Functional homogenization of bumblebee communities in alpine landscapes under projected climate change
10
0
0
Texte intégral
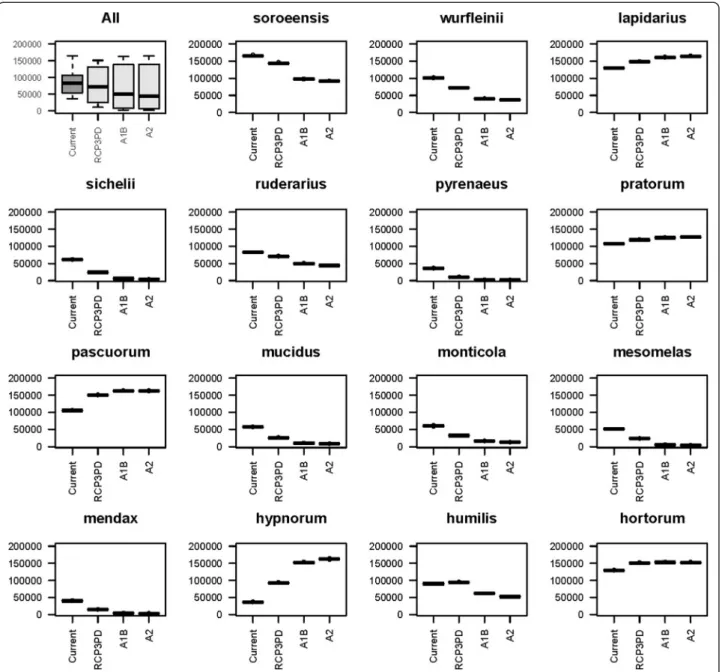
Figure



Documents relatifs