The Wnt-target gene Dlk-1 is regulated by the Prmt5-associated factor Copr5 during adipogenic conversion
Texte intégral
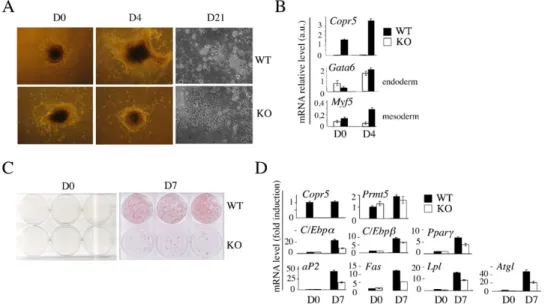
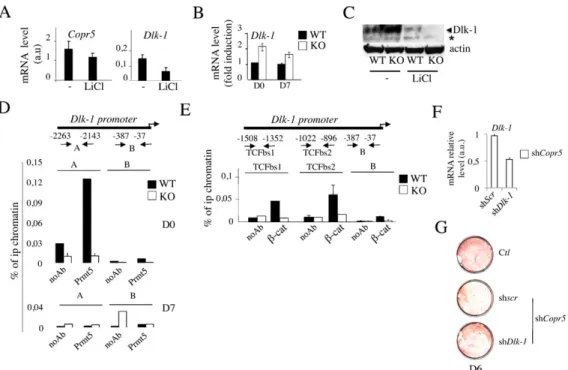
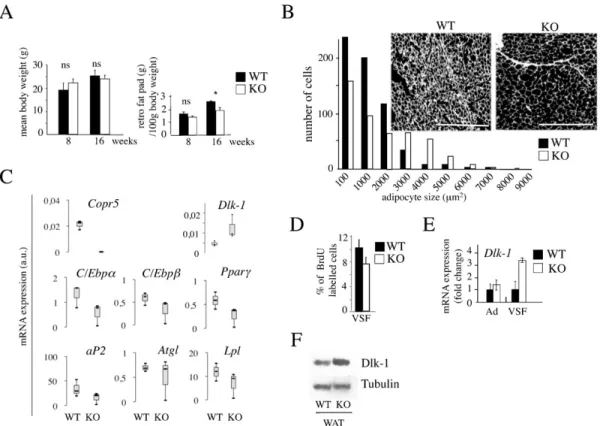
Figure
Documents relatifs
The model tracks the formation and transport in the disk of gaseous species and different solid species with their own vaporization temperatures ( see Table 1: ( i ) condensed
Infection with the protozoan Toxoplasma gondii is one of the most frequent parasitic infections worldwide. This obligate intracellular parasite was first described
Avec GOCAD, les objets sont considérés comme des ensembles de points liés entre eux afin de constituer un maillage dont la dimension est la caractéristique de l'objet. Ainsi,
La République et Canton du Jura a développé un projet de densification de l’offre en transports publics dans la région de Delémont, en collaboration avec l’Agence de Delémont de
L'amour que je ressens pour cette fille, c'est comme un roulement de tambour, un mouvement sans retour. Je t'ai choisie, parce que tu
the water dilution method, we examined the body composition of African striped
Data shown here clearly indicate that the stability of the mRNA nuclear export factor Hpr1p is controlled by the ubiq- uitin/proteasome pathway, whereas the other members of the
It can be seen that the dummy node in the center is the root node that connectes with all receptors/ligands.. The inverted triangles depict receptor
