Dual-Pitch Processing Mechanisms in Primate Auditory Cortex
14
0
0
Texte intégral
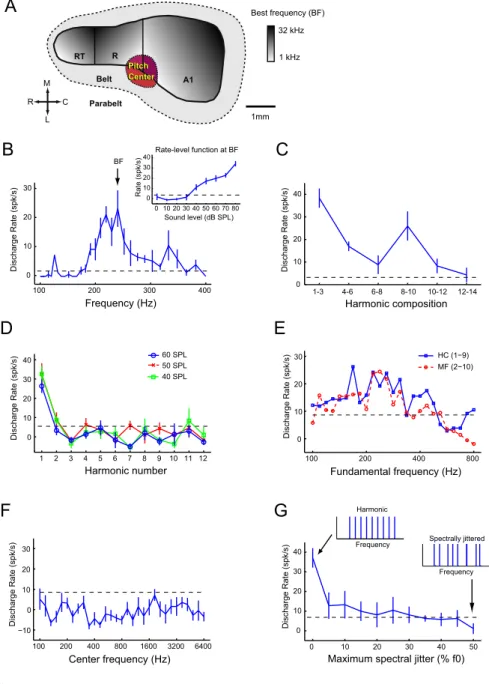
Figure

Documents relatifs