Potential candidate genes for improving Rice disease resistance
Texte intégral
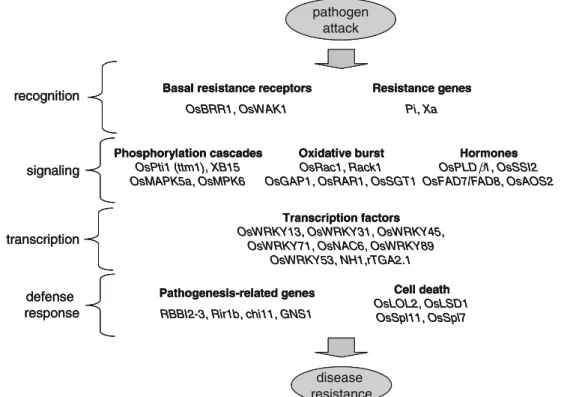
Figure
Documents relatifs
3K RG: 3000 Rice Genomes; ASTI-DOST: Advanced Science and Technology Institute, Department of Science and Technology of the Philippine Government; AWS: Amazon Web Services;
1 International Rice Research Institute, Los Baños, Philippines; 2 Centre for Crop Systems Analysis, Wageningen University, the Netherlands; 5 National Institute
After transformation we obtained 4 independent homozygous lines and all the lines show resistance against different isolates of rice blast (Magnaporthe oryzae), on
In parallel with genetics approaches, efforts have been made to screen mutant libraries for lines presenting alterations in root development, allowing for the identification of
While no product was effective in reducing Pythium damping-off in pepper, Prestop, Double Nickel LC, Rootshield Home and Garden, Rootshield PLUS WP, Mycostop, Previcur Flex, Serenade
Ten of these insertion lines were available through the OryzaTagLine (Larmande et al., 2008; Sallaud et al., 2004) collection, and we selected CEBiP, rTGA2.1, NH1, Pi21, SPL7
Blast incidence was reduced in DSMB cropping system and, on the contrary to the traditional cropping system, the mineral fertilisation had no effect on blast in DSMB.. N
