Dynamic and specific interaction between
synaptic NR2-NMDA receptor and PDZ proteins
The MIT Faculty has made this article openly available.
Please share
how this access benefits you. Your story matters.
Citation
Bard, L., M. Sainlos, D. Bouchet, S. Cousins, L. Mikasova, C. Breillat,
F. A. Stephenson, B. Imperiali, D. Choquet, and L. Groc. “Dynamic
and specific interaction between synaptic NR2-NMDA receptor and
PDZ proteins.” Proceedings of the National Academy of Sciences
107, no. 45 (November 9, 2010): 19561-19566.
As Published
http://dx.doi.org/10.1073/pnas.1002690107
Publisher
National Academy of Sciences (U.S.)
Version
Final published version
Citable link
http://hdl.handle.net/1721.1/84594
Terms of Use
Article is made available in accordance with the publisher's
policy and may be subject to US copyright law. Please refer to the
publisher's site for terms of use.
Dynamic and specific interaction between synaptic
NR2-NMDA receptor and PDZ proteins
Lucie Barda,b, Matthieu Sainlosa,c, Delphine Boucheta,b, Sarah Cousinsd, Lenka Mikasovaa,b, Christelle Breillata,b, F. Anne Stephensond, Barbara Imperialic, Daniel Choqueta,b, and Laurent Groca,b,1
aLaboratory for Cellular Physiology of the Synapse, Centre National de la Recherche Scientifique, Unité Mixte de Recherche 5091, 33077 Bordeaux, France; bUniversité de Bordeaux, 33077 Bordeaux, France;cDepartments of Chemistry and Biology, Massachusetts Institute of Technology, Cambridge, MA 02139; and dSchool of Pharmacy, University of London, London WC1N 1AX, United Kingdom
Edited by Richard L. Huganir, The Johns Hopkins University School of Medicine, Baltimore, MD, and approved October 4, 2010 (received for review March 3, 2010)
The relative content of NR2 subunits in the NMDA receptor confers
specific signaling properties and plasticity to synapses. However,
the mechanisms that dynamically govern the retention of synaptic NMDARs, in particular 2A-NMDARs, remain poorly understood. Here, we investigate the dynamic interaction between NR2 C termini and proteins containing PSD-95/Discs-large/ZO-1 homology (PDZ) scaffold proteins at the single molecule level by using high-resolution imaging. We report that a biomimetic divalent competing ligand, mimicking the last 15 amino acids of NR2A C terminus, specifically and efficiently disrupts the interaction between 2A-NMDARs, but not 2B-2A-NMDARs, and PDZ proteins on the time scale of minutes. Furthermore, displacing 2A-NMDARs out of synapses lead to a compensatory increase in synaptic NR2B-NMDARs, pro-viding functional evidence that the anchoring mechanism of 2A- or 2B-NMDARs is different. These data reveal an unexpected role of the NR2 subunit divalent arrangement in providing specific anchor-ing within synapses, highlightanchor-ing the need to study such dynamic interactions in native conditions.
lateral diffusion
|
glutamate receptor|
trafficking|
biomimetic multivalent ligand|
developmentT
he identification of the cellular mechanisms involved in the regulation of glutamate receptor trafficking is crucial to our understanding of synaptic maturation and plasticity. One com-mon paradigm of these processes is the activation of the calcium-permeable postsynaptic NMDA receptors (NMDARs). In the neocortex, the most abundant types of NMDARs are composed of NR1 subunits associated with NR2A (enriched in synapses) and/or NR2B subunits (1). Rapid changes in the synaptic 2A/2B NMDAR ratio have been reported during connection refine-ments and synaptic plasticity (2), and several key molecular interactions have been shown to control the trafficking of in-tracellular and membrane NMDARs (3–6).The intracellular proteins that interact with the C terminus of the subunits, through direct binding or modification of the phosphorylation state, are likely candidates for regulating the synaptic retention of NMDARs. Indeed, intracellular domains of NR2 subunits provide a binding motif for proteins of the post-synaptic density such as PSD-95 and SAP102 (7–10). The binding of the NR2B subunit C terminus to PDZ domain-containing scaffold proteins regulates, in part, the synaptic retention of this receptor (8, 9, 11–14). For the 2A-NMDARs, which make up the majority of synaptic NMDARs, the role of such interactions in synaptic retention remains controversial. Indeed, long-term ex-pression of NR2A subunits with a truncated or mutated C ter-minus does not affect synaptic NMDAR currents in cerebellar or hippocampal neurons (9, 15), whereas deletion of the NR2A subunit C terminus sequence significantly reduces NMDAR synaptic signaling (11, 14, 16, 17). Currently, there is no simple explanation for this discrepancy, and the use of long-term ex-pression of exogenous NR subunits and lack of good pharmaco-logical tools to discriminate between 2A- or 2B-NMDAR
signaling (18) render interpretation more difficult. Here, we apply biomimetic divalent peptide-based competing ligands to acutely interfere with the PDZ domain-containing scaffold proteins-2A-NMDAR interaction and use single quantum dot (QD) tracking to image, with subwavelength precision, the dynamics of surface synaptic NMDARs.
Results
Design of a Biomimetic Multivalent Ligand to Disrupt the Interaction Between NR2A Subunit and PDZ Domain-Containing Scaffold Proteins
(PDZ Proteins). The molecular mechanisms involved in the
dy-namic retention of 2A-NMDARs within postsynaptic membranes are not defined. To investigate these mechanisms, we developed a peptide-based ligand that strongly and acutely perturbs the interaction between NR2A subunit and PDZ proteins (Fig. 1A). Similar strategies have previously been used to dissociate the PDZ scaffold–NMDAR interaction (19–21). In these studies, disruption of the PDZ protein–NMDAR interaction was ach-ieved by using monovalent peptide sequences that corresponded to the last nine to 10 residues of a single subunit (NR2A or NR2B). We reasoned that the efficiency of such an approach could be improved by using synthetic ligands that would better mimic the native interactions. Indeed, because (i) NMDARs are heterodimeric complexes composed of NR2 subunit dimers and (ii) the scaffold proteins (e.g., PSD-95, PSD-93 and SAP-102), which interact with NMDARs, each contain clusters of PDZ domains that recognize similar targets (22), we hypothesized that a ligand composed of two NR2 C-terminal binding motifs would more efficiently dissociate the native scaffold PDZ domain– NMDAR interactions. In the current design, we conjugated two of the 15 residue C-terminal sequences of the PSD-95 NR2A binding motifs via their N-termini (Fig. S1A and B). Homologous monovalent sequences and a previously described nonsense se-quence were used as controls (Fig. S1 A–D) (23). A series of ligands incorporating a solvatochromicfluorophore was first used to evaluate the binding constants with recombinant PSD-95 PDZ domains 1 and 2 (23). The divalent ligand displayed a sevenfold increase of affinity for the tandem domain in comparison with the monovalent homolog (Fig. S1C), confirming the advantage of
divalency. The ligands were then appropriately modified for specific experiments, e.g., for cell studies, by addition of a TAT cell-transduction sequence to generate TAT-NR2A15 or
TAT-[NR2A15]2and/or a labeling dye (BODIPY-fluorescein).
Author contributions: F.A.S., B.I., D.C., and L.G. designed research; L.B., M.S., D.B., S.C., and L.M. performed research; M.S., C.B., and B.I. contributed new reagents/analytic tools; L.B. analyzed data; and L.G. wrote the paper.
The authors declare no conflict of interest. This article is a PNAS Direct Submission.
1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online atwww.pnas.org/lookup/suppl/doi:10. 1073/pnas.1002690107/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1002690107 PNAS Early Edition | 1 of 6
NEUROSCI
Interaction Between 2A-NMDARs and PDZ Proteins Regulates NMDAR
Synaptic Retention. To investigate the ligand efficacy in neuronal
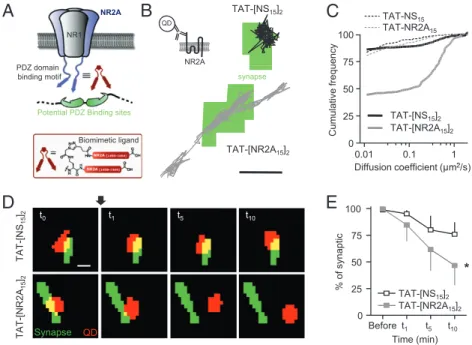
preparations, hippocampal cultured neurons were incubated with a saturating (5–10 μM) nontoxic (i.e., no neuronal damage ob-served) concentration of TAT-[NS15]2or TAT-[NR2A15]2. After
a 10-min incubation period, neurons were efficiently labeled by the different BODIPY-containing ligands (SI Materials and Methods). To investigate the specific impact of the ligands on the surface 2A-NMDAR anchoring, we used single QD tracking as a high-resolution approach to estimate 2A-NMDAR surface diffusion in live neurons (24). Native 2A-NMDARs were detected by using a QD-antibody complex directed against the extracellular N terminus of the NR2A subunit (Fig. 1B andSI Materials and Methods) and their surface localization, i.e., onto a postsynaptic marker or out-side synapse, determined during the recording session. The overall surface diffusion of 2A-NMDARs was increased after monovalent TAT-NR2A15and divalent TAT-[NR2A15]2ligand incubation,
al-though to very different extents: (i) the cumulative distribution of coefficient diffusion was highly shifted by TAT-[NR2A15]2
in-cubation; (ii) the diffusion coefficient medians were threefold and 27-fold increased by TAT-NR2A15 and TAT-[NR2A15]2,
re-spectively [i.e., TAT-[NS15]2median of 4.10−3μm2/s, interquartile
range (IQR) of 0–2.10−2 μm2/s,
n = 530 trajectories; TAT-[NR2A15]2median of 11.10−2μm2/s, IQR of 6.10−4-5.10−1μm2/s,
n = 303 trajectories; P > 0.05]; and (iii) the fraction of mobile 2A-NMDARs (membrane diffusion>0.0075 μm2/s) increased by 2%
and 42% after TAT-NR2A15 and TAT-[NR2A15]2, respectively
(Fig. 1C). Similar results were obtained when examining solely synaptic 2A-NMDARs (Fig. S2), indicating that disruption of the 2A-NMDAR anchoring increases the fraction of mobile receptors. To investigate the impact of TAT-[NR2A15]2 on identified
single synaptic 2A-NMDARs, QD-2A-NMDAR complexes were
tracked within synapse before and in the presence of TAT-[NR2A15]2 or TAT-[NS15]2 (Fig. 1D). After TAT-[NS15]2
in-cubation (10 min) the fraction of 2A-NMDARs that remained within synapses was unchanged, although a slight but not signifi-cant (P > 0.05) decrease is noted, consistent with the basal ex-change rate of surface NMDARs between synaptic and extra-synaptic membranes (25–27). However, within the same time frame in the presence of TAT-[NR2A15]2, approximately half of
the synaptic 2A-NMDARs escaped the synaptic area (Fig. 1 D and E), indicating that anchoring of synaptic 2A-NMDARs by PDZ scaffolds is a dynamic process. To further confirm the im-pact of the ligand on the surface NMDAR synaptic population, and not only single receptor, we expressed the NR1 subunit (obligatory subunit of surface NMDARs) fused to Super Ecliptic pHluorin at its extracellular N terminus (SEP-NR1) to isolate the surface fraction and quantify the average surface diffusion of SEP-NR1-containing NMDARs usingfluorescence recovery af-ter photobleaching (FRAP;Fig. S3A–C). The recovery of SEP-NR1fluorescence in dendrites was approximately 50%, whereas it was only 20% in synapses (25). Consistently, a decrease of the percentage of immobile synaptic receptors was observed, i.e., from 85% before incubation with TAT-[NR2A15]2to 65%
fol-lowing 20 min incubation (Fig. 1E). The proportion of immobile receptors outside synapses was not affected, suggesting that TAT-[NR2A15]2acts on synaptically enriched 2A-NMDARs (11,
28). Finally, these results were further confirmed with immuno-cytochemical staining of synaptic NR2A subunits (colabeled with PSD-95), as, over a large fraction of synapses (TAT-[NR2A15]2,
n = 1,684 synapses; TAT-[NS15]2,n = 1,981), TAT-[NR2A15]2
consistently reduced the synaptic content of 2A-NMDARs (Fig. S3D and E).
B
TAT-[NS 15 ] 2 75 100 TAT-NS15 TAT-NR2A15 u encyC
A
PDZ d i NR1 NR2A QD NR2A TAT-[NR2A 15 ] 2 0.01 0.1 1 0 25 50 TAT-[NS15]2 TAT-[NR2A15]2 Diff i ffi i t ( 2/ ) Cumulative freq u PDZ d oma i n binding motif Biomimetic ligandPotential PDZ Binding sites
sy napse
D
E
t 0 t 1 t 5 t 10 Diffusion coefficient (µm2/s) TAT-[NS 15 ] 2 * 50 75 100 synaptic Synapse QD TAT-[NR2A 15 ] 2 Before t1 t5 t10 0 25 Time (min) % of TAT-[NR2A15]2 TAT-[NS15]2Fig. 1. Acute disruption of the interaction between 2A-NMDARs and PDZ proteins using a NR2A-derived multivalent ligand. (A) Schematic representation of a membrane NMDAR, a scaffold MAGUK protein, and the newly developed divalent ligand mimicking the C terminus (15 last amino acids) of two NR2A subunit subunits (TAT-[NR2A15]2). (B) Representative trajectories of surface 2A-NMDARs, based on QD-coupled antibodies against an extracellular epitope of the NR2A
(Upper Left), after 10 to 20 min of incubation with TAT-[NS15]2(Upper) or TAT-[NR2A15]2(Lower). The green areas correspond to synaptic sites labeled with
Mitotracker. (Scale bar: 1μm.) (C) Cumulative distribution of the instantaneous diffusion coefficient of 2A-NMDARs. The first point corresponds to the percentage of immobile receptors (bin size, 0.0075μm2/s). Note the higher increase in the mobility of 2A-NMDARs induced by the divalent TAT-[NR2A
15]2(n = 303 trajectories;
solid gray line) compared with monovalent TAT-NR2A15(n = 170; dashed gray line) or TAT-NS15(dashed and solid black lines; TAT-[NS15]2, n = 530 trajectories;
TAT-NS15, n = 153 trajectories). (D) Displacement of individual NR2A-coupled QDs after incubation with TAT-[NR2A15]2. The neurons were incubated with Mitotracker
(green) and NR2A-coupled QDs (red spots). The localization of NR2A-coupled QDs was followed for 10 min after acute addition (arrow) of 5μM TAT-[NS15]2
(Upper) or 5μM TAT-[NR2A15]2(Lower). (Scale bar: 1μm.) (E) The synaptic localization of NR2A-coupled QDs decreased over the 10 min recording after incubation
with both TAT-[NS15]2(n = 11) and TAT-[NR2A15]2(n = 6). The reduction was significantly higher for TAT-[NR2A15]2(*P< 0.05).
NR2A-Derived Ligand Does Not Interfere with 2B-NMDAR, Kv
Potassium Channel, or GABAA Receptor Surface Trafficking. The
NR2 subunits are thought to associate with PSD-95 via a C-terminal 4-aa sequence, which is identical in NR2A and NR2B subunits. Other upstream amino acid sequences that differ be-tween NR2A and NR2B subunits have also been implicated in PSD-95 binding (29), and there is some evidence that, at least for potassium channels, binding to PSD-95 tandem PDZ domains involves up to 12 C-terminal residues (30). Interestingly, the amino acid sequence homology decreases to only 60% after alignment of NR2A and NR2B subunit 15 C-terminal residues (Fig. S1). Although most studies on isolated PDZ domains and minimal peptides derived from the C-termini of binding partners tend to limit the ligand interacting residues to the C-terminal 4 aa, we anticipated that the native interactions might achieve higher specificity by engaging additional residues, constituting the rationale for using the last 15 aa of the NR2A subunit. We first analyzed the effect of TAT-[NR2A15]2 on native
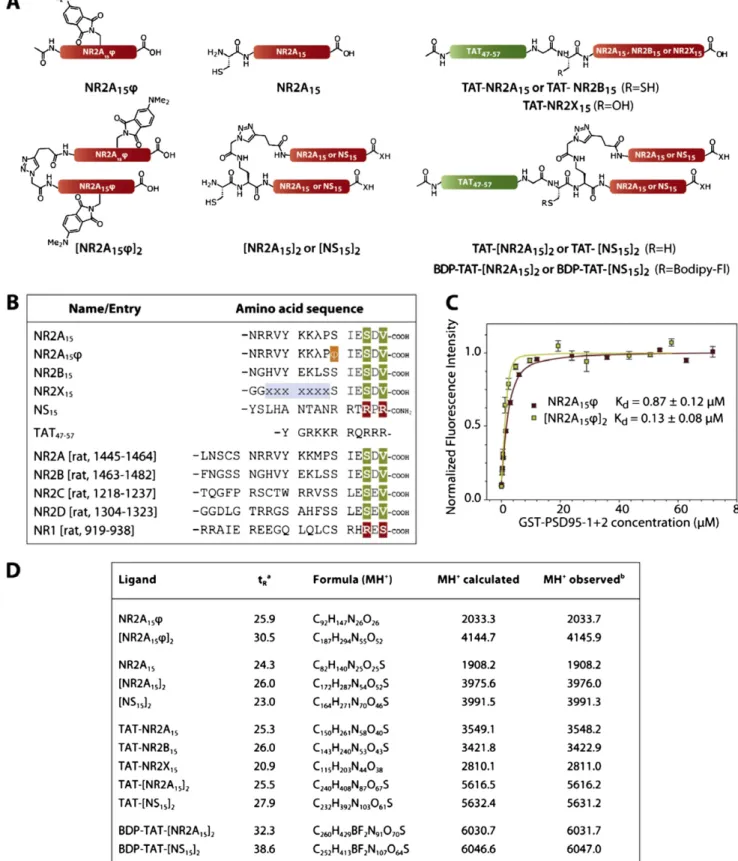
2B-NMDAR surface diffusion [measured in young hippocampal cultured neurons that do not express 2A-NMDAR (25)]. Strik-ingly, TAT-[NR2A15]2had no effect on the surface diffusion of
native 2B-NMDARs (Fig. 2A). The diffusion coefficient dis-tributions were superimposed, indicating that TAT-[NR2A15]2
did not affect the anchoring of 2B-NMDARs. Because the syn-aptic anchoring of 2B-NMDARs may not depend on the in-teraction with PDZ proteins, we compared the surface trafficking of NR2B WT (2B-WT) and a NR2B mutant (i.e., 2B-S1480A), which does not coimmunoprecipitate with PSD-95 (9). The dif-fusion coefficient was significantly higher for 2B-S1480A (me-dian of 0.57μm2/s, IQR of 0.24–1.08 μm2/s,n = 694 trajectories) than for 2B-WT (median of 0.32μm2/s, IQR of 0.13–0.69 μm2/s,
n = 344 trajectories) and the synaptic dwell time, defined as the mean time spent by a mobile receptor in the synaptic area, was significantly higher for 2B-WT compared with 2B-S1480A (Fig. 2C). Both 2B-WT and 2B-S1480A were confined within the synapse but to a significantly lower degree for 2B-S1480A (Fig. 2D). These results demonstrate that the synaptic retention of surface 2B-NMDARs is dynamically regulated by the interaction with PDZ proteins (9) and insensitive to TAT-[NR2A15]2ligand.
In addition, incubating the neurons with TAT-NR2B15(5μM, 10
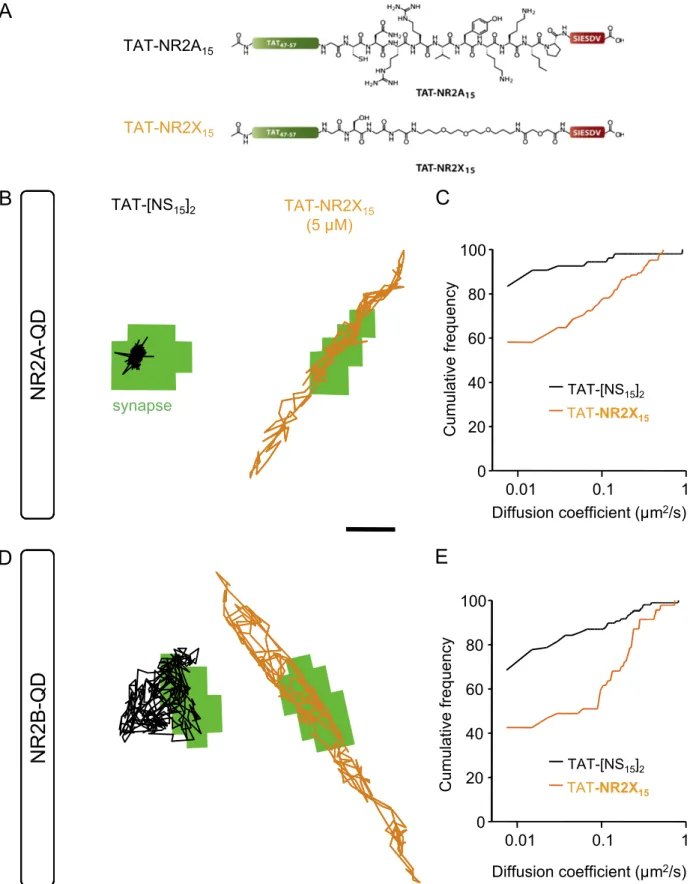
min) that mimics the last 15 aa of the NR2B subunit C terminus increased the surface diffusion of synaptic 2B-NMDARs without affecting the one of 2A-NMDARs (Fig. S4), consistent with previous biochemical reports using similar ligands (23, 24). Fi-nally, a monovalent ligand containing only the last 6 aa of the NR2 C-terminus sequence (TAT-NR2X15), which is identical for
NR2A and NR2B subunits, increased the surface diffusion of both 2A- and 2B-NMDARs (Fig. S5), indicating that the PDZ binding sequence (last few amino acids of the C terminus) is indeed necessary to anchor the receptor in the synapse, and upstream amino acid sequence(s) provide a specificity motif for NR2 subunit.
To further test the specificity of the ligand, we then reasoned that if TAT-[NR2A15]2competes specifically against the NR2A
synapse WT 2B-WT Mutant2B-S1480A 0 0.05 0.1 0 1 2 3 2B-S1480A 2B-WT Time (s) MSD (µ m 2) 0.0 0.2 0.4 0.6 0.8
**
Sy n. dwell tim e (s ) t0 t40s t0 t40s TAT-[NS15]2 TAT-[NR2A15]2 0 0.1 0 0.05 0.1 0.4 0.7 0.2 0.4 0.6 Frequenc y (relative ) 0.01 0.1 1 0 0.5 1 C u m u l. f req. Diff. Coef. (µm2/s) Diffusion coefficient (µm2/s) 2B-NMDAR 0 1.0 2.0**
Sy n. dwell tim e (s ) 0 0.05 0.1 0 0.5 1 - TAT-[NR2A15]2 + TAT-[NR2A15]22A-S1462A diff. coef. (µm2/s) P>0.05 KS test Frequenc y (relative )
A
B
C
D
E
F
Fig. 2. TAT-[NR2A15]2 incubation does not affect native 2B-NMDARs or
NR2A subunit mutant that does not bind to PDZ proteins. (A) Native 2B-NMDARs were tracked using QDs coupled to antibodies directed against an extracellular epitope of endogenous NR2B subunit in presence of TAT-[NS15]2
(n = 675 trajectories) or TAT-[NR2A15]2(n = 442 trajectories). The frequency
distribution (cumulative; Upper Right) of diffusion coefficients revealed that TAT-[NR2A15]2did not significantly affect the diffusion of native 2B-NMDARs
(P> 0.05, Mann–Whitney test). (B) The surface diffusion of recombinant 2B-NMDARs was assessed using recombinantflag-tagged NR2B subunits: WT (2B-WT) or mutant form (2B-S1480A) that does not bind PDZ proteins. These subunits were tracked using anti-Flag coupled QDs. Representative 40-s trajectories of anti-Flag QDs tracking 2B-WT (Left) or 2B-S1480A (Right). (Scale bar: 500 nm.) The starting and ending point are referred as t0and t40s,
respectively. The green areas correspond to synapses. (C) Synaptic dwell time was measured for NMDARs containing WT (n = 235 trajectories) or
S1480A (n = 532 trajectories). Note the reduction in the time spent by 2B-S1480A within the synapse (**P< 0.01, t test). (D) Plot of the mean square displacement (MSD) versus time for synaptic receptors containing the subunit 2B-WT or 2B-S1480A. The curves exhibit a negative curvature characteristic of a confined behavior. Note the higher degree of confinement for 2B-WT subunits. (E) Synaptic dwell time was measured for 2A-NMDARs containing either 2A-WT (n = 487 trajectories) or mutant 2A-S1462A (n = 474 trajecto-ries). Note the reduction in the time spent by the mutant within the synapse (**P< 0.01, t test). (F) The frequency distribution of diffusion coefficients of mutant 2A-S1462A in absence (n = 58 trajectories; black squares) or presence of TAT-[NR2A15]2(n = 94 trajectories; gray squares). No significant difference
was observed (P> 0.05, Mann–Whitney test).
Bard et al. PNAS Early Edition | 3 of 6
NEUROSCI
C-terminal domain for the binding to its PDZ proteins, it would have no additional effect on the diffusion of 2A-NMDARs containing a 2A-S1462A mutation in the C terminus that pre-vents NR2A/PSD-95 coimmunoprecipitation (co-IP) (9). The 2A-S1462A displayed a threefold higher surface diffusion (P < 0.001) and 1.4-fold shorter dwell time (Fig. 2E) than WT 2A-NMDARs. TAT-[NR2A15]2did not increase the surface
diffu-sion of 2A-S1462A (Fig. 2F), indicating that the TAT-[NR2A15]2-induced increase in 2A-NMDAR surface diffusion
was occluded by the S146induced increase in 2A-NMDAR surface diffusion. We then tested the specificity of TAT-[NR2A15]2 on other membrane proteins by imaging the
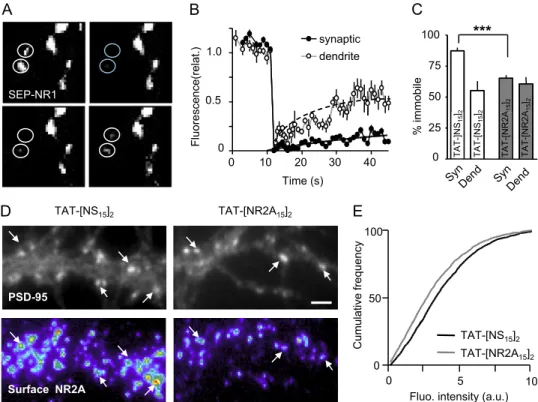
surface trafficking of the native potassium channel Kv1.3, en-dogenously expressed in hippocampal neurons (31), because its C terminus contains a PDZ binding site similar to those of NR2A and NR2B subunits (Fig. S6A) (32). Remarkably, TAT-[NR2A15]2 produced no change in the diffusion pattern, the
mobile fraction, the global diffusion coefficient, or the surface distribution of Kv1.3 channels (Fig. S6 B–E). Thus, although NR2A and Kv1.3 channel bind PDZ site with similar affinities (32) and exhibit a high similarity in their C-terminus amino acid sequence, TAT-[NR2A15]2 specifically acts on surface
2A-NMDARs and not on surface Kv1.3 channel. In addition, we report that TAT-[NR2A15]2 does not impact on the surface
GABAAreceptor (Fig. S6F), which is not anchored in synapse by
a PDZ domain-binding motif (33).
Interaction Between NR2A Subunits and PSD-95 Is Specifically
Disrupted by the NR2A Ligand.Because the PDZ-containing
scaf-folding proteins change during development, i.e., PSD-95 is the dominant scaffolding protein in mature neurons and SAP102 is the dominant scaffolding protein in immature neurons, the possibility that the TAT-[NR2A15]2ligand better interacts with certain PDZ
proteins remains to be tested. For this, wefirst measured, from forebrain homogenates, the impact of the NR2A ligand on the interaction between PSD-95 and NR2A or NR2B subunits using co-IP. The PSD-95/2A subunit interaction was specifically affected by the ligand whereas the PSD-95/2B subunit interaction remains unaffected (Fig. S7). Furthermore, we used a heterologous cell system to further determine the impact of the ligand on the in-teraction between NR2 subunit and the most abundant PDZ proteins, PSD-95. In heterologous cells, 2A- and 2B-NMDARs coimmunoprecipitate with the four PSD-95 MAGUK family of
scaffolding proteins (34). In addition, PSD-95 enhances 2A- and 2B-NMDAR cell surface expression through a process that requires the NR2 C terminus sequence -ESDV (34). To test the specificity of TAT-[NR2A15]2on 2A- and 2B-NMDAR
traffick-ing, we then measured the effect of PSD-95 on cell surface 2A- or 2B-NMDAR expression, as previously described (34). We first observed that either TAT-[NS15]2or TAT-[NR2A15]2had no
ef-fect per se on the basal expression of the subunit (Fig. 3). PSD-95 enhanced the cell surface expression of both 2A- and 2B-NMDARs (Fig. 3). The incubation with TAT-[NR2A15]2
com-pletely blocked the PSD-95-induced 2A-NMDAR surface ex-pression, whereas TAT-[NR2A15]2 had no effect on
PSD-95-induced 2B-NMDAR surface expression (Fig. 3). In all conditions, TAT-[NS15]2incubation had no significant effect on the
PSD-95-induced NR2-NMDAR surface expression. All together, these data demonstrate, in neuronal and heterologous systems, that the TAT-[NR2A15]2divalent ligand specifically blocks the interaction
between 2A-NMDARs (no effect on 2B-NMDARs) and the most abundant protein of the postsynaptic density, PSD-95.
Rapid Redistribution of 2A- and 2B-NMDARs in Excitatory Synapses.
As TAT-[NR2A15]2 specifically destabilizes synaptic
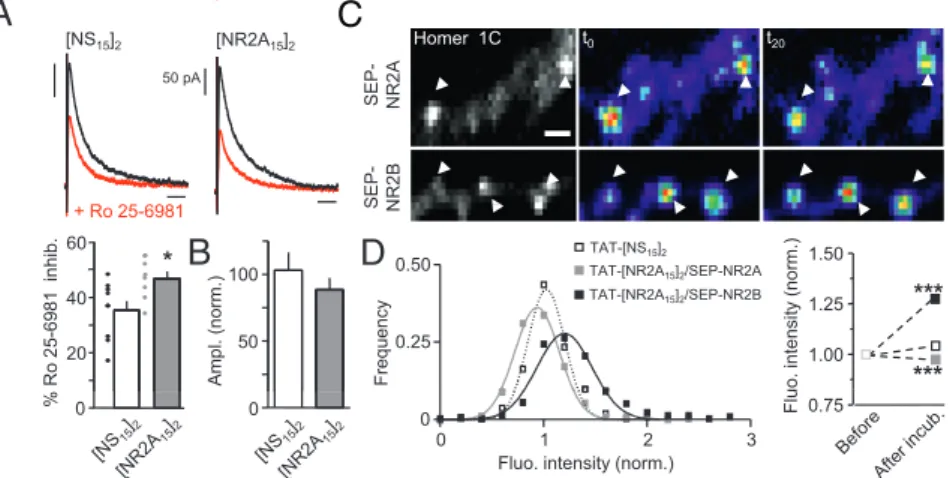
2A-NMDARs, we investigated the functional consequences of such an effect by first measuring NMDAR-mediated synaptic cur-rents. We report that, in the presence of [NR2A15]2(within
re-cording pipette), the kinetics of NMDAR miniature excitatory postsynaptic currents (mEPSCs) were significantly increased whereas AMPAR mEPSC remained unchanged (Fig. S8). We then recorded evoked NMDA excitatory postsynaptic currents (eEPSC) from CA1 pyramidal neurons (P16–20; Fig. 4A) and found that a 15- to 20-min dialysis of [NR2A15]2 significantly
increased the Ro 25-6981 (NR2B subunit antagonist, 1 μM)-in-duced inhibition of NMDAR current (Fig. 4A), consistent with an increased contribution of 2B-NMDARs to synaptic currents. Interestingly, the amplitude of both mEPSCs and eEPSCs remained unchanged in the presence of [NR2A15]2, indicating
that the [NR2A15]2-induced removal of 2A-NMDARs from
synapse was compensated by the insertion of other NMDARs with slower kinetics (Fig. 4B). To gain insight in the NR2-NMDAR trafficking at identified synapses, the fluorescence in-tensity of surface NR2A (SEP-NR2A) and NR2B (SEP-NR2B) subunits was measured over time before and after incubation with TAT-[NS15]2 or TAT-[NR2A15]2 (Fig. 4C). Synaptic and
extrasynaptic NMDARs were distinguished by coexpressing the synaptic marker Homer 1C-DsRed. First, the intensity of extra-synaptic surface NMDARs (SEP-NR1, SEP-NR2A, SEP-NR2B) was not significantly altered by the presence of TAT-[NS15]2or
TAT-[NR2A15]2. Within synapses, the intensity of 2A-NMDAR
clusters was decreased as demonstrated by the significant left shift of the cluster distributions (P < 0.001; Fig. 4D) or by the significant decrease of average values (Fig. 4E). Surprisingly, under the same conditions the intensity of 2B-NMDAR synaptic clusters was increased significantly (e.g., right shift of the distri-bution;P < 0.001; Fig. 4 D and E). The effect was observed 15 to 20 min after the ligand incubation and was stable over time. In such mature synapses, TAT-[NR2A15]2reduced the surface
dif-fusion of synaptic 2B-NMDARs and increased their synaptic dwell time, consistent with a higher retention of these receptors. These data demonstrate thus that 2A- and 2B-NMDARs rapidly redistribute within synaptic areas. In addition, displacing NMDARs out of synapses by preventing the interaction of 2A-NMDARs and PDZ proteins is compensated by the increase contribution of other NMDAR subtypes, indicating that an un-expected level of specificity between NR2 subunits and PDZ proteins is present in postsynaptic densities.
1.5 y +TAT-[NS 15 ] 2 +TAT-[NR2A 15 ] 2 Buffer 0 0.5 1.0 S t i v i t c a e r o n u m m i c i f i c e p 0 S
Fig. 3. TAT-[NR2A15]2specifically blocks the interaction between PSD-95
and NR2A subunit. HEK 293 cells were cotransfected in triplicate with NR1-1a/NR2A or NR1-1a/NR2B with or without PSD-95 and cell surface expressed NMDARs measured by ELISA using either anti-NR2A 44–58 Cys or anti-NR2B 46–60 Cys affinity-purified antibodies. The results are expressed as the ratio of absorbance and expressed as the means ± SEM (n = 2 independent transfections for each combination). As previously shown, PSD-95 enhanced cell surface delivery of 2A- and 2B-NMDARs. These effects were then ex-amined after incubation with TAT-[NS15]2 (10 μM; open bar) or
TAT-[NR2A15]2(10μM; gray bar). Note that PSD-95 failed to increase surface
2A-NMDAR expression in the presence of TAT-[NR2A15]2.
Discussion
Although synaptic NR2-NMDARs play a key role in synaptic re-finement (2), the molecular mechanisms as well as the dynamics that govern their surface distribution and rapid trafficking are largely unknown. To shed new light on this issue, we developed a biomimetic divalent ligand that acutely and efficiently blocks the interaction between PDZ proteins and native 2A-NMDARs (enriched at synapses). We unravel an unexpected role of the divalent arrangement of the NR2 subunits in providing efficient anchoring within synapses and strengthen the need to dynamically study such interactions in native conditions. Indeed, by using mono- or divalent ligands, we now identified that the binding ef-ficacy is highly dependent on the divalent structure of the 2A-NMDAR complex, and the specific binding of 2A-2A-NMDAR (ver-sus 2B-NMDAR for instance) relies on amino acid sequence(s) upstream to the C terminus, whereas the last C terminus amino acids are implicated in the direct binding to PDZ scaffold proteins (Fig. S9). Thus, NR2 subunits associate with PDZ proteins via a C-terminal 4-aa sequence (7–10), which is identical in NR2A and NR2B subunits, and other upstream amino acids that are within 15 aa of the C terminus, and as previously proposed, in more upstream sequences (29). Although the binding mechanism of these do-mains redo-mains poorly understood (35), it suggests that 2A- or 2B-NMDARs are engaged in different sets of interactions within the scaffold environment. Consistently, the NR2A-ligand-induced rapid exit of 2A-NMDARs from postsynaptic densities was paral-leled by a compensatory increase in 2B-NMDAR content, in-dicating that the 2A/2B-NMDAR synaptic ratio is dynamically regulated. Functionally, long-term potentiation of hippocampal
synapses has been associated with a rapid change in the synaptic content in 2A and 2B-NMDARs (36), consistent with a dynamic redistribution of surface 2A- and 2B-NMDARs around the synaptic area. Thus, understanding the rules that govern NR2-NMDAR surface distribution and, most importantly, their dynamic retention in the postsynaptic density will surely shed new lights on the nanodomain organization of NMDARs and the fine tuning of NMDAR-dependent forms of synaptic adaptations in physiological and pathological paradigms.
Materials and Methods
Complete discussions of ligand synthesis, cell culture, immunocytochemistry, synaptic live cell staining, protein expression, single particle (QD) tracking, fluorescence recovery after photobleaching (FRAP), electrophysiology, im-munoprecipitation, and in vitro cell surface assays are in SI Materials and Methods. The transduction and cell distribution of the ligand are de-tailed in Fig. S10. The impact of the ligand and its vehicle on receptor trafficking is detailed inFig. S11.
ACKNOWLEDGMENTS. We thank Laurent Ladépêche, Beatrice Tessier, Arnaud Frouin, and Christophe Blanchet for technical assistance; Robert Wenthold (National Institutes of Health, Bethesda, MD) for providing NR2 cDNA plasmids and constructive discussions; and Antoine Triller [Ecole Nor-male Superieure (ENS), Paris] for providing antibody. This work was sup-ported by Centre National de la Recherche Scientifique/Agence Nationale de la Recherche Grant JC08_329238 (to L.G.), Chem-Traffic (M.S. and D.C.), Human Frontier Science Program Grant MRGP0007/2006-C (to B.I., M.S., and D.C.), Fondation pour la Recherche Médicale (L.B., D.C.,and L.G.), Conseil Régional d’Aquitaine, Marie Curie postdoctoral fellowship (PICK-CPP to M.S.), Ministère de l’Enseignement Supérieur et de la Recherche, European Re-search Council Advanced ReRe-search Grant Nano-Dyn-Syn (to D.C.), and the UK Biotechnology and Biological Sciences Research Council (F.A.S.).
1. Cull-Candy SG, Leszkiewicz DN (2004) Role of distinct NMDA receptor subtypes at central synapses. Sci STKE 2004:re16.
2. Yashiro K, Philpot BD (2008) Regulation of NMDA receptor subunit expression and its implications for LTD, LTP, and metaplasticity. Neuropharmacology 55:1081– 1094.
3. Chen BS, Roche KW (2007) Regulation of NMDA receptors by phosphorylation. Neuropharmacology 53:362–368.
4. Lau CG, Zukin RS (2007) NMDA receptor trafficking in synaptic plasticity and neuropsychiatric disorders. Nat Rev Neurosci 8:413–426.
5. Pérez-Otaño I, Ehlers MD (2005) Homeostatic plasticity and NMDA receptor trafficking. Trends Neurosci 28:229–238.
6. Wenthold RJ, Prybylowski K, Standley S, Sans N, Petralia RS (2003) Trafficking of NMDA receptors. Annu Rev Pharmacol Toxicol 43:335–358.
7. Kornau HC, Schenker LT, Kennedy MB, Seeburg PH (1995) Domain interaction between NMDA receptor subunits and the postsynaptic density protein PSD-95. Science 269:1737–1740.
8. Mohrmann R, Köhr G, Hatt H, Sprengel R, Gottmann K (2002) Deletion of the C-terminal domain of the NR2B subunit alters channel properties and synaptic targeting 50 pA [NS 15 ] 2 [NR2A 15 ] 2
A
D
TAT-[NR2A 15 ] 2 /SEP-NR2A 0.25 0.50 TAT-[NS 15 ] 2 y c n e u q e r F u ) . m r o n ( y t i s n e t n i . o TAT-[NR2A 15 ] 2 /SEP-NR2B + Ro 25-6981 50 100 ) . m r o n ( . l p m A . b i h n i 1 8 9 6 - 5 2 o R 20 40 60 *B
1.00 1.25 1.50 *** *** 0 0 1 2 3Fluo. intensity (norm.)
l F u 0 % 0 0.75 Homer 1C t 0 t 20 - P E S A 2 R N S - P E N B 2 R S N
C
Fig. 4. Dynamic regulation of surface 2A- and 2B-NMDAR content in postsynaptic areas. (A) Evoked NMDAR EPSCs (recorded at +30 mV) averaged at 0 to 2 min (black trace) or 18 to 20 min (red trace) after dialysis with Ro 25-6981 (1μM, 2B-NMDAR antagonist; Left). (Horizontal scale bar: 100 ms.) The Ro 25-6981 incubation significantly reduced the NMDAR eEPSC amplitude (Right). In te presence of [NS15]2(n = 9 neurons) or [NR2A15]2(n = 8 neurons), Ro 25-6981
reduced the NMDAR eEPSC amplitude by 35% and 47%, respectively. (B) The amplitude (normalized) of NMDAR eEPSCs remained stable in presence of [NR2A15]2ligand ([NS15]2, n = 6 neurons; [NR2A15]2, n = 7 neurons). (C) Thefluorescence intensity of synaptic SEP-NR2A (Upper) and SEP-NR2B (Lower) clusters
colocalized with Homer 1C was followed over a period of 20 min after acute addition of 5μM of TAT-[NR2A15]2(Scale bar: 1μm.) (D) Left: Frequency
dis-tribution of thefluorescence intensity of SEP-NR2A and SEP-NR2B clusters after 20 min in the presence of TAT-[NS15]2or TAT-[NR2A15]2. The Gaussianfit is
centered on 1 after incubation with TAT-[NS15]2(n = 563 clusters; dashed black line) indicating that the receptor content within the cluster did not change
over time. Note the shift of the curve toward the left for SEP-NR2A (n = 451 clusters; solid gray line) and toward the right for SEP-NR2B (n = 309 clusters; full black line) after the 20 min incubation with TAT-[NR2A15]2showing, respectively, a decrease and an increase in the receptor content. Right: Normalized mean
fluorescence intensity of the clusters before and after a 20-min incubation with TAT-[NS15]2or TAT-[NR2A15]2(P< 0.001, paired t test).
Bard et al. PNAS Early Edition | 5 of 6
NEUROSCI
of N-methyl-D-aspartate receptors in nascent neocortical synapses. J Neurosci Res 68: 265–275.
9. Prybylowski K, et al. (2005) The synaptic localization of NR2B-containing NMDA receptors is controlled by interactions with PDZ proteins and AP-2. Neuron 47: 845–857.
10. Cui H, et al. (2007) PDZ protein interactions underlying NMDA receptor-mediated excitotoxicity and neuroprotection by PSD-95 inhibitors. J Neurosci 27:9901–9915. 11. Barria A, Malinow R (2002) Subunit-specific NMDA receptor trafficking to synapses.
Neuron 35:345–353.
12. Chung HJ, Huang YH, Lau LF, Huganir RL (2004) Regulation of the NMDA receptor complex and trafficking by activity-dependent phosphorylation of the NR2B subunit PDZ ligand. J Neurosci 24:10248–10259.
13. Mori H, et al. (1998) Role of the carboxy-terminal region of the GluR epsilon2 subunit in synaptic localization of the NMDA receptor channel. Neuron 21:571–580. 14. Sprengel R, et al. (1998) Importance of the intracellular domain of NR2 subunits for
NMDA receptor function in vivo. Cell 92:279–289.
15. Thomas CG, Miller AJ, Westbrook GL (2006) Synaptic and extrasynaptic NMDA receptor NR2 subunits in cultured hippocampal neurons. J Neurophysiol 95: 1727–1734.
16. Steigerwald F, et al. (2000) C-Terminal truncation of NR2A subunits impairs synaptic but not extrasynaptic localization of NMDA receptors. J Neurosci 20:4573–4581. 17. Köhr G, et al. (2003) Intracellular domains of NMDA receptor subtypes are
determinants for long-term potentiation induction. J Neurosci 23:10791–10799. 18. Neyton J, Paoletti P (2006) Relating NMDA receptor function to receptor subunit
composition: Limitations of the pharmacological approach. J Neurosci 26:1331–1333. 19. Aarts M, et al. (2002) Treatment of ischemic brain damage by perturbing NMDA
receptor- PSD-95 protein interactions. Science 298:846–850.
20. Gardoni F, et al. (2006) A critical interaction between NR2B and MAGUK in L-DOPA induced dyskinesia. J Neurosci 26:2914–2922.
21. Lim IA, Merrill MA, Chen Y, Hell JW (2003) Disruption of the NMDA receptor-PSD-95 interaction in hippocampal neurons with no obvious physiological short-term effect. Neuropharmacology 45:738–754.
22. Stephenson FA, Cousins SL, Kenny AV (2008) Assembly and forward trafficking of NMDA receptors (review). Mol Membr Biol 25:311–320.
23. Sainlos M, Iskenderian WS, Imperiali B (2009) A general screening strategy for peptide-basedfluorogenic ligands: Probes for dynamic studies of PDZ domain-mediated interactions. J Am Chem Soc 131:6680–6682.
24. Groc L, et al. (2007) Surface trafficking of neurotransmitter receptor: comparison between single-molecule/quantum dot strategies. J Neurosci 27:12433–12437. 25. Groc L, et al. (2006) NMDA receptor surface mobility depends on NR2A-2B subunits.
Proc Natl Acad Sci USA 103:18769–18774.
26. Tovar KR, Westbrook GL (2002) Mobile NMDA receptors at hippocampal synapses. Neuron 34:255–264.
27. Zhao J, et al. (2008) Synaptic metaplasticity through NMDA receptor lateral diffusion. J Neurosci 28:3060–3070.
28. Tovar KR, Westbrook GL (1999) The incorporation of NMDA receptors with a distinct subunit composition at nascent hippocampal synapses in vitro. J Neurosci 19: 4180–4188.
29. Cousins SL, Kenny AV, Stephenson FA (2009) Delineation of additional PSD-95 binding domains within NMDA receptor NR2 subunits reveals differences between NR2A/PSD-95 and NR2B/PSD-NR2A/PSD-95 association. Neuroscience 158:89–NR2A/PSD-95.
30. Pegan S, et al. (2007) NMR studies of interactions between C-terminal tail of Kir2.1 channel and PDZ1,2 domains of PSD95. Biochemistry 46:5315–5322.
31. Grosse G, et al. (2000) Expression of Kv1 potassium channels in mouse hippocampal primary cultures: Development and activity-dependent regulation. J Neurosci 20: 1869–1882.
32. Lim IA, Hall DD, Hell JW (2002) Selectivity and promiscuity of thefirst and second PDZ domains of PSD-95 and synapse-associated protein 102. J Biol Chem 277:21697–21711. 33. Fritschy JM, Harvey RJ, Schwarz G (2008) Gephyrin: Where do we stand, where do we
go? Trends Neurosci 31:257–264.
34. Cousins SL, Papadakis M, Rutter AR, Stephenson FA (2008) Differential interaction of NMDA receptor subtypes with the post-synaptic density-95 family of membrane associated guanylate kinase proteins. J Neurochem 104:903–913.
35. Ryan TJ, Emes RD, Grant SG, Komiyama NH (2008) Evolution of NMDA receptor cytoplasmic interaction domains: implications for organisation of synaptic signalling complexes. BMC Neurosci 9:6.
36. Bellone C, Nicoll RA (2007) Rapid bidirectional switching of synaptic NMDA receptors. Neuron 55:779–785.
Supporting Information
Bard et al. 10.1073/pnas.1002690107
SI Materials and Methods
Ligand Synthesis and in Vitro Characterization. The design and
synthesis (M.S., B.I.) of peptide-based ligands is schematically described in this paragraph. Briefly, the last 15 residues of the ligand binding motifs werefirst assembled by standard Fmoc-based solid phase peptide synthesis and capped with a mixture of azide-and alkyne-derived acids. The ligation was conducted by click chemistry (1) on resin. For the 4-DMAP-containing ligand, the fluorophore was inserted post ligation on resin. The TAT se-quence was coupled as a C-terminal thioester to the purified li-gand N-terminal Cys residue by native chemical ligation. BODIPY-FL was coupled using a maleimide derivative to the same Cys residue. Recombinant PSD-95 tandem PDZ domains expression andfluorescence titrations were performed as pre-viously described (2).
Single Particle (QD) Tracking and Surface Diffusion Calculation.QD
655 Goat F(ab′)2 anti-Rabbit or anti-mouse IgG (Invitrogen) werefirst incubated for 30 min with the polyclonal antibodies against NR2A (1μg) and NR2B subunits (1 μg), the monoclonal anti-Flag (Stratagene), the anti GABAAα2 (gift from A. Triller,
Ecole Normale Superieure, Paris), and the Kv1.3 antibodies (Alomone Labs; epitope location corresponds to thefirst extra-cellular loop between domains S1 and S2 amino acid, i.e., residues 263–276 of Kv1.3). For this Kv1.3 antibody, specificity control has been obtained only with Western blot (Alomone Labs). Non-specific binding was blocked by additional casein (Vector Labo-ratories) to the QD 15 min before use. For experiments using TAT peptides, neurons werefirst incubated for 10 min at 37 °C in culture medium with precoated QDs (final dilution 1:2,000 for anti-NR2A and anti-NR2B coupled QDs, 1:10,000 for anti-GA-BAAcoupled QDs, 1:5,000 for anti-kv1.3 coupled QDs), then
for 1 min with 20 nM Green Mitotracker (Molecular Probes), for 5 min with 50μM pyrene butyrate (3) and finally for 10 min with 5μM TAT-[NS15]2or TAT-[NR2A15]2(or TAT-NR2B15when
specified).
We first examined the penetration of the TAT-[NR2A15]2 in
neurons. As shown inFig. S10, the ligand efficiently penetrates
cul-tured neurons and was observed in all dendritic compartments, in-cluding postsynaptic densities. We next tested the nonspecific effects of pyrene butyrate or TAT-[NS15]2on the mobility of 2A-NMDARs
(Fig. S11). Whereas pyrene butyrate had no effect per se, TAT-[NS15]2increased the proportion of immobile receptors.
Impor-tantly, such effect of TAT-[NS15]2was observed on 2B-NMDARs,
GluR2-AMPARs, GABAARs, and reproduced using the
mono-meric TAT-[NS15]. This indicates that TAT-[NS15]2slightly reduced
the surface trafficking of neurotransmitter receptors, irrespective of the nature of the receptors or the structure of the TAT ligand.
For single particle tracking of Flag-tagged NR2B subunits, neu-rons were incubated for 10 min with precoated anti-Flag QDs (final dilution, 1:10,000). QDs were detected by using a mercury lamp and appropriateexcitation/emissionfilters.Imageswereobtainedwithan integration time of 5 to 30 ms with up to 2,000 consecutive frames. Signals were detected using a CCD camera (Quantem; Roper Sci-entific). QDs were followed on randomly selected dendritic regions for up to 20 min. QD recording sessions were processed with Met-amorph software (Universal Imaging). The instantaneous diffusion coefficient, D, was calculated for each trajectory, from linear fits of thefirst four points of the MSD versus time function:
MSDðtÞ ¼ < r2> ðtÞ ¼ 4Dt [S1]
Synaptic dwell time was calculated for exchanging receptors and defined as the mean time spent within the synaptic area. The 2D trajectories of single molecules in the plane of focus were con-structed by correlation analysis between consecutive images using a Vogel algorithm.
Cell Culture, Immunocytochemistry, Synaptic Live Staining, and
Protein Expression. Cultures of hippocampal neurons were
pre-pared from E18 Sprague–Dawley rats following a previously de-scribed method (4–6). Briefly, cells were plated at a density of 100 to 200× 103cells per milliliter on poly-lysine precoated coverslips. Cultures were maintained in serum-free neurobasal medium (Invitrogen) and kept at 37 °C in 5% CO2for 20 d in vitro at
maximum. For immunostaining, surface 2A-NMDARs were specifically stained using a polyclonal NR2A subunit anti-body (1:100; F. A. Stephenson, London, United Kingdom) for 15 min on live neurons at 37 °C in culture medium. The specificity of the antibody was previously described (5). Briefly, neurons were then fixed with 4% paraformaldehyde/4% sucrose for 15 min, washed, and incubated with secondary antibodies anti-rabbit Alexa 488 antibodies (1:1,000, 45 min; Molecular Probes). To label postsynaptic areas, neurons were permeabilized using 0.1% Triton X-100, incubated with a primary rabbit polyclonal anti-shank antibody (1:750, 45 min; Abcam), and finally incu-bated with secondary antibody anti-rabbit Alexa 568 antibodies (1:1,500, 30 min; Molecular Probes). Neurons were washed and mounted and preparations were kept at 4 °C until quantification. For the quantification of surface NR2A staining within individual shank cluster, the shank-synaptic staining served as a maskfilter to isolate surface NR2A staining in individual shank clusters. The integratedfluorescence level over the shank-cluster area was then measured for each cluster. Thefluorescence analysis was realized using imaging tools from Metamorph software (Universal Imag-ing). To label synapses live cultured neurons were incubated for 1 to 2 min at room temperature with 10 nM Mitotracker (Deep Red-Fluorescent Mitotracker; Molecular Probes) prior to imag-ing experiments. Neurons were transfected at 7 to 10 d in vitro with Homer1c-DsRed or Homer1c-GFP alone or with SEP-NR1, SEP-NR2A, or SEP-NR2B using the Effectene transfection kit (Qiagen). We mixed 2μg of DNA with 25 μL of Effectene and 8 μL of enhancer in 150 μL of reaction buffer, and then added the mixture to cultured neurons, which were transferred to serum-free neurobasal medium 10 min beforehand. After an incubation period of 45 min, neurons were placed in the old medium again.
Postsynaptic Localization of NMDARs. Surface diffusion of
post-synaptic NMDARs has been described using electrophysiological and high-resolution imaging approaches (5, 7–9). It is noteworthy that the diffusion of presynaptic NMDARs has also been de-scribed in synapses from the entorhinal cortex at early stages of development (10). Because single particle tracking on endogenous NMDARs does not distinguish between pre- and postsynaptic receptors, we investigated whether presynaptic tagged NMDARs were present in our hippocampal system. However, we found no evidence for surface NMDARs in presynaptic (VAMP-2-positive cluster) terminals, indicating that in our neuronal preparation surface NMDARs are mostly located in the postsynaptic area.
Electrophysiology.Neurons were continuously perfused with
extra-cellular solution containing (in mM): 145 NaCl, 2.5 KCl, 10 Hepes, 10 glucose, 2 CaCl2, and 0.1 mM Mg2+, pH 7.3, osmolarity 300
mOsm/L. Patch pipettes (3–5 MΩ) were filled with (in mM): 140 cesium methanesulfonate, 2 MgCl2, 4 NaCl, 5 phosphocreatine, 10
Hepes, 2 Na2-ATP, 0.33 Na-GTP, 5 QX-314, and 0.2 EGTA, pH
7.3, osmolarity 290 mOsm/L. Recordings in voltage clamp mode were performed with an EPC 10 double patch-clamp amplifier (HEKA). Recordings included for analysis were collected during periods of stable series resistance. Data were acquired and stored using Pulse-Pulsefit software, version 8.62. Miniature NMDAR EPSCs were recorded at +40 mV and isolated in the presence of TTX (1μM), bicuculline (20 μM; Ascent Scientific), and NBQX (10μM, Ascent Scientific). Miniature AMPAR EPSCs were re-corded at−50 mV and isolated in the presence of TTX (1 μM), bicuculline (20μM; Ascent Scientific), and AP5 (25 μM; Ascent Scientific) (11). Miniature EPSCs were analyzed by using the Mini-Analysis program (version 6.0.7; Synaptosoft) and IGOR (Wave-Metrics). The limit of detection was greater than 5 pA. The decay times of NMDA-mEPSCs werefitted using two exponentials.
Hippocampal slices were prepared from P16-20 C57Bl6 mice. Mice were decapitated and the brain was removed and placed in ice-cold solution composed of (in mM): 87NaCl, 2.5 KCl,10 glucose, 75 sucrose, 25 NaHCO3, 1.25 NaH2PO4, 0.5 CaCl2, and 7 mM
Mg2+. Transverse hippocampal slices (350 μm) were cut using
a vibrating tissue slicer (Leica) and transferred to a holding chamber and stored at 33 °C. For recording, slices were individually transferred to a recording chamber where they were perfused at 30 °C to 35 °C. The extracellular solution contained (in mM): 125 NaCl, 2.5 KCl, 25 glucose, 26 NaHCO3, 1.25 NaH2PO4, 2 CaCl2,
and 1 mM Mg2. GABAAR postsynaptic currents were blocked by
20μM bicuculline. AMPAR EPSCs were blocked by 10 μM NBQX. CA1 pyramidal cells were visually identified using IR-DIC video-microscopy and whole-cell patch-clamp recordings were performed with an EPC-10 patch-clamp amplifier (HEKA). The pipette so-lution contained (in mM): 140 cesium methanesulfonate, 2 MgCl2, 4
NaCl, 5 phosphocreatine, 10 Hepes, 2 Na2-ATP, 0.33 Na-GTP, 5
QX-314, and 0.2 EGTA, pH 7.3, osmolarity 290 mOsm/L. Patch pipettes were pulled using a horizontal puller (Sutter Instruments) and their resistance was 3 to 5 MΩ. The series resistance, which was continuously monitored during the experiments using a 5-mV hy-perpolarizing pulse. Recordings included for analysis were collected during periods of stable series resistance. Recordings with series resistance greater than 20 MΩ were discarded. Responses were sampled at 10 kHz. A glass pipettefilled with extracellular solution (NaCl 0.9%) was used to evoke unitary NMDA EPSCs. The pipette was placed in the stratum radiatum of the CA1 area. Stimulations were applied at 0.05 Hz and the cells were held at +30 mV.
FRAP and Follow-Up of Cluster Fluorescence Intensity.SEP-NR1 and
Homer1c-DsRed cotransfected neurons were placed on the heated stage (37 °C) of an inverted confocal spinning-disk mi-croscope (Leica). To test the population of surface SEP-NR1, we used low-pH solution adjusted to pH 5.4, which quenched all of thefluorescence indicating that SEP allows the specific visuali-zation of surface receptors. Fluorescence was excited using a monochromator controlled by Metamorph software (Universal Imaging). To photobleach locally, we used a sapphire laser 488– 20 to 50% power to avoid photo damage. The laser was coupled to the microscope via a galvometric mirror, which allowed us to photobleach several regions within a short time window. Re-covery from photobleaching was monitored by two consecutive acquisition periods at 2 and 0.5 Hz acquisition rates, respectively. Recovery curves were corrected for continuous photobleaching and background noise. For the follow-up of clusterfluorescence intensity to assess receptor content, neurons were cotransfected
with Homer1c-DsRed and either SEP-NR2A or SEP-NR2B. Clusters were imaged over a period of 20 min after acute addi-tion of 5 μM TAT-[NS15]2 or TAT-[NR2A15]2. Fluorescence
intensity was measured using Metamorph software (Universal Imaging) and corrected for photobleaching and background noise.
Immunoprecipitation and Western Blot Analysis.A frozen adult rat
brain (approximately 1.6 g) was thawed in 16 mL ice cold SHC buffer (320 mM sucrose, 1 mM Hepes, 1 mM MgCl2, 1 mM
NaHCO3) containing a protease inhibitor mixture (1:1,000;
Calbiochem) for 5 min and cut into small pieces. The tissue was homogenized using a Teflon-glass homogenizer and the ho-mogenate was spun at 1,000× g for 10 min at 4 °C. The resulting supernatant was spun at 10,000 × g for 15 min to obtain a P2 crude membrane fraction. P2 pellets were divided into aliquots and stored at−80 °C until solubilization and immunoprecipita-tion reacimmunoprecipita-tions were performed.
P2 fractions were solubilized with RIA buffer containing 1% SDS (200 mM NaCl, 10 mM EDTA, 10 mM Na2HPO4, 0.5% Nonidet
P-40, 1% SDS) for 5 min at 4 °C, followed by dilution to 0.1% SDS. The resulting lysate (200μg of protein) was diluted in RIA buffer containing 0.1% SDS, followed by incubation under constant agi-tation at 37 °C for 15 min with ligand TAT-[NR2A15]2(1.5μM),
TAT-[NS15]2(1.5μM), or a similar volume of dH2O. NR2A (0.6μg,
AGC-002; Alomone Labs) or NR2B (0.8μg, polyclonal antibody described in ref. 7) antibodies were incubated under constant agi-tation at 37 °C for 15 min with 10μL of prewashed Protein A beads (Dynabeads Protein A; Invitrogen). The lysates were added to the antibody-bead mixtures and incubated under constant agitation overnight at 4 °C. Immunoprecipitates were separated by SDS/ PAGE and analyzed by Western blotting using a rabbit monoclonal antibody to NR2A (04-901, clone A12W; Millipore), a rabbit polyclonal antibody to NR2B (A-6474; Molecular Probes), or a mouse monoclonal antibody to PSD-95 (MA1-046; Thermo Sci-entific). Detection was performed using the SuperSignal West Femto Maximum Sensitivity Substrate detection kit (Pierce), re-vealed with a Chemigenius system (Syngene). Quantification of bands intensity was performed using Genetools software (Syngene).
In Vitro Cell Surface Assay.HEK 293 cells were cotransfected in
triplicate in 24-well tissue culture plates with either pCIS vector, NR1-1a/NR2A or NR1-1a/NR2B NMDA receptors in the pres-ence and abspres-ence of PSD-95 (2μg of total DNA per well). Post-transfection (24 h), cells were incubated with either vehicle con-trol, TAT-[NR2A15]2or control TAT-[NS15]2(10μM) for 1 h at 37
°C. Cell surface NMDA receptor expression was determined by ELISA with affinity-purified antibodies directed against extracel-lular epitopes of NR2A and NR2B, i.e., anti-NR2A 44–58 Cys (0.25μg/mL) or anti-NR2B 46–60 Cys (0.5 μg/mL).
Data and Statistical Analysis.The instantaneous diffusion coefficient
is reported as the median± 25% to 75% (i.e., IQR). All of the other group values are expressed as mean± SEM. Comparisons between groups for instantaneous diffusion coefficients were performed using Mann–Whitney test (pair comparison) or Kruskal-Wallis followed by Dunn multiple-comparison test (group comparison). All of the other comparisons between groups were performed using parametric statistical tests, Studentt test (pair comparison), ANOVA followed by Newman-–Keuls multiple comparison test (group comparison), or Kolmogorov–Smirnov test (distribution comparison). Signifi-cance levels were defined as P < 0.05, P < 0.01, and P < 0.001.
1. Rostovtsev VV, Green LG, Fokin VV, Sharpless KB (2002) A stepwise Huisgen cycloaddition process: Copper(I)-catalyzed regioselective“ligation” of azides and terminal alkynes. Angew Chem Int Ed Engl 41:2596–2599.
2. Sainlos M, Iskenderian WS, Imperiali B (2009) A general screening strategy for peptide-based fluorogenic ligands: Probes for dynamic studies of PDZ domain-mediated interactions. J Am Chem Soc 131:6680–6682.
3. Takeuchi T, et al. (2006) Direct and rapid cytosolic delivery using cell-penetrating peptides mediated by pyrenebutyrate. ACS Chem Biol 1:299–303.
4. Banker GA, Cowan WM (1977) Rat hippocampal neurons in dispersed cell culture. Brain Res 126:397–425.
5. Groc L, et al. (2004) Differential activity-dependent regulation of the lateral mobilities of AMPA and NMDA receptors. Nat Neurosci 7:695–696.
6. Tardin C, Cognet L, Bats C, Lounis B, Choquet D (2003) Direct imaging of lateral movements of AMPA receptors inside synapses. EMBO J 22:4656–4665.
7. Groc L, et al. (2006) NMDA receptor surface mobility depends on NR2A-2B subunits. Proc Natl Acad Sci USA 103:18769–18774.
8. Tovar KR, Westbrook GL (2002) Mobile NMDA receptors at hippocampal synapses. Neuron 34:255–264.
9. Zhao J, et al. (2008) Synaptic metaplasticity through NMDA receptor lateral diffusion. J Neurosci 28:3060–3070.
10. Yang J, Chamberlain SE, Woodhall GL, Jones RS (2008) Mobility of NMDA autoreceptors but not postsynaptic receptors at glutamate synapses in the rat entorhinal cortex. J Physiol 586:4905–4924.
11. Groc L, Gustafsson B, Hanse E (2002) Spontaneous unitary synaptic activity in CA1 pyramidal neurons during early postnatal development: Constant contribution of AMPA and NMDA receptors. J Neurosci 22:5552–5562.
Fig. S1. Structure and amino acid sequence of the competing ligands. (A) Structure of the monovalent and divalent ligands (X: O for NR2A15or NH for NS15).
(B) Sequences used for the peptide-based ligands of panel A and C-terminus residues of NMDAR subunits.φ, Dab(4-DMAP); λ, norleucine; x. . .x, PEG spacer (20 atoms; reference 01–63-0141; Novabiochem). Critical residues at positions 0 and −2 are highlighted (green for the common residues found in PSD-95 ligands, red otherwise). (C) Fluorescence titrations of the 4-DMAP-containing ligands with recombinant GST-PSD95-1+2. (D) Peptide-based ligands characterization. Purity was assessed by analytical reverse-phase HPLC (YMC C18, ODS-A 5/120, 250× 4.6 mm) using a standard gradient (5% acetonitrile containing 0.1% TFA for 5 min followed by 5–95% acetonitrile containing 0.1% TFA over 50 min in water containing 0.1% TFA at a flow rate of 1 mL min−1). All peptide-based
ligands were more than 95% pure as judged by analytical HPLC. [at
Rindicates retention time;bpeptide-based ligands identity was confirmed by MALDI-TOF MS
(Voyager; PerSeptive Biosystems) using DHB as a matrix in linear or reflective modes.]
B 75 100 v . q e r f e A TAT-[NS 15 ] 2 TAT-[NR2A 15 ] 2 Neurons > 15 d.i.v. C 0.002 0.02 0.2 2 25 50 Diffusion coefficient (μm 2 /s) i t a l u m u C v TAT-[NR2A 15 ] 2 (n = 40) TAT-[NS 15 ] 2 (n = 50) 4 D synapse R D Q - A 2 mobile 0 . 0 0.2 0.4 0.6 * t n e i c i f f e o c n o i s u f f i D m μ ( 2 ) s / 1 2 3 4 ) ² m μ ( D S M TAT- [ NR2 A15]2 TAT-[NS 15 ] 2 N R 0 0 0 0.1 0.2 0
Time lag (sec) [ 15 ]2 D 75 100 f . q e r ) s ( 2.0 E F
*
Q - B 2 R N 0.0 0.5 1.0 1.5 0 25 50 75 TAT-[NS] 2 (n = 43) TAT-[NR2A] 2 (n = 84) NR2B diffusion coefficient (μ m 2 / s ) e v i t a l u m u C f P < 0.001 0.0 1.0 e m i t l l e w D (μ )Fig. S2. TAT-[NR2A15]2increases the mobility of synaptic 2A-NMDARs and decreases the mobility of 2B-NMDARs in mature (>15 d in vitro) synapses. (A)
Representative trajectories of synaptic QDs coupled to NR2A after 10 to 20 min of incubation with TAT-[NS15]2(Right) or TAT-[NR2A15]2(Left). The gray regions
correspond to synaptic sites labeled with Mitotracker. (Scale bar: 500 nm.) (B) Distribution of the diffusion coefficients of synaptic trajectories (bin size, 0.0075 μm2/s). Thefirst point of the curve corresponding to the proportion of immobile receptors shows the increase in mobility of 2A-NMDARs following incubation
with TAT-[NR2A15]2. (C) The diffusion coefficient of mobile synaptic receptors is significantly increased in the presence of TAT-[NR2A15]2(n = 13 trajectories/
group; *P< 0.05, Mann–Whitney test). (D) Plot of the MSD (in μm2) versus time lag (in s) for synaptic 2A-NMDARs in presence of TAT-[NS
15]2or TAT-[NR2A15]2.
(E) Distribution of the diffusion coefficients of NR2B synaptic trajectories. Note the significant shift toward lower values. (F) The synaptic 2B-NMDAR dwell time was significantly increased in presence of TAT-[NR2A15]2.
0.5 1.0 dendrite synaptic s ).t al er ( e c n e c 50 75 100 m eli b o m
***
A 5 1 ]2 A 5 1 ]2 2 2 SEP-NR1A
B
C
0 10 20 30 40 0 Time (s) er o ul F s 0 25 i % m 2 R N[ -T A T A 2 R N[ -T A T A S N] -T A T 5 1 ]2 S N[ -T A T 5 1 ]2E
D
TAT [NS ] TAT [NR2A ]PSD-95 TAT-[NR2A15]2 TAT-[NS15]2 50 100 u y c n e u q erf e vit al Surface NR2A 0 5 10 0 m u C u TAT-[NR2A15]2 TAT-[NS15]2
Fluo. intensity (a.u.)
Fig. S3. Acute disruption of the interaction between 2A-NMDARs and PDZ proteins increases the surface diffusion of surface NR1-NMDARs and decreases the synaptic content of NR2A subunits. (A and B) Clusters of SEP-NR1 were photobleached and the recovery of thefluorescence was followed over a period of 50 s and expressed as recovery versus time on synaptic and dendrite areas. Note the higher proportion of immobile receptors within the synaptic compartment compared with the dendrite. (C) The average immobile fraction of synaptic and dendritic NMDARs are quantified after incubation with TAT-[NS15]2(n = 52
synaptic clusters; n = 21 dendritic clusters) or TAT-[NR2A15]2(n = 61 synaptic clusters; n = 20 dendritic clusters). Note the specific decrease of the proportion of
immobile receptors within the synaptic area compared with the dendrite (***P< 0.001, t test). (D) Immunostaining of PSD-95 (synapse) and surface NR2A subunits in the presence of TAT-[NR2A15]2or TAT-[NS15]2. Arrowheads represent synapses, i.e., PSD-95 cluster. (Scale bar: 5μm.) (E) Fluorescence intensity of
surface NR2A subunits in synapses (TAT-[NR2A15]2, n = 1,684 synapses; TAT-[NS15]2, n = 1,981).
Fig. S4. TAT-NR2B15ligand specifically alters the surface diffusion of 2B-NMDARs without affecting the one of 2A-NMDARs. (A) Distribution of the diffusion
coefficients of synaptic surface NR2B-NMDARs in presence of the nonsense or TAT-NR2B15ligand (5μM, 10–15 min). Note the significant shift of the
dis-tributions in presence of TAT-NR2B15, consistent with an increase diffusion (nonsense, n = 57; TAT-NR2B15, n = 99 trajectories; P< 0.001). (B) Distribution of the
diffusion coefficients of synaptic surface NR2A-NMDARs in presence of the nonsense or TAT-NR2B15ligand (5μM, 10–15 min). No significant effect was
ob-served (nonsense, n = 71; TAT-NR2B15, n = 68 trajectories; P> 0.05).
A
B
TAT-NR2X
15TAT-NR2A
15D
TAT-NR2X
15(5 μM)
TAT-[NS
15]
2u
y
c
n
e
80
100
C
Q -
A
2
R
N
q
e r f
e
v i t
a l
u
m
u
C
u
0
20
40
60
TAT-[NS 15 ] 2 TAT -NR2X 15synapse
Diffusion coefficient (μm
2/s)
0.01
0.1
1
0
100
D
E
N
D
Q -
B
2
R
40
60
80
m
y
c
n
e
u
q
e r f
e
v i t
a l
u
TAT [NS ]N
0.01
0.1
1
0
20
u
C
m
Diffusion coefficient (μm
2/s)
TAT- [NS15 ]2 TAT -NR2X 15Fig. S5. TAT-NR2X15increases in a nonspecific manner the mobility of synaptic 2A- and 2B-NMDARs. (A) Comparison of TAT-NR2X15and TAT-NR2A15ligands.
The TAT-NR2X15ligand comprises the last six residues of the C terminus of both NR2A and NR2B subunits (−SIESDV), a linker composed of PEG and glycine
residues that provides a neutral backbone preventing any specific amino acid side chain interactions while maintaining the TAT sequence at a similar distance to that used for the other NR2A- and NR2B-derived ligands. (B) Representative trajectories of synaptic QDs coupled to NR2A after 10 to 20 min of incubation with TAT-[NS15]2(Left) or TAT-NR2X15(Right). The green regions correspond to synaptic sites labeled with Mitotracker. (Scale bar: 500 nm.) (C) Cumulative
distribution of the diffusion coefficients of synaptic trajectories (bin size, 0.0075 μm2/s). Thefirst point of the curve corresponds to the proportion of immobile
receptors. Note the significant increase mobility of 2A-NMDARs following incubation with TAT-NR2X15ligand (TAT-[NS15]2, n = 54; TAT-NR2X15, n = 105
trajectories; P< 0.001). (D) Representative trajectories of synaptic QDs coupled to NR2B after 10 to 20 min of incubation with TAT-[NS15]2(Left) or TAT-NR2X15
Legend continued on following page
(Right). The green regions correspond to synaptic sites labeled with Mitotracker. (Scale bar: 500 nm.) (E) Cumulative distribution of the diffusion coefficients of synaptic trajectories (bin size, 0.0075μm2/s). Thefirst point of the curve corresponds to the proportion of immobile receptors. Note the significant increase
mobility of 2B-NMDARs following incubation with TAT-NR2X15ligand (TAT-[NS15]2, n = 108; TAT-NR2X15, n = 47 trajectories; P< 0.01).
A B 0 2 0.4 0.6 0.8 1.0 K v1.3 MSD (μm²) TAT- [NS15]2
Name Sequence C-term
NR2A [rat] -LNSCS NRRVY KKMPS IE S D V -COOH
NR2B [rat] - FNGSS NGHVY EKLSS IE S D V -COOH
Kv1.3 [rat] - NPNSC VNIKK IF T D V -COOH
QD
CTTNN
0 0.50 1 1.5
0 0 . 2
Time lag (sec)
K 100 0 5 s ) TAT [NS15 ]2 TAT-[NR2A 15 ] 2 100 syn./peri extras y n. C-term C D E Kv1.3 0 20 40 60 80 Kv1.3 % mobile 0 0.1 0.2 0.3 0.4 0 . 5 K v1.3 diff. coef. (μm²/ s 0 20 40 60 80 100 K v1.3 surf. distribution (% of particule) y TAT-[NR2A 15 ] 2 TAT-[NS 15 ] 2 0 K 0 K 0 F α 2-GABA A QD QD-GABA A
Fig. S6. Impact of TAT-[NR2A15]2on potassium Kv1.3 channel and GABAAreceptor surface trafficking. (A) Amino acid sequences of NR2A, NR2B, and Kv1.3
C-termini (Rattus norvegicus). Note the high similarity among the three sequences in the last 5 aa, in the known PDZ binding site. The tracking of endogenous Kv1.3 potassium channel was done by detecting an extracellular epitope of the channel using an antibody-QD complex (schematic representation, Lower). For the experiments below, we analyzed 116 trajectories in presence TAT-[NS15]2and 209 trajectories in presence TAT-[NR2A15]2.(B) Plot of the MSD (inμm2) versus
time lag (in s) of surface Kv1.3 channel in presence of TAT-[NS15]2or TAT-[NR2A15]2.No significant effect (P > 0.05). (C) The percent of mobile surface Kv1.3
channel remain statistically similar in presence of TAT-[NS15]2or TAT-[NR2A15]2(P> 0.05). (D) The distributions (median, 25–75% range) and medians of Kv
diffusion coefficient were not significantly changed in presence of TAT-[NS15]2or TAT-[NR2A15]2(P> 0.05). (E) The surface distribution of Kv1.3 channels was
examined using single particle live distribution. The Kv channels are mostly extrasynaptic (approximately 80%). Incubations with TAT-[NS15]2(n = 6 dendritic
fields) or TAT-[NR2A15]2(n = 10 dendriticfields) did not affect the Kv1.3 channel surface distributions. (F) GABAAreceptor surface diffusion was assessed using
QDs coupled to an antibody directed against theα2 subunit of the receptor. Representative trajectories of single QDs show no effect on the mobility of GABAA
receptors after 10 to 20 min of incubation with TAT-[NS15]2or TAT-[NR2A15]2. (Scale bar: 1μm.)
Fig. S7. TAT-[NR2A15]2specifically blocks the interaction between 95 and NR2A subunit in rat brain homogenates. (A) Representative gels of co-IP of
PSD-95 with NR2 subunits in rat forebrain fractions treated with ligands. (B) TAT-[NR2A15]2reduced the OD ratio of PSD-95:NR2A whereas induced no significant
effect on OD ratio of PSD-95:NR2B. Values are means± SEM (n = 5 for TAT-[NS15]2and n = 8 for control and TAT-[NR2A15]2). Of note, although the specific
effect of the divalent ligand could be monitored in brain lysates with solubilized protein complexes, we also observed for this approach a high sensitivity with respect to the nature of the detergents used in particular with the existence of afine balance between conditions allowing for co-IP of the protein complexes of interest and conditions compatible with ligand-induced competition.
B 15-20 min [NR2A15]2 A NMDAR mEPSCs 20 pA C [NR2A ] [NS ] D 0-5 min 5 pA 50 ms + AP-5 50 ms [NS15]2 [NR2A15]2 0 5 15 20 0 0 5 15 20 0 0 5 15 20 0 5 15 20 10 20 30 ** 1 (ms) 10 20 30 40 Ampl. (pA) 0 10 20 30 0-5 15-20 0-5 15-20 0-5 15-20 0-5 15-20
Time (min) Time (min)
E F 20 pA 0-5 min [NR2A15]2 20 30 40 tude (pA) 10 15 20 e ncy ( H z ) 20 pA 100 ms 15-20 min [NR2A15]2 0-5 15-20 0 10 Time (min) Ampli 0-5 15-20 0 5 Time (min) Frequ e
Fig. S8. The [NR2A15]2functionally impacts only on miniature synaptic NMDAR currents. (A) Representative recordings of NMDAR mEPSCs at +30mV in the
presence of Ca2+/Mg2+(2/2 mM), NBQX (10μM; AMPAR antagonist), and bicuculline (20 μM; GABA
Aantagonist) in the whole cell configuration. The addition of
AP-5 (25μM) into the bath solution rapidly abolished NMDAR mEPSCs (Lower). The NMDAR mEPSC frequency range between 0.2 and 0.9 Hz, indicating that, on average, one event was detected every 2 s and overlap between events (example, Upper) was rare. (B) Averaged traces of NMDAR mEPSCs for two different time intervals: 0 to 5 min (black) and 15 to 20 min (gray) in the presence of [NR2A15]2(5μM) in the pipette solution. Note the slower decay after 15 to 20 min
infusion compared with 0 to 5 min. (C) Time constants,τ1, of the two time intervals (0–5 and 15–20 min). The decay was fitted using an exponential fit with
a fastτ1and a slowerτ2components. In the presence of [NR2A15]2(n = 6 neurons)τ1was significantly increased (**P < 0.01, paired t test) whereas no effect was
observed in the presence of [NS15]2(n = 5 neurons; P> 0.05, paired t test). The same results were obtained with the slower τ2component. (D) Amplitude of
NMDAR mEPSCs for the two time intervals (0–5 and 15–20 min) in the presence of [NR2A15]2or [NS15]2(P> 0.05, paired t test). (E) Representative traces of
AMPAR mEPSCs recorded at−50 mV and isolated with 1 μM TTX, 20 μM bicuculline, and 25 μM AP5 during two time intervals: 0 to 5 min (Upper) and 15 to 20 min (Lower) after whole cell configuration. (F) Neither the amplitude nor the frequency of AMPAR mEPSCs was affected by [NR2A15]2(n = 6 neurons).
Subunit NR2A 15 NR2B 15 NR2 X 15
Monovalent Divalent Monovalent Monovalent
A
Monovalent Divalent Monovalent Monovalent
NR2A + +++ - + NR2 B - - + + NR2A- S1462A - GABA A - Kv1. 3 -B
( - ) No detectable change in diff. ( + ) Higher diffusion ( +++ ) Very high diffusion
Fig. S9. Working model of TAT-[NR2A15]2. (A) Summary table of the impact of NR2-derived ligands on the synaptic diffusion of various surface subunits. (B) In
basal conditions, NMDARs interact with MAGUK proteins like PSD-95 through their C-terminal PDZ binding domain. This interaction strongly retains 2A-NMDARs within the synapse. The divalent ligand (TAT-)[NR2A15]2containing two binding motifs competes with 2A-NMDARs for the binding to specific
in-teractors. This highly specific competition induces a destabilization of 2A-NMDARs.
Fig. S10. Transduction of the BODIPY-TAT-[NR2A15]2ligand in hippocampal cultured neurons. (A) Neurons were incubated for 5 min with 50μM pyrene
butyrate and for 10 min with 5μM Bodipy-TAT-[NR2A15]2. Note the high transduction efficiency of the ligand within almost all processes. (Scale bar: 10 μm.) (B)
Colocalization of TAT-[NR2A15]2ligand with the postsynaptic marker shank. Neurons were incubated with 50μM pyrene butyrate followed by 10 min with 5
μM TAT-[NR2A15]2. After a 20-min wash, cells werefixed in PFA 4%/sucrose 4% in PBS solution for 15 min. They were then permeabilized using Triton 0.1% in
PBS solution. Neurons were incubated for 30 min with a rabbit anti-shank 3 antibody and for 30 min with a secondary anti-rabbit antibody coupled to an Alexa 568. (C) Consistent with the use of a saturating ligand concentration, no significant synaptic enrichment of both ligands was observed (TAT-[NS15]2, n = 939
synapses; TAT-[NR2A15]2, n = 426 synapses).
QD-NR2 Ab Syn. Pyr. But. TAT-peptide < 0.2 μg/ml 10-15 min Mitotracker 20 nM Pyrene Butyrate 50 μM 1-5 μM QD-NR2A Ab QD-Ab alone 0 4.10-3 8.10-3 Diffusion coefficient (µ m 2/s) QD-Ab + Py.Bu. QD-Ab + Py.Bu. + TAT-[NS15]2 500 nm
Fig. S11. Effect of pyrene butyrate and TAT peptide on basal NR-NMDAR surface diffusion. The nonspecific effects of pyrene butyrate or TAT-[NS15]2was
tested on the mobility of 2A-NMDARs. Whereas pyrene butyrate had no effect per se, TAT-[NS15]2increased the proportion of immobile receptors. Such effect
of TAT-[NS15]2was observed on 2B-NMDARs, GluR2-AMPARs, GABAARs, and reproduce using the monomeric TAT-[NS15]. This indicates that TAT-NS slightly
reduced the surface trafficking of neurotransmitter receptors, irrespective of the nature of the receptors or the structure of the TAT ligand.
![Fig. 2. TAT-[NR2A 15 ] 2 incubation does not affect native 2B-NMDARs or NR2A subunit mutant that does not bind to PDZ proteins](https://thumb-eu.123doks.com/thumbv2/123doknet/14753897.581508/4.877.56.419.89.874/fig-incubation-affect-native-nmdars-subunit-mutant-proteins.webp)
![Fig. 3. TAT-[NR2A 15 ] 2 specifically blocks the interaction between PSD-95 and NR2A subunit](https://thumb-eu.123doks.com/thumbv2/123doknet/14753897.581508/5.877.160.336.78.262/fig-tat-nr-specifically-blocks-interaction-psd-subunit.webp)
![Fig. S2. TAT-[NR2A 15 ] 2 increases the mobility of synaptic 2A-NMDARs and decreases the mobility of 2B-NMDARs in mature ( > 15 d in vitro) synapses](https://thumb-eu.123doks.com/thumbv2/123doknet/14753897.581508/12.877.218.660.83.540/increases-mobility-synaptic-nmdars-decreases-mobility-nmdars-synapses.webp)
![Fig. S6. Impact of TAT-[NR2A 15 ] 2 on potassium Kv1.3 channel and GABA A receptor surface traf fi cking](https://thumb-eu.123doks.com/thumbv2/123doknet/14753897.581508/15.877.216.659.137.647/fig-impact-potassium-channel-gaba-receptor-surface-cking.webp)
![Fig. S7. TAT-[NR2A 15 ] 2 speci fi cally blocks the interaction between PSD-95 and NR2A subunit in rat brain homogenates](https://thumb-eu.123doks.com/thumbv2/123doknet/14753897.581508/16.877.262.620.74.463/fig-speci-cally-blocks-interaction-subunit-brain-homogenates.webp)
