Men increase contributions to a public good when under sexual competition
Texte intégral
Figure
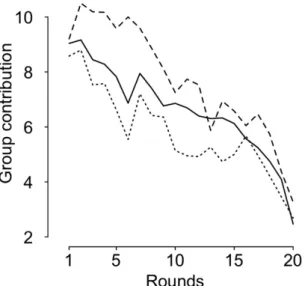
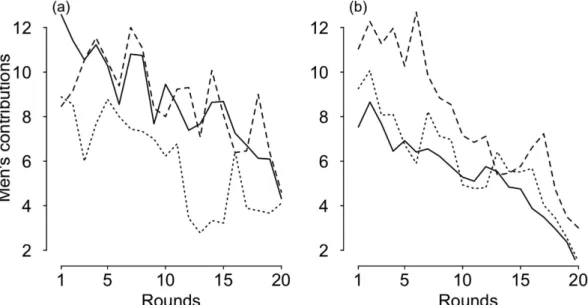
Documents relatifs
When NETED is used to create a file (that is, when it is invoked from command level with an argument which specifies the name of a file which does not already exist in the
In order to supply options for the DHCP server to send to the client, the relay agent sends an RSOO in the Relay-Forward message...
Speaker, I have to tell you at this point I can't help but recall a Public Utilities Committee meeting in 1970, when the Member for Lakeside and I sat in opposition and posed a
The maximum total new assistance in 1973 from the above programs for a municipality in the North would amount to 36 per cent of its 1972 municipal levy (i.e., 20 per cent from
In contrast to our Province, for the Maritime Provinces on average, all three components remained above the general Canadian level; Federal Government capital and
The increases range from 6 per cent to 8.7 per cent for our two largest health authorities, and I’d like to emphasize that every health authority in the province is receiving at least
Sheltered workshops are considered by the government an essential service program within a community as part of the total delivery of health and social
Budget 2013 provides a 2.7 increase in funding for health care to ensure that Manitoba families continue to have access to our existing health services, as well as services that
