Longevity strategies in response to light in the reef coral Stylophora pistillata
Texte intégral
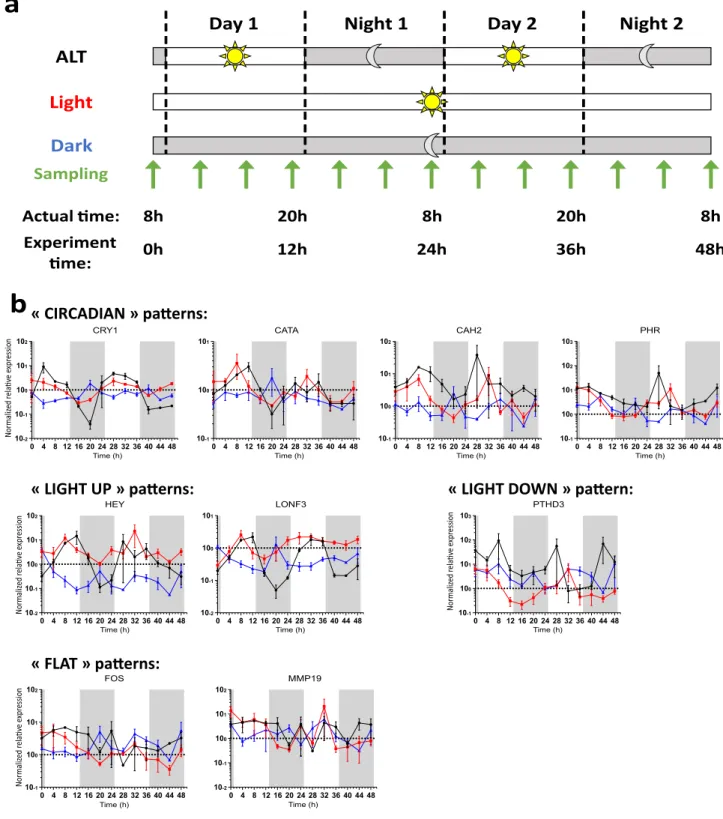
Figure

Documents relatifs
A formulation has been proposed to parameterize water inputs (wave setup and wave-induced current) generated by the breaking of oceanic swell on the reef, a method has been
In order to identify candidate genes that contribute to increased persis- tence in organs and macrophages, microarray analysis was used to compare gene expression in wild-type and
truncatula abi5 seeds, our RT-qPCR and transcriptome data showed that genes encoding enzymes of the PAO pathway were either not affected or even upregulated during maturation in
Running Head: nutrient enrichment on a coral reef Landscape-scale patterns of nutrient enrichment in a coral reef ecosystem: implications for coral to algae phase shifts... Schmitt
ici prolonge ces travaux par l’´ etude de l’´ evolution de l’att´ enuation et de la vitesse de phase en fonction de la temp´ erature et de la concentration en BSA dans un domaine
Whereas sustaining innovations improve existing technologies and products inside an existing value network and product architecture for a given market,
constructed 25 portfolios using only the ranked ex-ante dividend yield and holding the top k percent in portfolio 1 for a year, the next U percent in portfolio 2 for a year and
