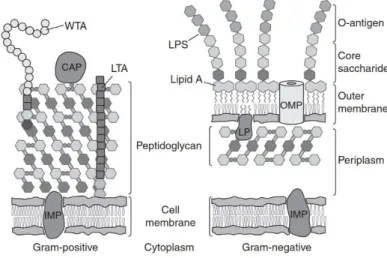
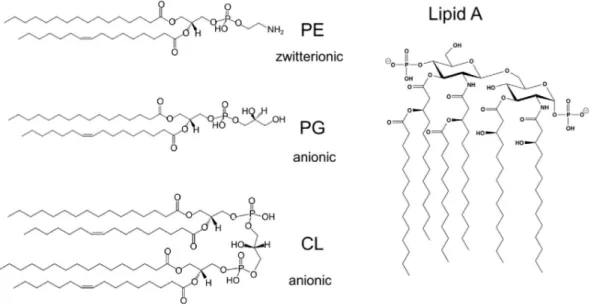
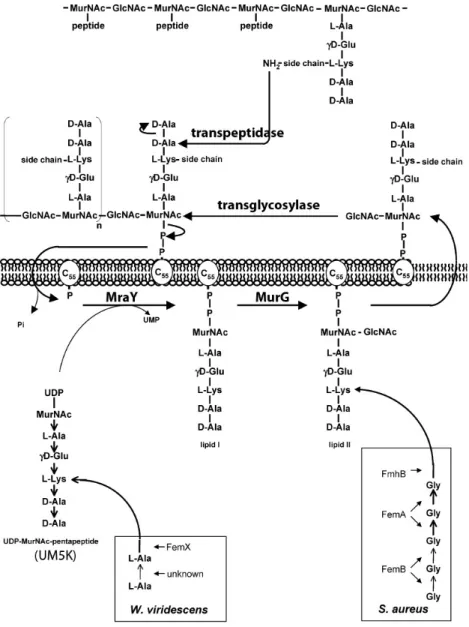
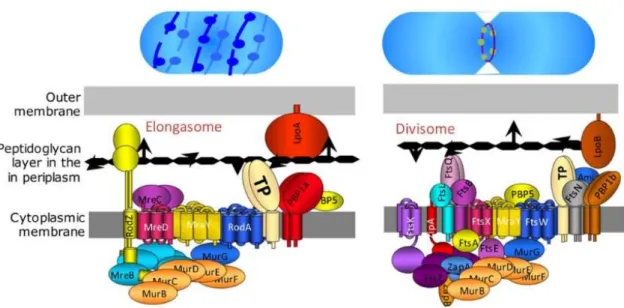
Stories from the wall: Envelope-associated functions in bacteria. Focus on cell wall synthesis, protein secretion & signal transduction machineries.
121
0
0
Texte intégral
Figure
+7
Documents relatifs