global!reduction!in!cases!between!2000!and!2013!(WHO!2014).!
! ! 22! ! Figure&1&:&Countries&with&ongoing&transmission&of&malaria&2013&(WHO!2014).& ! ! Figure& 2& :& Countries& reporting& insecticide& resistance& since& 2010,& by& number& of& insecticide& classes&(WHO!2014).&
!
!
Figure& 3& :& Areas,& or& tiers,& of& differing& risks& of& artemisinin& resistance,& South]East& Asia,& December&2014&(WHO!2014).&
2|WORLD MALARIA REPORT 2014
The World malaria report 2014 summarizes the status of global efforts to control and eliminate malaria. The report is produced every year by the WHO Global Malaria Programme, with the help of WHO regional and country offices, ministries of health in endemic countries, and a broad range of other partners. Data for this year’s report were assembled from 97 countries and territories with ongoing malaria transmission, and an additional six countries that are working to prevent reintroduction.
This section outlines the public health burden posed by malaria, the strategies that can be used to reduce that burden, and the goals, targets and indicators that have been set for 2015. The report then reviews global progress towards the goals and targets in relation to funding (Section 2), intervention coverage (Sections 3–7), and malaria cases and deaths (Section 8). The review is followed by Regional profiles that summarize trends in each WHO region. Country profiles are provided both for countries with ongoing malaria transmission and for those recently achieving zero local cases. Finally, annexes provide sources of data, details of the methodology used in the analysis, and tables containing country and regional data.
1.1 The public health challenge posed by malaria
Malaria transmission occurs in all six WHO regions. Globally, an estimated 3.3 billion people in 97 countries and territories are at risk of being infected with malaria and developing disease (Figure 1.1), and 1.2 billion are at high risk (>1 in 1000 chance of getting malaria in a year). According to the latest estimates, 198 million cases of malaria occurred globally in 2013 (uncertainty range 124–283 million) and the disease led to 584 000 deaths (uncertainty range 367 000–755 000), representing a decrease in malaria case incidence and mortality rates of 30% and 47% since 2000, respectively. The burden is heaviest in the WHO African Region, where an estimated 90% of all malaria deaths occur, and in children aged under 5 years, who account for 78% of all deaths.
DATA WERE ASSEMBLED FROM 97 COUNTRIES AND TERRITORIES WITH ONGOING MALARIA TRANSMISSION, AND AN ADDITIONAL SIX COUNTRIES WORKING TO PREVENT REINTRODUCTION.
1. INTRODUCTION
Figure 1.1 Countries with ongoing transmission of malaria, 2013
Source: National malaria control programme reports
Confirmed malaria cases per 1000 population
No ongoing malaria transmission Not applicable 0.1–1 0–0.1 10–50 1–10 >100 50–100
VECTOR CONTROL FOR MALARIA
WORLD MALARIA REPORT 2014|17
Insecticide resistance, especially to pyrethroids, is widespread in malaria vectors. Of the 65 countries reporting any monitoring data for 2010–2013, 53 reported resistance to at least one insecticide in one malaria vector from one collection site (Figure 3.10), and 41 countries reported resistance to two or more insecticide classes. Pyrethroid resistance was the most commonly reported (Figure 3.9); in 2013, two thirds of the countries monitoring this class reported resistance.
Recommendations of the GPIRM are slowly being implemented at the country level. In 2013, 14 of 63 countries reported the use of multiple insecticides of different classes for IRS. Seven countries that used pyrethroids for IRS in 2011 or 2012 used an alternative class with a different mode of action in 2013. In six cases this change was associated with a decline in IRS coverage, possibly due to the higher cost of the alternative. The GPIRM recommends that country programmes develop long-term plans for insecticide resistance monitoring and management that include full costing of activities. Development of these plans has only just begun: seven countries in the WHO African Region have such plans in place while there has been limited progress in development of plans in other WHO regions.
International initiatives in support of GPIRM have commenced. In 2013, WHO established a Vector Control Advisory Group to facilitate the development of new tools, approaches and technologies, and to shorten the process of bringing these to market. To improve affordability of existing and new tools, initiatives such as pooled procurements, improved global forecasting, long-term contracts and tax incentives are being explored. WHO is also supporting the development of comprehensive regional and national databases on insecticide resistance. Information will be used to inform locally appropriate vector control, guide policy for managing insecticide resistance and facilitate resource mobilization for implementation. SINCE 2010, INSECTICIDE
RESISTANCE HAS BEEN REPORTED IN 53 COUNTRIES.
Figure 3.10 Countries reporting resistance since 2010, by number of insecticide classes
Source: National malaria control programme reports, African Network for Vector Resistance, Malaria Atlas Project, President’s Malaria Initiative, published literature
Reported resistance status from standard tests No resistance detected to any classes tested Confirmed resistance (<90% mortality) to one class Confirmed resistance to two classes
Confirmed resistance to three classes Confirmed resistance to four classes No data reported
No ongoing malaria transmission Not applicable
Countries reporting resistance since 2010 , by number of insecticide classes
Not applicable
Non endemic or no ongoing malaria transmission No confirmed resistance to any classes tested
Confirmed resistance (<90% mortality) to one cla Confirmed resistance to two classes
Confirmed resistance to three classes
MALARIA TREATMENT
WORLD MALARIA REPORT 2014|29 Resistance of P. falciparum to artemisinin has been detected in five countries in the Greater Mekong subregion. Drug efficacy studies have detected resistance of P. falciparum to artemisinins in Cambodia, the Lao People’s Democratic Republic, Myanmar, Thailand and Viet Nam (Figure 6.9). Despite changes in parasite sensitivity to artemisinins in these countries, ACTs have generally remained clinically and parasitologically efficacious, provided the partner drug remains efficacious. Resistance in P. falciparum to most currently available antimalarial medicines has been detected in areas at the border of Cambodia and Thailand, complicating the choice of effective treatment for medical practitioners. P. falciparum resistance to artemisinins has not been detected outside of the Greater Mekong subregion. Reports of an increased proportion of day-3 positive patients after treatment with ACTs in TES conducted in two South American countries are still being investigated. Confirmed chloroquine resistance in P. vivax, which requires measurement of drug blood levels, has been detected in 10 countries; ACTs are now recommended for the treatment of chloroquine-resistant P. vivax. The identification of multidrug resistance, including artemisinin resistance, in the Greater Mekong subregion makes elimination of P. falciparum transmission in this region an important goal. As a follow-up to the Global plan for artemisinin resistance containment (GPARC) (36), launched in 2011, WHO released the Emergency response to artemisinin resistance in the Greater Mekong subregion: A regional framework for action 2013–2015 (ERAR) (37) in 2013. The emergency plan provides further guidance for field implementation of the containment activities outlined in the GPARC. The confirmation of independent emergence of P. falciparum resistance to artemisinins in different locations in the Greater Mekong subregion, and the development of resistance to most available antimalarial medicines at the border between Cambodia and Thailand, highlight the importance of eliminating P. falciparum transmission in the region. Such elimination is considered technically and operationally feasible, and was endorsed as a goal by the MPAC in September 2014.
RESISTANCE OF P. FALCIPARUM
TO MULTIPLE ANTIMALARIAL
MEDICINES HAS BEEN
DETECTED IN AREAS AT THE
BORDER OF CAMBODIA AND
THAILAND.
Figure 6.9 Areas, or tiers, of differing risks of artemisinin resistance, South-East Asia, December 2014
Tier 1 Tier 2 Tier 3 Tier 1 are areas where there is credible evidence of artemisinin resistance; Tier 2 are areas with significant inflows of people from Tier 1 areas, including those immediately bordering Tier 1; Tier 3 are areas with no evidence of artemisinin resistance and limited contact with Tier 1 areas PDR, People's Democratic Republic
Source: Global Malaria Programme, WHO, December 2014
! !
23! Current!methods!can!control!malaria,!but!are!unlikely!to!lead!to!eradication.!LargeSscale! use!of!insecticides!and!antiSmalarial!drugs!has!(again)!led!to!the!problem!of!resistance,!by! both! the! mosquito! vector! and! the! parasite! (See! Figure! 2! and! Figure! 3).! If! resistance! continues! to! spread! and! increase,! the! main! control! strategies! will! no! longer! be! effective.! Therefore! it! is! vital! that! research! leading! to! new! treatments! and! an! effective! vaccine! are! continued.!A!better!understanding!of!both!the!malaria!parasite!and!the!immune!response!to! infection! will! greatly! aid! these! efforts.! As! the! most! deadly! human! malaria! parasite,! P.! falciparum! is! considered! the! first! priority! for! future! interventions! and! vaccines! and! consequently!is!the!most!studied.!!
1.1.1 P.*falciparum&lifecycle&
Similar! to! other! Plasmodium! spp,! P.! falciparum! has! a! complex! lifecycle! alternating! development! in! the! mosquito! vector! and! the! human! host.! The! extracellular,! liverSinvasive! form,!the!sporozoite,!is!injected!into!the!skin!of!the!human!host!from!the!salivary!glands!of! an!anopheles!mosquito!when!it!takes!a!blood!meal.!From!here,!the!sporozoite!enters!the! blood! flow! and! travels! to! the! liver,! where! it! eventually! invades! hepatocytes.! Inside! the! hepatocyte,! the! parasite! differentiates! and! divides,! before! rupturing! from! the! cell! in! a! membraneSbound!sack!called!a!merosome.!This!is!broken!up!in!the!small!capillaries!of!the! lungs! to! release! the! extracellular,! blood! stage! form! of! the! parasite! called! merozoites.! Merozoites!then!invade!RBCs.!This!is!the!clinical!stage!of!infection!where!disease!symptoms! manifest.!Inside!the!RBCs!the!parasite!has!a!48Shour!life!cycle.!To!avoid!passage!through!the! spleen,! mature! intracellular! blood! stage! forms! (trophozoites! and! schizonts)! adhere! to! endothelial!cell!walls!or!form!cellular!aggregates!with!blood!cells!such!as!RBCs!or!platelets.! Severe!malaria!combines!a!high!parasite!biomass!with!systemic!inflammation!and!results!in! sequestration!of!large!parasite!numbers!in!specific!organs.!This!sequestration!in!the!tissues! causes! local! tissue! damage! and! inflammation,! and! is! a! hallmark! of! falciparum! malaria.! Sequestration! in! the! brain! is! thought! to! be! a! causative! factor! in! cerebral! malaria,! and! adherence!to!Chondroitin!sulphate!A!(CSA)!in!the!placenta!is!the!cause!of!placental!malaria! (Magistrado! et! al.! 2008).! For! an! inSdepth! review! of! the! role! of! sequestration! in! pathophysiology!see!(White!et!al.!2013).!A!small!fraction!of!the!newly!invaded!merozoites! engage! in! the! sexual! developmental! cycle! and! mature! into! male! and! female! intraerythrocytic!gametocytes.!If!taken!up!in!the!blood!meal!of!an!anopheles!mosquito!these! become!gametes,!mate!to!form!a!zygote!that!encysts!in!the!midgut!wall,!before!releasing! sporozoites!that!migrate!to!the!salivary!glands!ready!to!be!injected!into!another!human!host.! See!(Figure!4)!for!summary.!!
1.2 Immune&responses&to&P.*falciparum&infection&
For! over! 100! years! it! has! been! observed! that! partial! immunity! to! malaria! in! endemic! areas! is! only! acquired! after! multiple! disease! episodes! (Koch! 1900;! Trape! &! Rogier! 1996;! Marsh!&!Kinyanjui!2006).!In!endemic!settings,!immunity!is!developed!first!to!severe!malaria! (usually!before!5!years!old)!then!to!clinical!malaria!(by!10S15!years!old)!(Struik!&!Riley!2004;! Marsh! &! Kinyanjui! 2006;! Langhorne! et! al.! 2008;! Doolan! et! al.! 2009).! Acquired! immunity! appears! to! be! strainS! and! variantS! specific! and! in! endemic! areas! people! are! frequently! reS infected!by!novel!variants!with!novel!antigen!combinations.!This!complicates!the!assessment! of!sterility,!however!it!is!commonly!accepted!that!full!sterile!immunity!is!never!reached!and! low!parasitemia!with!no!clinical!symptoms!is!instead!maintained!(Tran!et!al.!2013;!Bousema! et!al.!2014).!!
! ! 24! ! ! ! Figure&4&:&Life&cycle&of&Plasmodium*falciparum*parasites&(Inoue!et!al.!2013).& Summary!of!liver!and!blood!stage!human!infection!by!P.!falciparum.!See!text!for!details.!
Inoue et al. IFN-g and gd T cells in malaria
FIGURE 1 | Life cycle of Plasmodium parasites. A bite from a
Plasmodium-infected Anopheles mosquito inoculates human hosts with
sporozoites in the dermis of skin. The sporozoites move into the bloodstream, and parasitize hepatocytes. Invaded sporozoites develop into merozoites with replication (liver-stage). Then, merozoites are released from hepatocytes.
Merozoites parasitize red blood cells (RBCs), replicate, and then rupture the RBCs, leading to new infection of normal RBCs (nRBCs; blood-stage). Merozoites have several developmental phases: the ring form (early trophozoite), trophozoite, and schizont. Furthermore, some merozoites mature into female and male gametocytes.
related to adaptive immunity. gd T cells play critical roles in protec-tive immune responses against protozoan parasites, bacteria, and viruses that are associated with various infectious diseases (26–
32). This review focuses on the protective abilities of gd T cells and IFN-g in the response against malaria infection.
IFN-g MEDIATES PROTECTIVE IMMUNITY AGAINST BLOOD-STAGE PLASMODIUM PARASITES
Mice (on a C57BL/6 or CBA background) that are genetically IFN-g-deficient or IFN-g receptor (IFN-gR)-deficient or that are treated with anti-IFN-g antibody and infected with blood-stage P. berghei are unable to control the infecting parasite (33–35). In the cases of infection with blood-stage P. chabaudi and P. yoelii parasites, genetically IFN-g-deficient or IFN-g receptor (IFN-gR)-deficient mice or anti-IFN-g antibody-treated mice on a C57BL/6 or CBA background show delayed elimination of the parasites (36–39). These experimental malaria models demonstrate that IFN-g is a key pro-inflammatory cytokine for controlling blood-stage Plasmodium parasites (Table 1). IFN-g is produced by many cell types and involved in many steps of immune responses. ab T cells, NK cells, NKT cells, and gd T cells have been shown to produce IFN-g after infection with Plasmodium parasites. The
contributions of producers of IFN-g to protective immunity against Plasmodium parasites are complicated (Figure 2). Thus, more detailed experiments using IFN-g signaling-deficient mod-els should be performed to determine the mechanism(s) under-lying the involvement of IFN-g in immune protection against Plasmodium parasites.
IFN-g PRODUCERS AND THEIR ACTIVATION IN
PLASMODIUM INFECTION
CD4+abT CELLS AND CD8+abT CELLS
Given that MHC class II-deficient mice on a C57BL/6 background are unable to control P. yoelii- and P. chabaudi-infected RBCs (iRBCs), CD4+ abT cells respond to Plasmodium-iRBCs after priming with malarial antigens in an MHC class II context on DCs (40). CD4+abT cells strongly increase their ability to pro-duce IFN-g after infection with P. berghei (32). High proportions of CD4+abT cells from P. falciparum-infected human subjects respond to iRBCs and produce IFN-g, compared to CD4+ ab T cells from naïve human subjects (4). Moreover, some studies have shown that CD4+abT cell-depleted C57BL/6 or CBA mice treated with anti-CD4 antibody are unable to control blood-stage P. chabaudi or P. berghei (32,33,41,42). These lines of evidence
! !
25! At! each! stage! during! its! life! cycle! the! parasite! faces! many! and! various! players! of! the! innate! and! adaptive! immune! responses.! At! each! stage! it! manages! to! escape! total! destruction.!Much!research!has!been!done!to!understand!this!immune!response,!and!how! the! parasite! inhibits! the! induction! of! full! immunity.! This! section! shall! outline! the! main! immune!responses!to!preSerythrocytic!and!erythrocytic!parasites,!with!a!focus!on!the!role!of! the!adaptive!response,!and!the!means!by!which!the!parasite!inhibits!or!evades!this!response.! Antigen!presenting!cell!(APC)!inhibition!will!be!discussed!in!depth,!as!relevant!to!the!current! study.!Finally!vaccination,!and!the!issues!surrounding!induced!protective!adaptive!immunity! will!be!addressed.!
1.2.1 Pre]erythrocytic&immunity&
In!naturally!acquired!immunity!(NAI)!sterile!preSerythrocytic!immunity!is!never!achieved.! This! is! likely! due! to! three! factors! –! the! low! sporozoite! inoculation! numbers! (roughly! 100/bite),!naturally!immune!regulatory!environments!of!both!the!skin!and!the!liver!(Crispe! 2009;! Good! &! Doolan! 2010;! Guilbride! et! al.! 2010;! Honda! et! al.! 2011),! and! that! only! 1! infected! hepatocyte! can! release! sufficient! merozoites! to! establish! erythrocyte! stage! infection!(Schmidt!et!al.!2010).!!1.2.1.1 Humoral&immunity&
Humoral!immunity!is!developed!to!preSerythrocytic!parasites!in!NAI!(reviewed!(Offeddu! et!al.!2012)).!There!appears!to!be!an!age!dependent!increase!in!antiSsporozoite!antibodies! (Abs)! in! endemic! regions,! which! reflects! malaria! exposure! and! correlates! with! disease! resistance! (Tapchaisri! et! al.! 1983;! Druilhe! et! al.! 1986).! In! mice! vaccinated! with! radiation! attenuated!sporozoites!(RAS),!Ab!have!been!shown!to!contribute!to!immunity!by!decreasing! sporozoite! motility,! increasing! sporozoite! clearance,! blocking! hepatocyte! invasion! and! by! AbSdependent!cellSmediated!cytotoxicity!(ADCC)!(Vanderberg!&!Frevert!2004;!Sinnis!&!Coppi! 2007;!Duffy!et!al.!2012).!!
1.2.1.2 Cellular&immunity&&
Numerous!antigens!are!expressed!by!infected!hepatocytes!(Tarun!et!al.!2008),!including! the! major! sporozoite! surface! protein,! circumsporozoite! protein! (CSP),! which! is! also! expressed! in! the! first! hours! of! intraShepatic! development! (Kappe! et! al.! 2004).! Erythrocyte! stage! Ags! are! produced! at! the! end! of! hepatic! schizogony.! Vaccines! using! radiation! or! genetically!attenuated!sporozoites!are!able!to!induce!sterile!immunity!(Hoffman!et!al.!2002;! Vaughan!et!al.!2010)!that!is!mediated!by!AgSspecific!CD4!and!CD8!T!cells!(Trieu!et!al.!2011;! Krzych! et! al.! 1995;! Doolan! et! al.! 2003).! The! size! of! the! AgSspecific! CD8! T! cell! response! correlates! with! protection! (Webster! et! al.! 2005).! Sporozoite! and! liver! stage! AgSspecific,! pluripotent,! effector! memory! T! cells! have! also! been! found! in! immunity! generated! by! prophylaxisSaccompanied!infection!(Roestenberg!et!al.!2009;!Bijker!et!al.!2013).!!!
The!functional!roles!of!αβ!T!cells!have!been!further!addressed!in!mice.!Sporozoite!AgS specific!CD8!T!cells!primed!in!skin!draining!lymph!nodes!can!mediate!killing!of!intracellular! liver!stages!(Chakravarty!et!al.!2007;!Radtke!et!al.!2015),!although!high!levels!are!required!to! clear! infection! (Schmidt! et! al.! 2010).! Imaging! studies! reveal! that! killing! of! infected! hepatocytes! involves! the! formation! of! large! clusters! of! effector! CD8! T! cells,! driven!by! AgS specific! CD8! T! cells! (Cockburn! et! al.! 2013).! The! specific! mechanism! of! intracellular! hepatocyte! stage! cytotoxicity! is! yet! to! be! established,! but! it! is! likely! to! be! mediated! by! redundant! cytotoxic! T! lymphocyte! (CTL)! functions! such! as! perforin! and! granzyme! release,! interferonSγ! (IFNγ)! triggered! nitric! oxide! (NO)! release! and! Fas! ligand! (FasSL)! binding! (reviewed! (Overstreet! et! al.! 2008)).! CD4! T! cell! clones! obtained! from! RAS! or! CSP! peptide!
! ! 26! immunised!mice!were!found!to!be!protective!against!sporozoite!challenge!when!transferred! to!naïve!mice!(Tsuji!et!al.!1990;!Rénia!et!al.!1993).!CD4!T!cells!are!primarily!involved!in!helper! functions.!They!have!been!shown!to!be!vital!for!effector!and!memory!CD8!T!cell!expansion! and! survival! (Overstreet! et! al.! 2011)! and! are! important! in! APC! licensing! (reviewed! (Crispe! 2014)).! There! is! also! evidence! for! direct! contribution! to! parasite! clearance! as! CD4! T! cell! clones!from!CSP!peptide!immunised!mice!were!able!to!clear!parasites!from!in!vitro!P.!yoellii! infected!hepatocytes!cultures!(Rénia!et!al.!1993).!!
In!humans,!the!exact!location!of!Ag!presentation!is!currently!unknown.!It!is!thought!that! priming!of!CD8!T!cells!to!sporozoites!Ag!from!the!skin!is!vital!for!liver!immunity!to!follow.! More! detailed! mechanisms! have! been! outlined! in! mouse! models! of! Plasmodium! infection! (recently!reviewed!for!CD8!T!cells!(Corradin!&!Levitskaya!2014)).!Within!hours!of!inoculation,! 10S20%!of!sporozoites!enter!the!blood!stream!and!continue!to!the!liver,!while!15S25%!enter! the!lymphatic!system!and!are!taken!to!skin!draining!lymph!nodes!where!they!are!exposed!to! the!immune!system!(Amino!et!al.!2006;!Chakravarty!et!al.!2007).!Classical!CD11c+!Dendritic! cells! (DCs)! have! been! shown! to! take! up! sporozoite! Ag! by! phagocytosis,! and! cross! present! these!to!CD8!T!cells!(Cockburn!et!al.!2011;!Jung!et!al.!2002).!It!is!possible!that!APCs!in!the! liver,! such! as! Kupffer! cells,! may! also! present! Ag! to! both! CD8! and! CD4! T! cells! (reviewed! (Crispe!2014)).!Prolonged!Ag!presentation!is!required!for!optimal!CD8!T!cell!responses,!and! sporozoite!molecules!can!be!held!in!lymph!nodes,!the!liver!and!spleen!of!infected!mice!for! months!post!inoculation!(Cockburn!et!al.!2010).!!
1.2.2 Erythrocytic&immunity&
Clinical! disease! is! caused! by! the! blood! stage! of! infection! and! appears! roughly! 3! days! after!merozoite!release!from!the!liver.!A!small!percentage!of!cases!results!in!severe!infection,! mostly! in! young! children! or! naïve! adults.! There! is! a! large! heterogeneity! in! acquisition! of! severe!disease!immunity.!Some!infants!become!immune!to!severe!disease!after!just!one!of! two!infections,!while!others!(about!1/3)!remain!very!susceptible,!indicating!that!there!are! genetic!or!environmental!factors!that!influence!the!speed!of!NAI!acquisition.!
1.2.2.1 Humoral&immunity&
Protective!immunity!to!clinical!malaria!has!long!been!known!to!rest!on!Abs.!The!work!of! Cohen!et!al!showed!that!transfer!of!immunoglobulin!(Ig)SG!from!immune!African!adults!can! decrease!parasite!numbers!and!fever!in!acutely!infected!African!children!(Cohen!et!al.!1961).! IgG!from!immune!African!adults!also!induced!a!marked!decrease!of!parasite!biomass!in!Thai! children!with!acute!P.falciparum!malaria!(Sabchareon!et!al.!1991),!indicating!that!these!Ab! transcend!differences!of!antigenic!makeup!on!different!continents.!Ab!targeting!the!asexual! blood!stages!block!merozoite!invasion!of!RBCs!(Persson!et!al.!2013)!and!infected!red!blood! cell!(iRBC)!sequestration!(Fried!et!al.!1998).!Abs!also!opsonize!merozoites!and!iRBCs,!which! targets!them!for!complementSprocesses!(Biryukov!&!Stoute!2014),!phagocytosis!(Osier!et!al.! 2014)! and! antibody! dependent! cellular! inhibition! (ADCI)! (BouharounSTayoun! 1990).! In! malaria!endemic!areas,!protective!Ab!immunity!is!only!established!after!multiple!infections.! Two!reasons!have!been!proposed!for!this;!Ag!variance!and!inhibited!memory!B!cell!(MBC)! development.!Ag! diversity! includes! antigenic! mosaicism! created! by! random! segregation! of! the! chromosomes!during!sexual!reproduction!and!allelic!polymorphism!endowed!by!repertoires! of!variant!antigens!(Scherf!et!al.!2008).!Individuals!in!endemic!areas!are!exposed!to!multiple! strains!of!parasite!that!possess!different!gene!alleles!(Takala!&!Plowe!2009),!particularly!for! surface! expressed! Ag! such! as! merozoite! surface! protein! (MSP)S1! (Jiang! et! al.! 2000;! Simpalipan!et!al.!2014).!An!example!of!allelic!polymorphism!is!the!Variant!Surface!Ags!(VSAs)!
! !
27! on! the! iRBC,! which! are! prime! targets! for! Ab! (and! T! cell)! recognition.! VSA! are! encoded! by! multiple! var! genes! (Su! et! al.! 1995)! and! the! parasite! is! able! to! change! which! is! expressed! (Roberts!et!al.!1992),!thereby!evading!established!memory!responses!and!presenting!a!new! Ag!for!the!immune!system!to!recognize!(Staalsoe!et!al.!2002).!A!‘hole!in!the!wall’!hypothesis! was!developed,!which!proposed!that!while!a!person!is!developing!humoral!immunity!they! remain! susceptible! to! infection! by! the! parasites! for! which! they! have! no! specific! Ab! (i.e.! which!they!have!not!encountered!before).!Therefore!to!mount!an!extended!repertoire!of!Ab,! multiple! exposures! are! necessary! due! to! the! diversity! of! Ag! expressed! by! parasites! circulating!in!endemic!areas!(Bull!et!al.!1998).!
Normal!Ab!responses!are!long!lasting!(Amanna!et!al.!2007),!however!this!does!not!seem! to! be! the! case! for!Plasmodium! infection! and! there! is! evidence! that! the! response! is! shortS lived.! Indeed,! in! the! absence! of! reSinfection! Ab! levels! have! been! observed! to! decline! or! disappear!3S9!months!post!infection!(Crompton!et!al.!2010;!Cavanagh!et!al.!1998).!Long!lived! Ab!protection!relies!on!establishing!populations!of!MBC!and!longSlived!plasma!cells!(LLPCs),! in!a!process!which!requires!CD4!T!cell!help!(GoodSJacobson!&!Shlomchik!2010).!It!has!been! found!that!long!lived!MBCs!are!established!in!Plasmodium!infection,!but!inefficiently!(Weiss! et!al.!2010;!Ndungu!et!al.!2012;!Wipasa!et!al.!2010;!Dorfman!et!al.!2005;!Nogaro!et!al.!2011;! Kinyanjui!et!al.!2007;!Fonjungo!et!al.!1999).!! A!possible!reason!for!delayed!memory!acquisition!is!the!expansion!of!an!atypical!B!cell! subset!that!are!CD19+CD20+CD21SCD27SCD10SFcSreceptorSlikeS4+!(Weiss!et!al.!2009;!Doolan! et! al.! 2003).! This! subset! has! also! been! found! in! human! immuneSdeficiency! virus! (HIV)! patients,!and!is!classed!as!exhausted!(Moir!et!al.!2008).!Functional!exhaustion!of!both!B!and! T!cells!was!observed!in!chronically!exposed!African!children!(Illingworth!et!al.!2013).!It!has! been!demonstrated!in!African!children!and!adults,!that!atypical!MBCs!upregulate!inhibitory! receptors!and!have!reduced!B!cell!receptor!(BCR)Smediated!signalling.!Further,!in!vitro!these! atypical!MBCs!did!not!differentiate!into!AbSsecreting!cells!or!actively!secrete!Ab,!indicating! greatly! reduced! effector! functions! (Portugal! et! al.! 2015).! However! contrasting! this,! other! studies! have! demonstrated! atypical! MBC! proliferation! and! Ab! production! after! sporozoite! immunization! of! naïve! adults! (Scholzen! et! al.! 2014;! Nahrendorf! et! al.! 2014).! In! African! immune! adults,! Muellenbeck! et! al! found! that! atypical! memory! B! cells! produced! broadly! neutralizing!antibodies!against!blood!stage!P.falciparum!parasites!and,!unlike!classical!MBCs,! showed!signs!of!active!antibody!secretion,!(though!in!this!study!Ab!secretion!was!measured! by! secretory! Ig! mRNA! expression,! not! Ig! levels! directly)! (Muellenbeck! et! al.! 2013).! More! work! is! therefore! required! to! fully! understand! the! contribution! of! this! subset! to! AbS mediated!malaria!immunity,!and!their!impact!on!long!lived!Ab!protection.! It!is!possible!that!impaired!Ab!response!longevity!is!Ag!dependent,!and!that!different!Ags! require!different!strength!of!response!to!be!protective.!For!example,!longSlived!protective! immunity!is!seen!against!VSAs.!B!cellSmediated!protection!to!pregnancy!associated!malaria! (PAM)Sassociated!VSAs!does!not!wane!once!developed!in!pregnant!women!(Ampomah!et!al.! 2014)!and!in!a!study!in!Senegal,!increased!exposure!(and!therefore!increased!probability!of! specific! var! gene! encounter)! was! associated! with! increased! prevalence! of! Ab! to! rosetteS forming! parasites! (ViganSWomas! et! al.! 2011).! Also,! in! contrast! to! what! was! found! by! (Crompton!et!al.!2010;!Cavanagh!et!al.!1998)!in!Mali!and!Sudan,!in!Thailand,!P.!falciparum! AMAS1!and!MSP1!AgSspecific!Ab!and!MBC!responses!were!found!to!be!stable!over!5!years! post! infection! (Wipasa! et! al.! 2010).! This! indicated! that! longSlived! protection! is! possible.! However,!it!is!possible!that!subsequent!infections,!even!at!low!density,!are!providing!the!Ag! stimulus!for!response!maintenance.!
A!model!has!been!proposed!where!exposure!induces!shortSlived!Ab!responses,!mediated!