BIOLOGY AND BIOCHEMISTRY OF THE MICRO-ORGANISMS IN ACTIVATED SLUDGE ]BY ROSS I. Mc KINNEY B.A. B.S.C.E.
Southern Methodist University 1948
Massachusetts Institute of Technology 1949
SUBMITTED IN PARTIAL FULFILLMNT OF THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF SCIENCE IN SANITARY ENGINEERING from the
MASSACHUSETTS INSTITUTE OF TECHNOLOGY 1951
Signature of Author:
Signature Redacted
Department of Civil and Sap )E nginxeringCertified
by
Signature Redacted
Chairman of DepartmentaSignature Redacted
on Graduate Students:I
I
BIOLOGY AND BIOCHEMISTRY OF THE MICRO-ORGANISKS IN ACTIVATED SLUDGE
By Ross E. Mc Kinney
Submitted for the degree of Doctor of Science in Sanitary Engineering in the Department of Civil and Sanitary
Engineering on May 11, 1951
The object of this study was to isolate some of the floc-producing organisms in the activated sludge process and to study their biochemical relationships in the puri-fication of sewage.
The isolation of the floc-forming organisms was the first step in this study. Two techniques were used to isolate the floc-formers, direct aeration and multiple dilutions. The direct aeration process consisted of using miniature aeration tanks to concentrate the floc-formers
for subsequent isolation by standard bacteriological pro-cedures. The multiple transfer method used the principle which had been successful in the isolation of plankton, that of repeated transfer with the desired organism
building up until a pure culture was obtained in a liquid medium. These two processes yielded several floc-forming
organisms.
After the isolation of these organisms had been made, biochemical tests were set up to identify them. The iden-tification tests used were those set forth by the Society of American Bacteriologists for the identification of bacteria. The bacteria were identified with the aid of Bergy's Manual of Determinative Bacteriology (87). The cultures identified in this study included: Zooglea ramigera. Bacillus cereus, Escherichia intermedium, Paracolobactrum aerogenoides and Norcardia actinomorpha.
Biochemical tests were set up in small aeration units to determine the organism's ability to floc and to remove B.O.D. under conditions similar to activated sludge. Both
sterile settled sewage and sterile synthetic sewage were used as substrates. The B.O.D. removals ranged from 5% to 75% in 24 hours aeration period. Higher removals were
obtained when several pure cultures were mixed than when the pure cultures were aerated alone. The addition of Paramecium to the pure culture sludge caused the B.O.D. removals to be increased. The results of these organisms checked favorably against the results obtained by other investigators who worked with pure culture sludges.
-u
-The next phase of the study concerned the respiration of the organisms. The Warburg apparatus was used to deter-mine the oxygen uptake and the carbon dioxide production of the pure cultures in suspension and in sludges. Other sew-age organisms were studied in an effort to learn of the comparative rates of oxidation. The substrates used in this study included dextrose, peptone and the combination of dextrose and peptone. Continual readings were made over a two-day period of the oxygen uptake and the carbon dioxide production. The oxygen utilization curves followed the
bacterial life cycle curve indicating that the rate of oxi-dation was a function of the number of organisms present. The Respiratory Quotient, ratio of 002:02 by volumes, was determined as a means of learning to what degree the organ-ic material was oxidized. The majority of the organisms were capable of oxidizing the organic material to carbon dioxide and water.
The final phase concerned the sludge proper, what it was made of and how it was formed. Chemical analyses were run on the sludge ash to determine its mineral components. Other analyses were made to learn of the nitrogen distri-bution in the sludge. The theories of colloidal chemistry were reviewed and combined with previous experiments by other investigators in the field of bacteriology. This information was correlated with known information in the field of sanitary engineering and with knowledge gained through experiments and a theory set forth explaining the formation and operation of activated sludge.
Professor Joseph
s.
Newell, S.E.
Secretary
ofthe Faculty
Massachusetts Institute of Technology
Cambridge,
39,Massachusetts
Dear
Sir:
Cambridge, Mass.
May 11, 1951
In partial fulfillment
ofthe requirements for
the degree of Doctor of Science in Sanitary
Engineer-ing .fromthe Massachusetts Institute
ofTechnology,
I present thia thesis entitled BIOLOGY
ANDBIOCHEMISTRY
OF THE MICRO-ORGANISMS IN ACTIVATED SLUDGE.
Respectfully submitted,
Signature redacted
-~
-ACKNOWLEDGMENTS
The author wants to take this opportunity to acknow-ledge those who have helped make this thesis possible.
Dr. M. P. Horwood - thesis supervisor
The Sanitary Engineering Department Staff - thesis advisors
Mr. Richard Engelbrecht - associate on N.I.H. Project
RG-2605
Miss Margaret Curtis assistant on the Warburg studies
This investigation was supported by a research grant from the Division of Research Grants, of the National Institutes of Health, Public Health Service
TABLE OF CONTENTS
Page I. INTRODUCTION - - - - - - - - - - - - -
-II. TBEORETICAL CONSIDERATIONS
A. Isolation - - - - - - - - - - - - -B. Respiration - - - - - - - - - - - - -III. PROCEDURE A. Method of Attack 1. Isolation - - - - - - - - - - - -2. Biochemistry - - - - - - - -Respiration - - - - - - - - - -4-- -udge - - - - - - - - - - - - -B. Apparatus 1. Isolation - - - - - - - - - - -2. Biochemistry - - - - - - - - -3. Respiration - - - - - - - - - - -4. Sludge - - - - - - - - - - - - -C. Procedure 1. Isolation - - - - - --- - - - -2. Biochemistry - - - - - - - -. Respiration - - - - - - - -- - 4.Sludge -D. Computations 1. Biochemistry - - - - - ---2. Respiration - - - - - - - - - -3. Sludge - - - - - - - - - - - - -E. Error 1. Biochemistry - - - - - - - - -2. Respiration - - - --- - - - - -3. Sludge - - - - - - - - - - - - -IV. RESULTS
A. Isolation and Identification- - -1. By Aeration Technique - - -2. By Dilution Technique - - - -3. Identification - - - - - - - -B. Biochemistry - - - - - - - - - - -C. Respiration - - - - - - - - - - - - -1. Respiration Studies With Dextrose 2. Respiration Studies With Peptone 3. Respiration Studies With Dextrose
and Peptone - - - - - - - ---D. Sludge - - - - - - - - - - - - -
-1. Mineral Analyses of Sludges- - 2. Organic Analyses of Sludges -E. Microscopic Analyses - - - - - - - -I)
5
42 43 50 50 50 51 5155
58
58
6o
64
68
69
7173
75
76
78 7981
81
86
86
88
108
109 115 123134
134
136
141
TABLE OF CONTENTS (Continued) Page V. DISCUSSION OF RESULTS A. Isolation - - - - 176 B. Biochemistry - - - -
- - - -
177 C. Respiration - - - - - - - - - - - - - 17 D. Sludge - - - - 179 E. Microscopic Analyses - - - - - - - - - 182VI. A THEORY OF ACTIVATED SLUDGE - - - - - - - 183
VII. CONCLUSIONS - - - - - - - - - - - - - --- 189
VIII. SUGGESTIONS FOR FUTURE WORK - - - - - - - 206
IX. BIBLIOGRAPHY - - - - - - - - - - - - - - - 208
X. APPENDIX A - - - - 214
XI. APPENDIX B - - - - - - - - - - - - - - - - 215
XII. APPENDIX C - - - - - - - - - - - - - - - - 219
Table No. IV II III IV VI VII VIII VIII Ix
xi
XI XII XIII XIV XV XVII
Treatment of Activated Sludge for
Isolat-ing Floc-producIsolat-ing Organisms
Culture Media Used in Isolating the Floc-producing Organisms
Cultural Characteristics of the
Floc-producing Organisms
Biochemical Study No. 1 - B.O.D. Removals Biochemical Study No. 1 - Suspended Solids
in Mixed Liquor
Biochemical Study No. 2 - The Effect of pH
on Culture B6a
Biochemical Study No. 3 - B.O.D. Removals
and Suspended Solids in Mixed Liquor
Biochemical Study No. 3A - B.O.D. Ronmovals Biochemical Study No. 3A - Suspended Solids
in Mixed Liquor
Biochemical Study No. 4 - B.O.D. Removals and Suspended Solids in Mixed Liquor
Biochemical Study No. 4A - B.O.D. Removals and Suspended Solids in Mixed Liquor
Biochemical Study No. 5 - B.O.D., B.O.D. Removed and Per cent B.O.D. Removed
Biochemical Study No. 5 - Suspended Solids in Mixed Liquor, Settleable Solids and ppm B.O.D. Removed/ppm S.S. in Mixed Liquor
Biochemical Study No. 5 - 48 Hour Total
Bacteria Count/Ml. Mixed Liquor and pH
Biochemical Study No. 6 - B.O.D., B.O.D. Removed and Per cent B.O.D. Removed
Biochemical Study No. 6 - Suspended Solids in Mixed Liquor, Settleable Solids and ppm
Page 82
85
87
89
90
92
9293
93
95
97
98
99
100 104 INDEX OP TABLES TitleITDEX OF TABLES (Oontinued)
Table No. Title Page
XVII Biochemical Study No. 6 - 48 Hour Total Bacteria Count/Ml. Mixed Liquor, pH and
Total Plate Count-Yost Probable Number 106 XVIII Respiration Study - Oxygen Utilization 219 XIX Respiration Study - Oxygen Utilization
-Dextrose 220
XX Respiration Study - Oxygen Utilization
-Dextrose 222
XXI Respiration Study - Oxygen Utilization
-Peptone 224
XXII Respiration Stu'dy - Oxygen Utilization
-Peptone 226
XXIII Respiration Study - Oxygen Utilization
-Dextrose Peptone 228
XXIV Respiration Study - Oxygen Utilization
-Dextrose Peptone 230
XXV Respiration Study - Oxygen Utilization
-Dextrose Peptone 232
XXVI Respiration Study - Carbon Dioxide
Production 234
XXVII Respiration Study - Carbon Dioxide
Production - Dextrose 235
XXVIII Respiration Study - Carbon Dioxide
Production - Dextrose 237
XXIX Respiration Study - Carbon Dioxide
Production - Peptone 239
XXX Respiration Study - Carbon Dioxide
Production - Peptone 241
XXXI Respiration Study - Carbon Dioxide
Production - Dextrose Peptone 243
XXXII Respiration Study - Carbon Dioxide
I7\rDE:X OF TABLES (Continued)
Table No. Title P
XXXIII Respiration Study - Carbon Dioxide
Production - Dextrose Peptone 247
XXXIV Respiration Study - Respiratory Quotient 249 XXXV Respiration Study - Respiratory
Quotient
-Dextrose 250
XXXVI Respiration. Study - Respiratory
Quotient
-Dextrose 252
XXXVII Respiration Study - Respiratory
Quotient
-Peptone 254
XXXVIII Respiration Study - Respiratory Quotient - 256
Peptone 256
XXXIX Respiration Study - Respiratory Quotient
-Dextrose Peptone 258
XL Respiration Study - Respiratory quotient
-Dextrose Peptone 260
XLI Respiration Study - Respiratory Quotient
-Dextrose Peptone 262
XLII Respiration Study - Suspended Solids 132
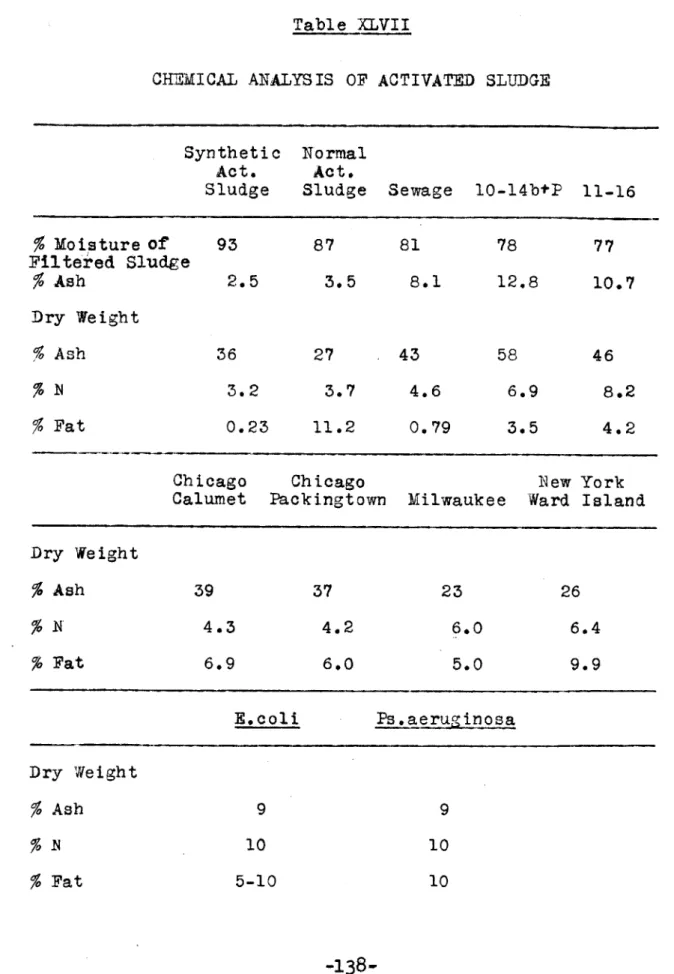
XLIII Respiration Study - Comparative Oxidation 133 XLIV Chemical Analysis of Activated Sludge Ash 135 XLV Chemical Analysis of Activated Sludge Ash 137 XLVI Chemical Analysis of Activated Sludge Ash 180 XLVII Chemical Analysis of Activated Sludge 138 XLVIII Nitrogen Distribution in Activated Sludge 1140
INDEX OF GRAPHS Figure
Number Title
A. Respiration Study - Gas Relationships -Boston Sewage
B. Respiration Study - Gas Relationship
-Normal Activated Sludge -. Dextrose
C. Respiration Study - Gas Relationship -Culture 10-14b - Dextrose
D. Respiration Study - Gas Relationship
-Culture 10-14b Sludge - Dextrose
-t.' Respiration Study - Gas Relationship -Culture 11-16 - Dextrose
F. Respiration Study - Gas Relationship
-A. aerogenes Sludge - Dextrose
G. Respiration Study - Gas Relationship -Normal Activated Sludge - Dextrose H. Respiration Study - Gas Relationship
-Synthetic Activated Sludge - Dextrose
I. Respiration Study - Gas Relationship -Culture 8-16 - Peptone
J. Respiration Study - Gas Relationship
-Culture 10-14b - Peptone
K. Respiration Study - Gas Relationship
-A. aerogenes - Peptone
L. Respiration Study - Gas Relationship
-Culture E4 - Peptone
M. Respiration Study - Gas Relationship -Culture 11-16 - Peptone
N. Respiration Study - Gas Relationship
-Normal Activated Sludge - Peptone 0. Respiration Study - Gas Relationship
-A. aerogenes Sludge - Peptone
P. Respiration Study - Gas Relationship
-Synthetic Activated Sludge - Peptone
p Page 110 140 112 112 114 114 116 116
117
117 119119
121 121 122 122Figure Number
INDEX OF GltAPHS (Continued)
Title
Respiration Study - Gas Relationship
-Culture 10-14b - Dextrose Peptone
R. Respiration Study - Gas Relationship
-Culture E4 - Dextrose Peptone
S. Respiration Study - Gas Relationship -A. aerogenes - Dextrose Peptone
T. Respiration Study - Gas Relationship
-Sewage Organisms - Dextrose Peptone
U. Respiration Study - Gas Relationship
-Culture 11-16 - Dextrose Peptone V. Re-.piration Study - Gas Relationship
-Ps. aeruginosa - Dextrose Peptone W. Respiration Study - Gas Relationship
-Ps. fluorescens - Dextrose Peptone
X. Respiration Study - Gas Relationship -Culture 8-16 Sludge - Dextrose Peptone
Y. Respiration Study - Gas Relationship -Culture 10-14b Sludge - Dextrose Peptone
Z. Res-iration Study - Gas Relationship
-Synthetic Activated Sludge - Dextrose
Peptone Page 124 124 125 125 128 128 179 129 130
130
INDEX OF ILLUSTRATIONS Figure
Number Title Page
1. Small Aeration Rack Used In Isolation Runs
54
2. Small Aeration Rack In Operation (15 Tubes)54
3. Medium Aeration Racks Used In TheBiochemical Studies In Operation 57 4. Large Aeration Rack Used In The
Biochemical Studies 57
5. The Warburg Apparatus
59
6. The Nephelometer 59
7. The Photomicrograph Equipment 61
8. Large Aeration Tubes Used To Keep Normal
Activated Sludge Alive 61
9. Normal Activated Sludge (10OX) 142 10. Normal Activated Sludge (250X) 142 11. Normal Activated Sludge (485X) 143 12. Normal Activated Sludge (970X) 143 13.. Normal Activated Sludge (970X) 144 14. Normal Activated Sludge (10X) 144 15. Normal Activated Sludge (100X) 146 16. Normal Activated Sludge (100X) 146 17. Normal Activated Sludge ( 50X) 147 18. Normal Activated Sludge (970X) 147 19. Normal Activated Sludge (970X) 148 20. Young Normal Activated Sludge (430X) 148
21. Synthetic Activated Sludge (10OX) 149 22. Synthetic Activated Sludge (430X) 149
INDEX OF ILLUSTRATIONS (Continued)
Figure
Number Title Page
23. Synthetic Activated Sludge (970X)
150
24. Culture B6a Plus Diatomaceous EarthAt pH 7.2 (100X)
25. Culture B6a Plus Diatomaceous Earth
At pH 7.3 (10OX) 151
26. Culture B6a Plus Diatomaceous Earth
At pH 7.5 (10OX) 151
27. Culture B6a Plus Diatomaceous Earth
At pH 7.8 (10OX) 152
28. Culture B6a Plus Diatomacecu3 Earth
At pH 7.2 (970X) 152
29. Culture B6a Plus Diatomaceous Earth
At pH 7.5 (970X) 153
30. Culture B6a Plus Diatomaceous Earth
At pH 7.8 (970X) 153
31. Monolayer of Culture B6a (970X) 154 32. Culture B6a in Filtered Sewage (430X) 154 33. Culture B6a in Filtered Sewage (970X) 155 34. Culture B6a in 1/10 Nutrient Broth (970X) 155 35. Early Formation of Culture B6a Floc (970X) 157
36. Culture Fla (430X)
157
37. Culture Fla (970X) 158 38. Culture Elb (101X) 158 39. Culture Elb (430X) 159 40. Culture B9 (100X) 159 41. Culture B9 (430X) 160 42. Culture B9 (970X)INDEX OF ILLUSTRATIONS (Continued)
Figure
Number Title
43. Culture B9 (970X)
44. Culture B9 in Calcium Phosphate Floc (970X) 45. Culture 10-14b in Calcium Phosphate
Floc (970X)
46. Culture 10-14b in Calcium Phosphate Floc (970X)
47. Culture 10-14b in Calcium Phosphate Floc (970X)
48. A. aerogenes in Calcium Phosphate Floc (970X)
49. Synthetic Sludge - Sewage Culture (970X)
50. Synthetic Sludge - Sewage Culture 51. Synthetic Sludge - Sewage Culture
52. Synthetic Sludge - Sewage Culture
53. Synthetic Sludge - Culture 10-14b 54. Synthetic Sludge - Culture 10-14b
55. Synthetic Sludge - Culture 10-14b
56. Synthetic Sludge -57. Synthetic Sludge -58. Synthetic Sludge -Paramecium (970X) 59. Synthetic Sludge -Paramecium (970X) Culture 10-14b (97 0X) (970X) (970X) (100X) (970X) (970X) (970X) Culture 10-14b (970X) Culture 10-14b + Culture 10-14b + 60. Synthetic Sludge - Culture 11-16 (970X) 61. Synthetic Sludge - Culture 11-16 (970X)
62. Synthetic Sludge - Culture 11-16 (970X)
Page 111 162 162
163
163
165
165
166
166
167
167
168
168
169
169
170
170 171171
WINDEX OF ILLUSTRATIONS ( Continued) Figure
Number Title Page
63. Synthetic Sludge - Culture 11-16 (970X)
173
64. Magnesia Sulfate Flocculation of Culture10-14b (970X) 173
65. Thorium Nitrate Flocculation of Culture
10-14b (970X) 174
66. Copper Sulfate Flocculation of Culture
10-14b (970X) 174
67. Calcium Chloride Flocculation of Culture
10-14b (970X) 175
68. Zinc Sulfate Flocculation of Culture
10-14b (970X) 175
69. Double Layer Representation of a Colloid
189
Al Aerobic Breakdown of Organic Matter 214BI Theory of Activated Sludge
The practical applications of the activated sludge
process have exceeded the fundamental knowledge concerning the process ever since its innovation. The understanding of the mechanisms of the activated sludge process has centered on the solution of practical problems as they arose. Many of these problems have not been satisfactorily solved, but knowledge of the fundamental aspects of the process has in-creased. Some of the problems which have been encountered in activated sludge are sludge bulking, rising sludge, poor floc formation and sludge age to mention a few. During the years research has become directed at the fundamental unit of the activated sludge process, the bacterial cell. It has been a
slow change from research on the chemical and physical aspects to the biology of the process as a whole and finally to the individual bacterial cells. This change can best be seen in
the literature concerning the activated sludge process.
Although the activated sludge process itself is not quite forty years old, the ideas which culminated in its formation go back almost twice as long. For the real beginnings of the knowledge of this process began with the advent of the science of bacteriology during the middle of the nineteenth century. It was near the beginning of this era that the first mention of the organism that was to be positively identified with activated sludge occurred. Itzigsohn (1), studying zoogleal formations, came upon a species that attracted his attention
-5-which he called Zooglea ramigera. But his findings did not go unchallenged. Others studied this new bacterium and Zopf, for one, was reported to have stated that the Zoog2a ramigera was nothing more than one phase in the life cycle of Cladothrix. Unfortunately, the means at the disposal of these early bacteriologists were not sufficient to permit positive identification either way. This resulted in
con-siderable confusion and the final discard of this organism to temporary obscurity.
During this same period two bacteriologists began to take notice of a phenomenon that was as old as the world was, the stabilization of organic matter in soil. They knew that organic matter placed on the soil was taken into the soil and changed in chemical composition to supply plants with the necessary elements. They also had listened to chemists ex-plain this phenomenon in strict chemical terms. But they were not satisfied with this explanation. In 1877 Muntz and
Schloesing (2) showed that heated soil failed to stablize sterile nitrogenous materials placed on the soil with the re-sulting ammonia, nitrite and nitrate end products. These ex-periments led them to believe that there were certain organisms in the soil which stabilized the nitrogenous materials and that the reaction was not entirely a chemical one.
With this clue and with improved modes of isolation of bacteria, Heraeus (3) in 1886 isolated 14 known species of
bacteria and about as many unknown species from water and soil.
He placed these organisms in an ammoniacal solution and after a suitable incubation period obtained nitrous acid. Thus, he had proved to himself that bacteria were responsible for nitrification. Adametz and Frank (3), also working on the same problem, introduced garden soil into ammoniacal solutions and obtained nitrification. With this result they set about to isolate the organisms responsible for the nitrification. Succesb was not entirely theirs; for, none of the cultures isolated from the garden soil would nitrify the ammoniacal solution. Frank concluded that there were not any organisms capable of nitrification; the process was purely chemical.
Further study by Warrington and Frankland (3) in 1888 tended to prove Frank's statements. They concluded that there was not any reliable evidence that any individual
species of bacteria was able to oxidize ammonia to either nitric or nitrous acid. Heraeus's work was under continual attack, but was not entirely without support.
While to some the problem of nitrification was solved, others failed to be satisfied with the answers put forth by the leading authorities. One of the men not entirely satis-fied was Jordan (3). He had seen for himself that Boston city water delivered to the laboratories at Massachusetts Insti-tute of Technology when used for ammoniacal solutions would always nitrify. This nitrification was prevented by heating the water before using. Therefore, it seemed reasonable to Jordan to assume that the nitrifying organisms were present
in the Boston tap water. This seemed even more so when a sterile ammoniacal solution inoculated with 1 ml of Boston tap water produced nitrification.
The next step in the solution of the problem was the isolation of the organisms from the water. Once this had been done, ammoniacal solutions were inoculated with the pure cultures. In all cases there was not any nitrification But Jordan was not easily discouraged as he knew that the water contained something capable of nitrification. Mixed samples of pure cultures were tried in order to determine if the bacterial action was dependent on several organisms
rather than just one. After a considerable number of nega-tive results, success was obtained. One of the ammoniacal solutions had undergone nitrification. But further investiga-tions failed to reproduce the desired results. What was
thought to be a known mixed culture turned out to be an un-known mixed culture. After exhausting all possible channels, Jordan decided that maybe it was his method of isolation of
the organisms that was at fault. Maybe the nitrifying organisms would not grow on gelatin. Thus, work was begun on growing
the bacteria in a liquid medium which had fallen into some dis-repute when it was learned that liquid media was not suitable for growing pure cultures.
Frankland (3), still investigating the phenomenon of nitrification, also came to the conclusion that the gelatin medium was at fault and resorted to a liquid medium. By many
careful dilutions and transfers, Frankland isolated an organ-ism capable of nitrifying ammoniacal solutions.
Winogradsky (3), working independently from Jordan and Frankland, had also succeeded in isolating an organism cap-able of nitrification. This had been accomplished largely
through microscopic analyses and the growing of the organisms in a solution devoid of organic matter. This latter fact opened a new field to the bacteriologists. Up until this point it was considered a foregone conclusion that the
bac-teria had to have organic matter in which to grow.
Jordan continued his work and soon had isolated organisms capable of nitrification. While Winogradsky's and Frankland's bacteria carried the nitrification to the nitrite stage,
Jordan's organisms carried the process to the nitrate stage. Winogradsky (1) continued his study of nitrification. In 1892 he noted that nitrite forming bacteria existed in both the motile state and the zoogleal state. The zoogleal state was assumed to represent a resting stage in the devel-opment of the organisms.
The zoogleal aspect of bacteriology continued to draw the interest of some. Flugge (1) in 1896 stated that the zooleal masses were generally composed of the same type of cell. This led to the conclusion that they were pure cul-tures. The big discussion of the day was whether the bac-teria were imbedded in a gelatinous matrix which they had synthesized or whether they were residing in a
chemically-formed matrix.
At the turn of the century Beijerinck (4) began work-ing with the problem of isolatwork-ing aerobic organisms capable of nitrification. In 1901 he was successful in isolating aerobic nitrifiers. Thus the organisms capable of nitrifi-cation aerobically and the zoogleal masses forming the body of the sludge had been successfully isolated and studied to a slight degree.
While the bacteriologist argued about the mechanisms of biological oxidation of ammonia and the existence of a zoogleal stage in the bacterial life cycle, the chemists were concerned with the removal of colloids from sewage and the chemical oxidation of organic matter. G. J. Fowler (5) used air as early as 1897 in experiments on the removal of colloids from the effluent of a chemical treatment plant. The clotting and settling of the colloids was noted. Sam-ples taken of the mixed liquor showed no reduction in organ-ic matter. At this time it was thought that the organorgan-ic matter in sewage was completely oxidized into soluble prod-ucts or gasses. Since this had not occurred, the use of air as a means of treatment was abandoned as being unsatisfactory. As a result, the discovery of the activated sludge process
was delayed about fifteen years.
Fowler happened to visit this country in 1912 in re-gards to a study on the disposal of *wage in New York City. While he was here, he visited the famous Lawrence Experiment
Station in Massachusetts. At this time an experiment was being conducted on the treatment of sewage by algae. The theory being tested concerned the formation of nascent oxygen by the algae which would chemically oxidize the organic matter in sewage. The algae were supplied with air under pressure. Although the algae failed to produce the nascent oxygen, Fowler noted that the sewage being used in the experiments was purified in about 19 hours. He also noted that the ex-perimenter did not throw out the sludge which formed but re-tained it for the next experiment.
On his way back to England Fowler reflected on his experi-ments with the chemical treatment plant effluent as well as
on the experiments he had just witnessed at Lawrence. In both cases sewage was being used as was air. Yet, in one case the organic matter was stabilized and in the other it was not. The only difference in the two experiments was that he had thrown out the settled sludge at the end of each experiment and the experimenter at Lawrence had retained it. On his re-turn to England he suggested to Ardern and Lockett that they should try to set up an experiment on the same order as the one at Lawrence using black bottles to exclude the formation of algae.
The experiments which Ardern and Lockett performed re-sulted in the formulation of activated sludge as we know it today. They concluded that there were two phases in the process, the -clotting out phase and the oxidation phase.
-11-The clotting out phase was concerned with the coagulation of colloidal matter and with the transfer of the pollute
foea the liquid to the sludge. This was believed to be strict-ly physio-chemical. The oxidation phase dealt with the sub-sequent oxidation of the adsorbed carbonaceous and nitrogen-ous materials to carbon dioxide and nitrates. This phase was strictly biological. For the most part the theory of activated sludge is the same as it was stated back in 1913.
While the activated sludge process was still in the experimental stage, J.W.H. Johnson (6) wondered what organ-isms were responsible for the transformations that took place in the aeration chamber. The study was primarily a
micro-scopic one without any attempt to identify any specific organism. One conclusion that Johnson came to was that the purification of the sewage was accomplished by soogleal masses.
Purdy (6) in 1918 had occasion to work with activated sludge treating strawboard wastes. He noted that there was a very large number of protozoa present in the sludge. He failed to follow up this study and it was not until.1923 that the first report was made on the complete study of the organisms of activated sludge. In that year Long and Buswell (7) published the results of their extensive
re-seorch. They had studied activated sludge from its formation through its full growth. In the beginning, as the sludge was forming, the Holotriche were the first to appear in large
-12-numbers. But after only a few days of aeration the
Peritrich, Carachesium and Vorticella, appeared. As the sludge matured and the zoogleal mass became stabilized, the Peritrichs predominated with occasional colonies of Hypo-trichs. The zoogleal mass was made up of filamentous bac-teria. It was felt certain that the zoogleal mass contained Crenothrix polyspora, Sphaerotilus dichotomus and Zooglea ramigera in large numbers.
This study was followed in 1926 by a similar one made in Germany by Kolkwitz (6). He reported almost identical results as Buswell and Long but carried the study a little further to include over-ripe sludge. At this point he found that the Hypotriths and Peritrichs were suppressed.
Interest in the biology of the activated sludge pro-cess increased rapidly. The year after Kolkwitz's study Harris, Cockburn and Anderson(8) examined sewage and found that the organisms were composed of 61% Bacillus aerogenes and 38% Bacillus proteus. They also noted that the numbers and types of protozoa differed considerably with the seasons of the year, operation practices and other variables. This led them to the conclusion that protozoa did not enter into the treatment process.
Agersborg and Hatfield (6) in 1929 found that sludge in preaeration systems consisted almost entirely of Zooglea and 3phaerotilus organisms. Increased aeration favored the succession of small flagellates and hypotrichous and peri-trichous ciliates.
Interest in the relative importance of bacteria and protozoa in the process increased as the third decade of the twentieth century began. O'Shaughnessy (9) suggested that the floc was formed to provide a matrix on which the bacteria could sustain themselves in the presence of moving water and that it was on this phenomenon that the activated sludge process rested. He did not believe that protozoa were essential for a good activated sludge.
In 1931 R. Cramer (10) conducted extensive experiments with protozoa in an effort to learn of their exact
impor-tance. He found that the material to which the bacteria and protozoa clung was largely organic. This organic matter was capable of being reduced by either aerobic or anaerobic bacteria. The aerobic bacteria broke down the organic matter into smaller colloidal particles which were incapable of settling. He believed further that the pro-tozoa synthesized the colloidal matter into larger particles which were capable of settling out.
At this same time Heukelekian (11) studied the effect that the protozoa had on the process by means of partial and complete sterilisation of the sewage. He found that the protozoa played a very minor part in the purification process; but that they did stimulate the bacterial pro-liferation and thereby helped to maintain the bacterial purification at a high level.
Buswell (12) presented a review of the activated sludge process from a biological viewpoint as it was known in 1931.
-14-He divided the activated sludge process into two biological stages: bioprecipitation and oxidation. Buswell felt that the bioprecipitation was caused by Chlamydobacteriaceae, fungi, larger protozoa and higher animals. The oxidation was brought about by the bacteria in the zoogleal masses which included the filamentous bacteria and occasionally Zoogloea ramigera. The zoogleal mass was believed to be made up of a synthetic gelatinous matrix similar to that
of Nostoc.
In 1935 G. P. Edwards (13) reviewed the various theories put forth to explain the mechanisms in the activated sludge process. He put the process in three separate stages rather than two. These stages were: clarification, reactivation and nitrification. The clarification process was primarily physico-chemical whereas the reactivation and nitrification stages were biological. He concluded that although the acti-vated sludge process had come a long way, very little was known of the organisms involved but that the bacteria and protozoa were both important.
The mechnism of clarification came under observation by R. J. Theriault (14) in 1935. The question as to whether or not the mechanism was physical or biological had been argued and reargued for many years without any success. Theriault reviewed all of the previous literature on the subject and concluded that the biological elements did not play a major part in the clarification stage. He believed
that the floc was the primary absorbent, apart from em-bedded bacteria, secreted enzymes or attending protozoa. This brought up the question as to the chemical composition of the sludge.
Following up the chemical aspects of the sludge,
Theriault (15) came forth with a biozeolite theory of puri-fication for activated sludge. This theory was evolved from chemical apalyses of the sludge. Theriault found
that the zoogleal masses after compensation for the bacter-ial cells had a chemical formula - 0.84 M20'R2 03'38i02; where
M20 represented interchangeable groups such as Na20, K20, CaO, Mg0 and R203 represented the groups such as A1
203,
Fe203. This was the formula for a zeolite. He showed that
the sludge readily absorbed ammonia from solution and that the ammonia could be regenerated with sodium chloride in the same manner as commercial zeolites.
The following year in 1936 Theriault and McNamee (16) investigated, adsorption by activated sludge. From their work they concluded that the gelatinous matrix served a double purpose. Not only did the matrix hold the bacterial cells together but also acted as an intermediary between the bacteria and the sewage. The gelatinous matrix seemed to adsorb the organic material and hold it until the bac-teria could assimilate it. They concluded that activated sludge was 30% inorganic zeolite.
As a result of these experiments Theriault (17) came
forth with his biozeolite theory of sewage Purification. He resolved his theory into five basic points:
(1) Inorganic colloids of sewage are zeolite derived from feces and are fully saturated with organic matter.
(2) The colloids are negatively charged and can be agglomerated by plain substances in the tanks to give negatively charged organic matter.
(3) Activation of sludge is accomplished when adsorbed organic matter is dialized to
bacterial cells, leaving the non-diffusible alumino-silicate behind for further
adsorption.
(4) Clarification of sewage is accomplished by adsorption of organic matter in sludge
zeolites without accompanying oxidation or bacterial intervention.
(5) organic cations transferred to the bacteria from the zeolites are decomposed and liber-ated to the solution by diffusion. The decomposition products are highly reactive and combine readily with the dissolved oxygen in solution.
Thus, Theriault had explained how the activated sludge pro-cess operated. To him the propro-cess was largely an exchange
between the sewage and the gelatinous matrix with the bac-teria serving only as a means of regeneration of the so-called Ozeolite".
Better, Ridenour and Henderson (18) were also intrigued by the exchange capacity of activated sludge. They felt that the exchange process progressed in two steps: the adsorp-tion of colloids and organic electrolytes and the regenera-tion through oxidaregenera-tion. The sludge and the sewage colloid were found to be negatively chatged particles as the result of electrophoretic studies. The adsorption of two similarly charged bodies was explained by the exchange of a mobile ion on the sludge matrix with a similar immoble ion on the sewage colloid. The regeneration of the sludge by oxidation caused the sludge to revert to its original properties.
While Theriault was studying the bioseolite theory, C. T. Butterfield (19) began to work on the very basis of activated sludge, the organisms themselves. He began by washing the sludge matrix to remove all extraneous organisms and then attempted to isolate the organisms that formed the main mass. At first the results were not very favorable.
The organism did not seem to grow very well in ordinary agar or gelatin media. Only after making a special agar using sludge liquid was he able to successfully isolate the organisms he believed responsible for the majority of the stabilization of the organic matter. Using pure colonies obtained from the agar, Butterfield made successful plants
into nutrient broths containing peptone. After careful study of the organisms and the literature concerning zoog-leal organisms, he concluded that the bacterium was Zooglea ramigera. Using a synthetic sewage and pure cultures,
Butterfield set up a laboratory activated sludge plant. He obtained 72% B.O.D. removals with three hours aeration.
Next sterile sewage was tried and 68% B.0.D. removals were obtained with the same aeration period.
Heukelekian (20) studied the carbon dioxide production of the sludge in relation to the oxygen uptake. He found that the Respiratory Quotient (R.Q. ), ratio of C02:02 by volume, varied considerably from 1.32 - 2.14. He also found
that the R.Q. of sewage was variable from 1.13 - 1.41. The maximum theoretical R.Q. is 1.00 for complete oxidation. The high values of Heukelekian's results were caused by the fact that the CO2 and 02 determinations were not made simul-taneously on the same substrates.
The clarification stage also attracted the interest of Heukelekian (21). The idea of an exchange phenomenon had been put forth but had been studied only slightly with am-monia as the material to be adsorbed. Heukelekian studied
a wide range of inorganic and organic substances. These substances included infusorial earth, Kaolin, butyric acid, dextrose, caesein, peptone, ammonia and B. coli. He found that 30% of the infusorial earth was removed in thirty minutes by aeration of sl(:dge solids or by plain mixing.
The Kaolin was reduced 25% in this same period; whereas butyric acid, dextrose, caesein, peptone and ammonia gave little or no reduction. The numbers of B. coli increased rapidly over a 10 minute aeration period. The greatest increase came from the sample having the lowest bacterial concentration at the start.
E. V. Mills (22) studied the sewage colloids and the
mechanism of their removal. He determined the electrophoretic velocities of various sewage colloids at various pH levels. It was learned that the electrophoretic velocities were pro-portional to the pH. The colloids had a negptives charge above the pH 4 and a positive charge below this pH. He did not feel that the electrolytes in the sewage caused the charge on the colloids but that the charge was due to ioni-zation of the particle surface. The surface was believed to be made up of condensed soaps. As a result of his studies Mills wrote that activated sludge was nothing more than ag-gregates of minute particles fabricated by the active bac-teria.
Wooldridge and Standfast (23), studied the rates of oxidation using suspensions of pure cultures of various or-ganisms found in sewage. They studied several protozoa as well as bacteria. No effort was made to tie these organisms into the activated sludge process or to any sewage treatment method. Soe of the organisms which they investigated were B. alkaligenqs, I. coli. A. aerogenes, Ps. vulgaris, Ps.
-20-fluorescens, Chr. prodigiosum, Polytoma and washed acti-vated sludge. In the presence of the protozoa, Polytoms, ps.
Sluorescens exerted the highest oxygen demand of any of the
above organisms. The rate of oxidation was found to be de-pendent on the number of viable cells. The period of study was limited to a few hours .
In 1937 Butterfield, Ruchhoft and McNamee (24) studied the oxidation of pure culture sludges using Zooglea ramigera. They used two substrates: sterile raw sewage and a synthetic sewage. The synthetic sewage that was used did not contain any carbohydrates other than that normally present in pep-tone or beef extract. They learned that 50% of the 5-day
B.0.D. was removed in five hours aeration and 80% in 24 hours. Nitrogenous materials were not included in this oxidation
as the bacteria were not capable of such action. They did not feel that Z. ramigera was the only organism capable of rapid oxidation under aeration. Although they knew of no other organisms which were capable of such a phenomenon, they felt that the primary prerequisite of such an organism would be the ability to produce a floc that would hold to-gether during aeration.
Heukelekian (25) continued his study into the clari-fication stage by expanding his first studies slightly.
He learned that if he saturated activated sludge with sewage colloids the clarification was impaired; but that if he
unimpaired. The effect that butyric acid and ammonia had on the clarification was determined and found to be nought. From this he concluded that the removal of Kaolin was
ac-complished by either adsorption or ion exchange and that the removal of sewage impurities was a biological phenomenon.
This series of studies was continued by Heukelekian
and Ingols (26) with a study on the carbon dioxide production during the clarification and oxidation stages. It was learned
that greatest rates of carbon dioxide production and oxygen uptake occurred during the initial period of aeration. This meant that the bacteria were the most active when the sewage came into contact with the sludge and that the concentrated
activity was dependent on the quantity of food material present. They felt that the division of the activated sludge process
into the two divisions, clarification and oxidation, was not warranted since both proceeded simultaneously.
In 1938 Butterfield, Ruchhoft and Mciamee (27) collabor-ated to study the oxidation by normal activcollabor-ated sludge rather than by pure culture sludges. They learned that the capacity to oxidize food at a high rate was developed in the sludge as a result of continuous feeding. The oxidation capacity of the sludge was believed to be limited and therefore only enough sludge should be maintained in the aeration tank for optimum oxidation.
This same year Butterfield and Wattie (28) joined in an experiment dealing with the numbers of bacteria and the
-22-dispersion of floe and their relationship to the oxidation of organic matter. Their results showed that oxidation took place as the result of the metabolic activity and prolifera-tion of living bacteria. The magnitude of the oxidaprolifera-tion was dependent on the number of living cells produced which were in turn dependent on the amount of food available.
The rate of oxidation was also found to be affected by the degree of dispersion of the floe.
In 1938 Wilson and McLachlan (29) looked into the matter of carbon dioxide production from the standpoint of where the carbon present in the sewage goes. It was earned that 10% of the organic carbon was oxidized to carbon dioxide, while the remaining 90% of the carbon went into bacterial
synthesis.
Continuing the clarification stage series, Heukelekian and Schulhoff) (30) studied the organisms which were res-ponsible for clarification now that they had learned that
biological activity was very great during this period.
They isolated a number of organisms from clarified sewage in contact with activated sludge under quiescent conditions and began to study the organisms. The organisms were placed
in sterile sewage and the sewage was aerated. The sewage cleared much more rapidly than by settling alone. Still, the pure cultures were not able to clear the sewage as well as natural activated sludge. This indicated that all of the organisms responsible for clarification had not been
isolated or that there was also a secondary mode of clarifi-cation present. The clarificlarifi-cation of the sterile sewage with the pure cultures was accompanied by a transformation
of some suspended matter to soluble matter. Some of the organisms accomplished clarification by oxidation and others by flocculation mechanisms.
R . S. Ingols (31) in 1938 came forth with the next
experiment in this series. It was conducted to show the physical properties of adsorption of the sludge floc. This tas done by taking ordinary activated sludge and adding glucose to the incoming sewage. The sludge growth was
stimulated by the glucose. Next, ammonia was added and its adsorption by the sludge was noted. Ammonia and glu-cose were added together with a resulting increase in the ammonia uptake. Since ammonia and glucose did not enter
into direct chemical reaction, it was assumed that the bacterial cells responsible for ammonia stabilization were stimulated by the glucose. In order to test the base ex-change capacity of the sludge, chloroform was added to the sludge to inactivate the bacterial cells without damaging the gelatinous matrix which had been assumed capable of base exchange. There was no uptake of ammonia by the bio-logically inactive sludge, indicating the complete dependence of the uptake of ammonia upon the biological phenomena of the sludge without the help of so-called physical processes. In the light of present knowledge concerning the surface structure
of the bacterial cell, the chloroform probably reacted with the chemical groups responsible for adsorption and prevented adsorption.
Heukelekian and Ingols (32), that same year, presented their work on the hydrolysis of starch. The starch was hydrolyzed very rapidly with the rate of hydrolysis
depen-dent on the quantity of starch, concentration of sludge, presence of sewage, adequate air supply and quality of
sludge. The hydrolysis of starch did not fesult in an ac-cumulation of the products of hydrolysis which indicated that they were oxidized as rapidly as they were formed.
Next in this series was a study of complex nitrogenous materials. Heukelekian and Ingols (33) continued their studies using complex proteins in feeding the activated sludge. The protein molecules were first hydrolyzed by
the bacteria by means of extra-cellular enzymes. The pro-tein hydrolyzed to proteoses, peptones and peptides which further hydrolyzed to amino acids. The bacteria used the amino acids as a source of energy, breaking them down into ammonia, carbon dioxide and water. The ammonia served as a source of nitrogen for synthesis of new protoplasm if sufficient carbon was present. The ammonia also served as food for the nitrifying bacteria. The order of biological action, hydrolysis and reduction of B.O.D. varied inversely with the complexity of the molecules and directly with the
Lastly, Ingols (34) made a study on the uptake of sol-uble organic compounds. His study confirmed the previous one, showing that amino acids follow the laws of biologi-cal activity in their disappearance from the liquid. When glucose was present in large quantities, the amino acids were only slightly used as a source of energy and less
am-monia was produced. In all cases the soluble organic
sub-stances were removed from solution by biological or bio-chemical processes independent of purely physical phenomena.
While working with dyestuffs and activated sludge, Dickinson (35) found that the dyes containing
electroposi-tive ions were readily adsorbed to the sludge. This
indicated that the sludge was made up of negatively charged particles.
Sawyer and Nichols (36) studied the Respiratory Quo-tient of activated sludge in 1939. They found that on the whole the R. Q. values that they obtained were about equal
to the theoretical R. Q. values for the substrates under consideration. The R. Q. of a sludge-sewage mixture was found to vary with the nature of the sludge used. In their experiments the R. Q. of sludge-sewage mixtures varied from
0.77 to 1.18.
The total purification, oxidation, adsorption and syn-thesis of nutrient subatrates by activated sludge came under the study of Ruchhoft, Butterfield, McNamee and Wattie (37).
Their work included both pure culture and normal activated
-26-P
sludge, using synthetic and sterile raw sewage as substrates. The pure cilture zoogleal sludges oxidized 50% of the total B.O.D. in 24 hours. The greater portion of work took place
during the first 30 minutes of the aeration period. During this period 30-50% of the B.0.D. was removed by adsorption
and synthesized while 16-21% was being oxidized. These figures were slightly lower for normal activated sludge for the same aeration period, 40% adsorbed and synthesized and 10% oxidized.
The problem of carbon and nitrogen transformation was studied by Heukelekian and Littman (38). They set about to find out if activated sludge contained one or more species of zoogleal organisms, the behavior of the organisms under varying environmental conditions, the biochemical reactions brought about by the organisms and the C/N ratio of pure
sludges. In all, 15 different act1Vated sludges were studied. The isolation of the organisms followed the technique set
up by Butterfield. This consisted of taking a small clump of sludge in a capillary pipette and placing it in distilled water. This solution was centrifuged 30 seconds at 750 RPM. The supernatant was poured off and the process repeated 12
times. The final tube was shaken violently and then planted in nutrient broth. The tubes showing growth in the highest dilutions were used to plant on nutrient agar. The colonies picked off the agar plates were replanted in nutrient broth and the process was repeated many times until relatively pure
cultures were obtained. The organisms isolated from the sludge belonged to the class of Zooglea ramigera, being gram negative rods, 1.5 - 3.5u long, 1 - 2u
wide,
capsu-lated and motile in young cultures. In nutrient broth only slow growth was obtained; agar colonies were very small, al-most pinpoint in size. There was a slight growth with gela-tin stab and no liquefaction. A 0.2% starch broth failed to give hydrolysis and carbohydrate broth failed to result in acid formation, although moderate growth was obtained. The pure cultures were next used to set up a laboratory acti-vated sludge plant. The sludge was grown in cotton filtered, heat sterilized sewage containing 100 ppm glucose and 100 ppm urea and aerated under sterile conditions. When glucose and urea were added, the turbidity increased. This increase in turbidity was believed to have been caused by bacteria and that under normal operating conditions, the protozoa fed on these cells, reducing the turbidity. The C/N ratio of this sludge was 4-5:1.
Using the preceding study as a basis of attack,
Heukelekian and Littman (39) made a concentrated study on the purification qualities of pure zoogleal- sludges. These zoogleal sludges, having varying concentrations of
sus-pended solids from 526 - 1777 ppm, were aerated with sterile sewage a sufficient period for purification and then over-rated. Pure culture sludges reduced the turbidity 58 - 70% and the B.0.D. 60 - 90% in 24 hours aeration. Early removal
-28-of suspended carbon materials was attributed to adsorption. The suspended organic carbon was removed from solution be-fore the dissolved organic carbon was. The period of bio-logical assimilation by the pure cultures resulted in an
increased 002 production. They were able to attribute one-half of the total 002 production by activated sludge to the zoogleal organisms. The C/N ratio was essentially the same in this experiment as the preceding one, 4.7:1. Zooglea ramigera was identified as the primary organism capable of purifying sewage with the nitrifying bacteria and protozoa acting as secondary organisms.
Jenkins and Roberts (40) found that 20% of the organic carbon removed was oxidized to carbon dioxide in 24 hours. This was double the value set forth by Wilson and McLachlan
(29). They also learned that 44% of the organic carbon re-mains in the sludge.
Tomlinson (41) set forth the biology of the activated sludge process as it was known in 1939. He felt that the majority of the protozoa present feed on other organisms. Since bacteria were so plentiful, it was thought that they
were the common source of food.
Masazo Watanabe (42) put forth a series of four articles on the activated sludge process as it was known in Japan. He studied the biological action, the enzymic action,
ad-sorption and the sludge ash constituents. The enzymatic study resulted in the demonstration that such hydrolytic
_ --A
ensymes as the amylases, proteases and lipases existed in bacteria-free effluents from activated sludge plants.
The adsorption study showed that saccharides were adsorbed in weak solutions, fatty acids were adsorbed in relation to their molecular weight and that iodine was also adsorbed. The ash study consisted of both polarographic and chemical determinations of the various components. The oxides of the common metals only made up 58.5% of the ash content.
The problem of removing glucose from substrates by activated sludge was the next attacked. In 1940 Ruchhoft, Kachmar and Moore (43) found that glucose was removed much more rapidly from solution by natural activated sludge than by domestic sewage alone, pure cultures of Bacillus coli,, Bacillus aerogenes, Sphaerotilus natans or zoogleal sludge. The rate of removal of the glucose was a function of the
sludge which required a definite aeration to maintain the high rate of glucose removal. Yet, the ratio of oxygen to glucose removed was relatively low. If glucose was added
to a sludge alone, the sludge was retarded in glucose ad-sorption until the sludge finally refused to take up glu-cose at all. Thus, the value of nitrogen or phosphorus was becoming evident in the normal function of the sludges.
The sludge was also able to acclimate itself to high dosages of glucose under proper nutritional requirements. This
ac-climatization phenomenon was explained on the basis of multiplication of certain special glucose removing
-30-organisms or by the development of adaptive glucose enzymes of the predominant bacteria of the sludge.
Ruchhoft, Kachmar and Placak (44) continued the study of glucose metabolism using normal activated sludge and pure culture sludge. The normal activated sludge oxidized 4 - 24% of the glucose in one and one-half hours and 11 -32% in 24 hours; whereas the pure culture sludge oxidized
31% of the glucose in one and one-half hours and 45% in
24 hours. Almost all of the glucose removed within one and one-half hours aeration period was assimilated and appeared as protoplasm. The glucose removal in pure
cul-ture sludge was adaptive and could be accelerated by re-peated feedings. From 80 - 100% of the glucose removed was accounted for as the sum of glucose oxidized and that assimilated to protoplasm.
N. W. Barritt (45) made a study of the ecology of activated sludge. As a result of previous work, he felt that Z. ram and other bacteria were present. Of the protozoa present he found Amoeba, Chilodon, Colpidium and Glaucoma. In one experiment the sludge was aerated for ten days without feeding. The protozoa died and the sludge be-gan to disintegrate. He concluded that the protozoa were unable to live off of the floc alone. All of his efforts to isolate Z. ramigera by either the plating or dilution method ended in failure.
P
with activated sludge came under scrutiny by Ruchhoft and Kachmar (46). The purpose of their study was to determine
the exact function that Sphaerotilus natans had in bulking. They believed that bulking was a response of all the sludge organisms to a sudden disturbance in the biological
equil-ibrium. The disturbance affected, primarily, the biophysi-cal character of the sludge as was indicated by a reduction
in short time adsorption capacity and by the formation of a light, non-compact floe. The oxidizing capacity of the sludge was not affected by the disturbance.
The zoogleal organisms found on the trickling filter were found by Butterfield and Wattie (47) to be very
sim-ilar to those found in activated sludge. They took a mix-ture of several of their zoogleal organisms isolated from trickling filter slimes and set them up in aeration units. The average B.O.D. removal in 24 hours was 87%, while 72% of the B.O.D. removed was oxidized.
Although Butterfield had succeeded in isolating Z_. ramigera, it was not until 1942 that the cultural charac-teristics of this organism were studied in great detail. Elsie Wattie (48) used Butterfield's technique in isolating
Z. ramigera and improved on it slightly. The technique employed in this study consisted of placing 10 ml. of sludge in a 30 ml. sterile bottle with several sterile glass beads. The bottle was agitated very violently for ten minutes. The remainig portions of the sludge were
planted in serial dilutions in lactose broth and incubated at 2000 for 48-120 hours. Using the tube showing growth
in the highest dilution, plants were made on standard nutrient agar diluted 1:3. After 96 hours incubation at
2000, colonies were picked off the agar and planted in
lactose broth. This procedure was repeated several times until it was felt certain that pure cultures had been ob-tained. In all, fourteen zoogleal forming organians were isolated; but for all practical purposes they could be
considered in one group. They were all short, gram
nega-tive rods incapable of producing H28 or acetyl methyl
carbinal, but capable of acid .production with arabinose and not with raffinose. Wattie used pure culture sludges containing 1500 ppm suspended solids in a small scale
acti-vated sludge unit and found that 36-84% of the B.O.D. was removed in three hours aeration and 55-92% in five hours aeration. At the same time studies were made to determine
the minimum food requirements, the effect of pH, soap,
sodium ricinoleate, sodium formate, pectin, glucose, glycer-ine and sucrose on the organisms.
. B Phelps and J. G. Bevan (49 ) tried to show that it was possible to build a biochemical floc from A. aerogenes and 5,ppm FeSO4. The problem of contamination was so great that it was impossible to maintain pure cultures for longer than a few days or a week at most. They felt that if the cultures could have been kept pure over a sufficiently long