Applying the multivariate time-rescaling theorem to neural population models
Texte intégral
Figure




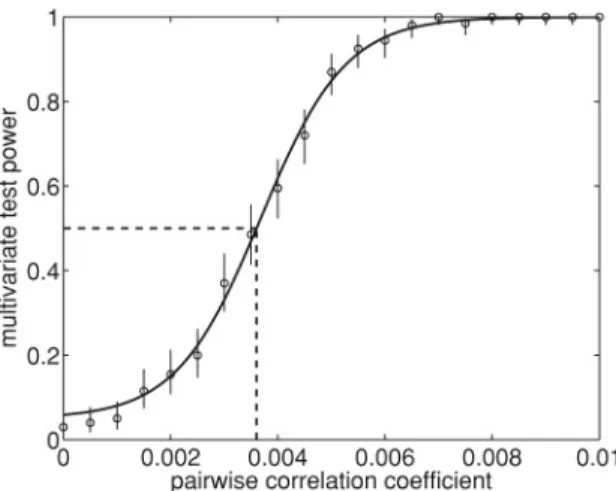

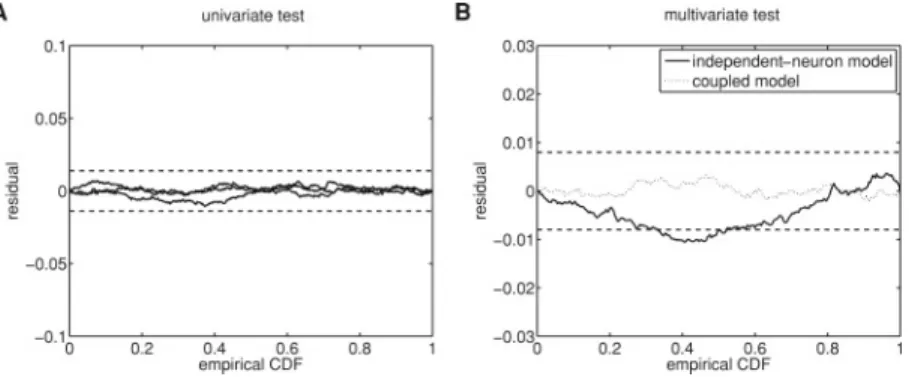

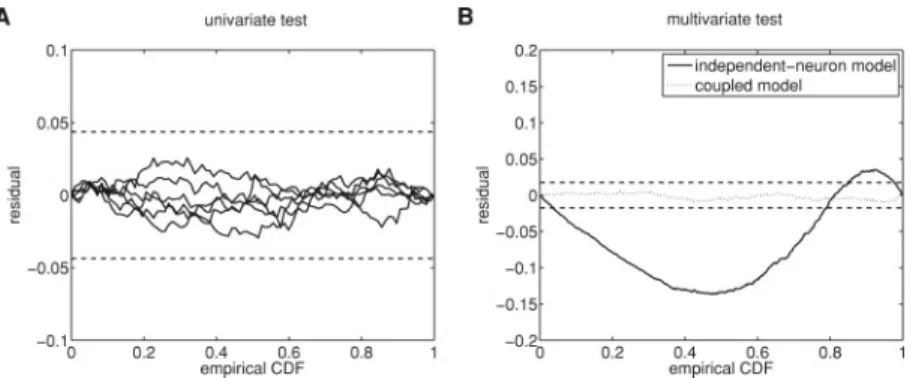
Documents relatifs
Holding meduc and f educ xed, by how much does sibs have to increase to reduce expected years of education by one year?. (A noninteger answer is acceptable
To fulfil these aims the ACT project has: defined the skills and competences of the two complementary roles of access coordinator and access manager, developed a curriculum for their
~ber 5 willkiirlichen Punkten des R.~ gibt es eine Gruppe yon 32 (involutorischen) Transformationen, welche auch in jeder anderen iiber irgend 6 besonderen
Augmented Born-Infeld equations, symmetrizable hyperbolic systems, vibrat- ing strings, singular limits, continuation principle, asymptotic expansions, energy
But crediting the play or the author with any illustrative intention is even more suspicious for it would either imply a case of prophetic or intuitive reconstitution of Beckett’s
When the transcription rates were slowed by diluting both UTP and rNTPs, there was still a slight preference towards the WRC hotspot motifs but activity also peaked at
At the conclusion of the InterNIC Directory and Database Services project, our backend database contained about 2.9 million records built from data that could be retrieved via
However, other than ICMP Source Quench messages, nothing fundamentally prevents congestion col- lapse when TCP is run over the ARPANET / MILNET and packets are being dropped at
