Ecosystem engineers’ contribution to soil structure formation in floodplains
152
0
0
Texte intégral
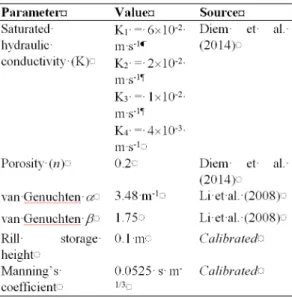
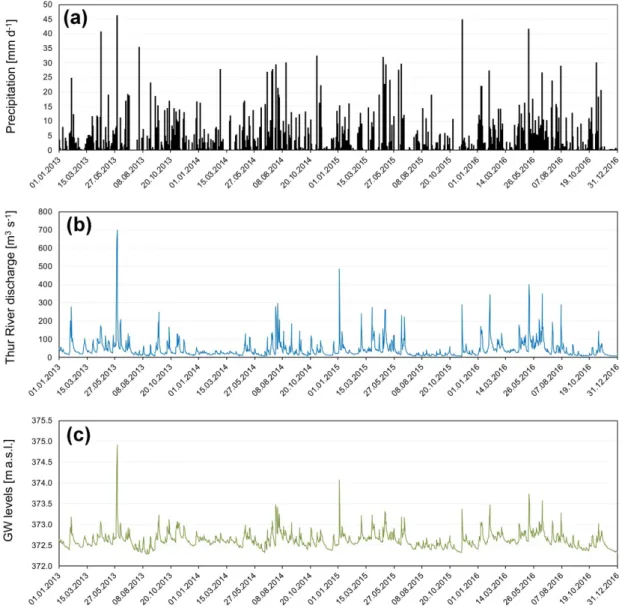
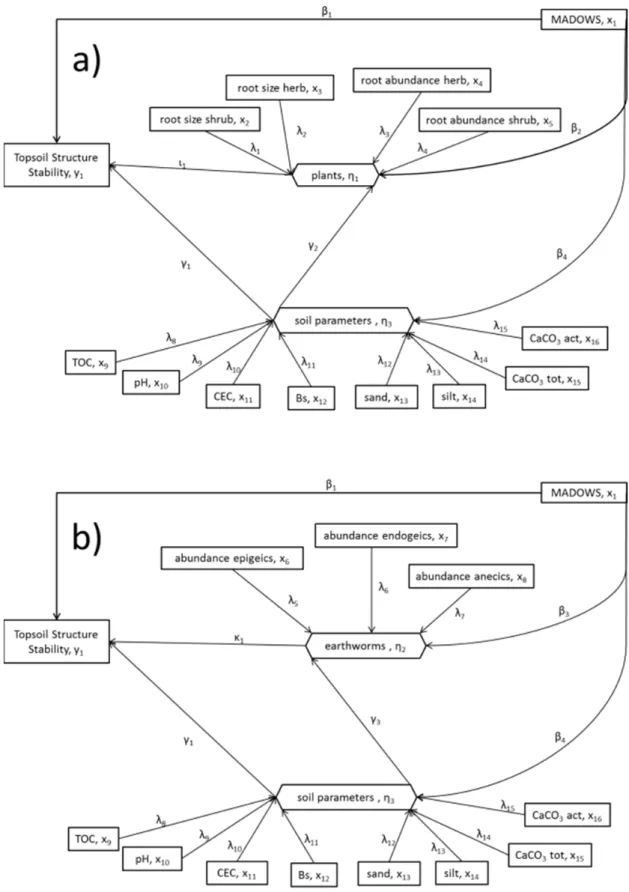
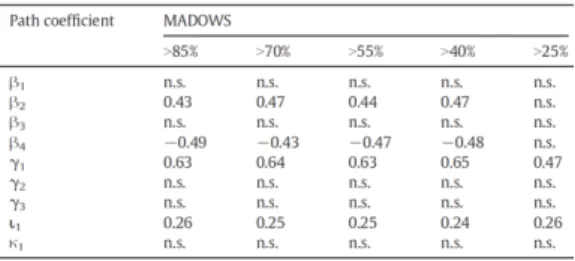
Figure

+7

Documents relatifs
