Synthesis, purification and structural and functional characterization of recombinant form of a common genetic variant of human luteinizing hormone
16
0
0
Texte intégral
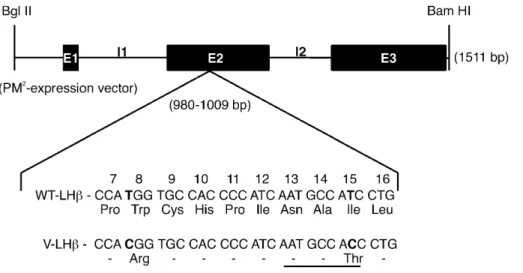
(2) 302. Human Molecular Genetics, 2002, Vol. 11, No. 3. Figure 1. Schematic presentation of the human LHβ gene, as well as the mutational changes in exon 2 of the V-LHβ gene with corresponding amino acid substitutions. To produce rec LH, the full-length (1511 bp) LHβ gene, a BglII–BamHI fragment was inserted into the eukaryotic expression vector PM2 (31,63–65). Three exons (E1, E2, E3) and two introns (I1, I2) and the glycosylation consensus sequence at position 13 (underlined) in V-LHβ, are illustrated.. The processing of N-linked oligosaccharides of glycoprotein hormones is tissue and dimer specific. Whereas LH and TSH, synthesized in gonadotropes and thyrotropes, respectively, possess sulfated (SO4) oligosaccharides and N-acetylgalactosamine (GalNAc) at their carbohydrate termini, pituitary FSH and placental hCG bear oligosaccharides terminating with sialic acid and galactose (Gal) (2,10). These differences determine the longer circulatory half-lives of FSH and hCG, whereas LH and TSH are cleared faster from the circulation by a hepatic receptor specific for sulfated glycoproteins (1,2,11,12). For this reason, the physiological relevance of recombinant gonadotropin preparations depends to a great extent on the nature of their glycosylation (see below). LH action is important for stimulating steroid biosynthesis of the ovary and triggering ovulation, whereas it maintains androgen production of testicular Leydig cells (13). Strong direct evidence for these functions has been provided by gonadal disorders observed in patients with inactivating mutations of the LHβ and LHR genes (14,15), as well as by the recently developed knockout models for the LHR gene (16,17). To date, only one inactivating mutation of the LHβ gene has been detected in a male with lack of postnatal sexual development (18), whereas no germ line mutations are known in the common α-subunit gene. In addition, there is a common polymorphic allele of the LHβ gene, caused by two point mutations and resulting in amino acid changes Trp8Arg and Ile15Thr, as well as in formation of an extra N-linked glycosylation consensus site (Asn-X-Ser/Thr) by the latter substitution (Fig. 1) (19–21). The carrier frequency of this common polymorphism varies from 0 to 53% in various populations (22–24). Evidence is mounting that it may predispose its carriers to mild alterations in LH-dependent reproductive functions, including changes of gonadal steroidogenesis, delayed pubertal progression, subfertility, lower incidence of polycystic ovarian syndrome (PCOS), and delayed appearance of breast cancer (15,25–30; I.Huhtaniemi, P.Salvén and H.Nevanlinna, unpublished data). This variant form of LH (V-LH) is structurally closer to hCG. which carries the same two altered amino acids in its β-chain (19–21,31). To gain better insight into the structure–function relationships of WT- and V-LH, reflecting the molecular basis of the differences observed between carriers of the WT and V-LHβ alleles in clinical observations, recombinant (rec) forms of these hormones were synthesized in human embryonic kidney (HEK 293) cells, purified and studied for their functional and structural properties. Earlier studies have shown that the C-termini of rec LH produced in Chinese hamster ovary (CHO) cells are sialylated in contrast to being sulfated, as is characteristic of pituitary LH. This is due to lack of N-acetylgalactosamine transferase and sufotransferase in CHO cells (2,32,33). It has also been demonstrated that rec bovine and human LHs synthesized in CHO cells have lower in vitro bioactivity than the respective pituitary hormones (2,32,34–36). It is therefore important for the biological relevance of the information that the carbohydrate structures of recombinant LH resemble as closely as possible those present in the native pituitary hormone. For this reason, HEK 293 cells were selected in this study for the rec LH production, since these cells have previously been shown to sulfate the carbohydrate moieties of glycoproteins in closely similar fashion to that observed in pituitary cells (37–39). RESULTS Purification of the rec WT- and V-LH The synthesized rec LHs were purified employing redsepharose CL-6B affinity, immunoaffinity and fast protein liquid chromatography (FPLC) Mono Q anion exchange chromatography (Fig. 2). Progesterone (P) production was determined as index of bioactivity during each step of purification. Increasing concentrations of rec WT- and V-LH (0–1000 µg/l) showed dose-dependent stimulation of P synthesis in mLTC-1 cells (Fig. 2A). The potency of V-LH was ∼4-fold higher over WT-LH, whereas the ED50s of these LH preparations were.
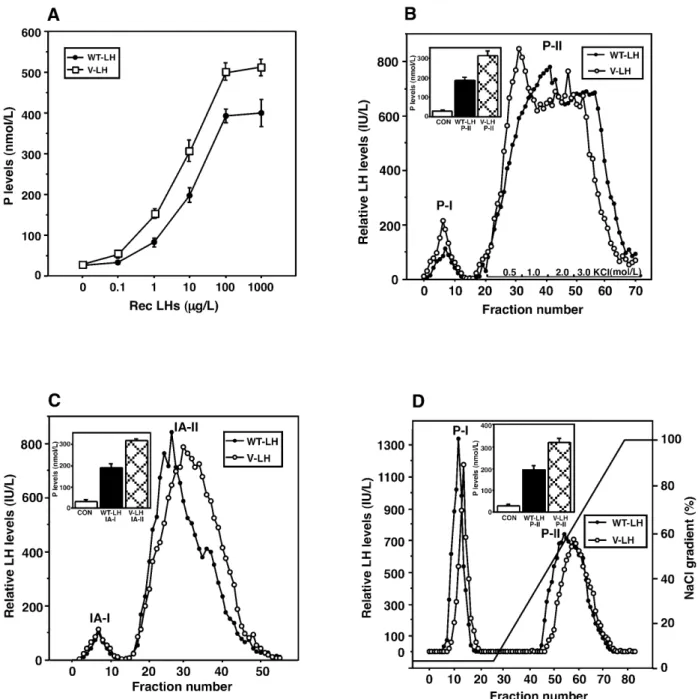
(3) Human Molecular Genetics, 2002, Vol. 11, No. 3. 303. Figure 2. Purification of the rec WT- and V-LH, and assessment of their biological activities, based on stimulation of P production at various steps. MLTC-1 cells were stimulated for 6 h with increasing concentrations of the rec LHs (0–1000 µg/l), and P levels in the media were determined by RIA (A). (B) Elution profile of WT- (closed circles) and V-LH (open circles) on red-sepharose CL-6B column chromatography. Biological activity of pooled peak-II (P-II) fractions in terms of P production was tested using 10 µg/l of rec LHs (inset). (C) Elution profile of WT- (closed circles) and V-LH (open circles) on immunoaffinity chromatography. Separation of unbound from bound materials was carried out by washing the column with buffer A (10 mmol/l sodium phosphate + 0.6% saline, pH 7.2). Bound materials were eluted with buffer B (buffer A + 3.0 mol/l NaI), as described in Materials and Methods. Pooled IA-II fractions were assessed for their bioactivity by mLTC-1 cell P synthesis (inset). (D) Elution profile of WT- (closed circles) and V-LH (open circles) on FPLC Mono Q anion exchange chromatography. After loading the materials, the column was washed with buffer A (20 mmol/l Tris–HCl pH 6.5) to remove unabsorbed materials, whereas adsorbed materials were eluted with buffer B (buffer A + 1.0 mol/l NaCl), as described in Materials and Methods. Inset, bioactivity of Mono Q P-II fractions in P production using 10 µg/l of the rec LHs. P levels measured are the means ± SEM of 3–5 experiments.. found to be 8–11 µg/l. Figure 2B shows the elution pattern and assessment of bioactivity of the WT- and V-LH on red-sepharose CL-6B column. Maximum elution of the LHs occurred at 0.5 mol/l of KCl; however, 3.0 mol/l KCl was used for their complete recovery. The pooled immunoreactive LH fractions (P-II) showed significant stimulation of P production, when. compared at a sub-maximal concentration (10 µg/l; corresponding to ∼60 IU/l) of the two rec LHs (Fig. 2B, inset). The pooled and dialyzed materials obtained from red-sepharose affinity chromatography were re-chromatographed on immunoaffinity column to achieve further purification. As illustrated in Figure 2C, a small unbound peak (IA-I) was eluted upon washing.
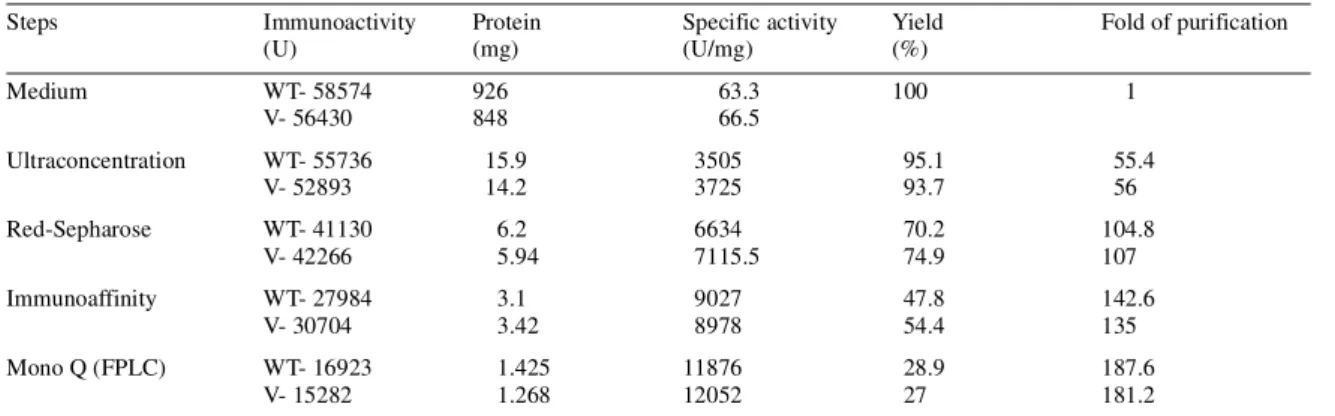
(4) 304. Human Molecular Genetics, 2002, Vol. 11, No. 3. Figure 3. Assessment of purity of the rec WT- and V-LH preparations by SDS–PAGE. Rec LH preparations (5 ng) from different steps of purification (lanes 2–9) as indicated, were electrophoresed on SDS buffer strips as described in Materials and Methods. The protein bands were visualized with silver staining. Molecular size markers are shown on both sides (lanes 1 and 10).. Table 1. Summary of purification of the rec WT- and V-LH Steps. Immunoactivity (U). Protein (mg). Medium. WT- 58574 V- 56430. 926 848. Ultraconcentration. WT- 55736 V- 52893. Red-Sepharose. WT- 41130 V- 42266. Immunoaffinity Mono Q (FPLC). 15.9 14.2. Specific activity (U/mg) 63.3 66.5. Yield (%) 100. Fold of purification 1. 3505 3725. 95.1 93.7. 55.4 56. 6.2 5.94. 6634 7115.5. 70.2 74.9. 104.8 107. WT- 27984 V- 30704. 3.1 3.42. 9027 8978. 47.8 54.4. 142.6 135. WT- 16923 V- 15282. 1.425 1.268. 11876 12052. 28.9 27. 187.6 181.2. The rec LHs were purified employing following steps, in consideration with their synthesis into the media. Fold of purification was calculated using specific activity of the rec LHs (WT, WT-LH; V, V-LH), considering pooled media as 1.. the column with buffer A (Materials and Methods). The bulk of the rec LHs was eluted with buffer B, as demonstrated by a clear immunoreactive peak (IA-II). Bioassay of the pooled IA-II fractions showed 7- and 9.5-fold increases in P production with WT- and V-LH, respectively, over non-stimulated cells (Fig. 2C, inset). The final purification upon FPLC Mono Q anion exchange chromatography (Fig. 2D) demonstrated an unabsorbed peak (P-I) during washing of the column with starting buffer A, while an absorbed peak (P-II) was eluted with buffer B at ∼60% of the NaCl gradient. Assessment of bioactivity of fractions P-II (10 µg/l) in terms of P production documented, as with the less pure fractions, higher bioactivity for V-LH than WT-LH (Fig. 2D, inset). The purities of the rec LH preparations were analyzed by SDS–PAGE (using 5 ng) during the progress of purification. The data presented in Figure 3 show that rec LHs obtained upon ultrafiltration exhibited several molecular weight protein bands (lanes 2 and 3). Stronger bands appearing at higher molecular weight were most likely due to aggregation of proteins in both cases. The P-II fraction on red-sepharose CL-6B column demonstrated significant removal of the non-specific protein. bands (lanes 4 and 5). Mono Q P-II fraction showed a clear single band of 31 kDa in size in both cases. Table 1 summarizes the purification procedure. The yield of immunoreactive LH protein gradually decreased as the specific activity and fold of purification increased. The recoveries of these LH preparations upon red-sepharose CL-6B column were 70 and 75% for WT- and V-LH, respectively, while the degree of purification was found to be 100-fold in both cases. Using immunoaffinity chromatography, the specific immunoreactivity of the LHs increased to ∼9000 IU/mg, with a recovery of ∼50% in both preparations. Finally, FPLC Mono Q chromatography resulted in ∼180-fold purification of rec WT- and V-LH, with yields of ∼1.4 and 1.2 mg, respectively. The specific activities of the purified rec WT- and V-LH preparations were 11876 and 12052 IU/mg, respectively. Effects of the rec LHs on steroidogenic acute regulatory protein (StAR) mRNA expression and cAMP production The biopotency of V-LH over WT-LH was ∼4-fold higher in terms of P production, which was consistent during the process of purification (Fig. 2). To further ascertain the stimulatory.
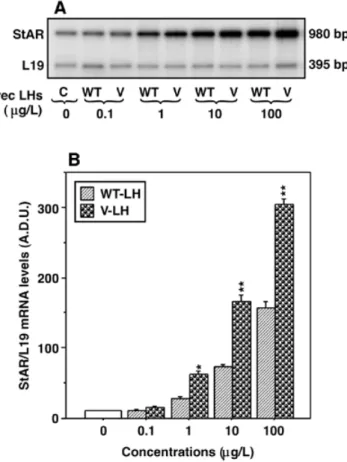
(5) Human Molecular Genetics, 2002, Vol. 11, No. 3. 305. Figure 5. Effects of rec LHs and hCG on cAMP production in mLTC-1 cells. Cells were stimulated for 2 h with increasing concentrations (0–1000 µg/l) of the hormones, in the presence of 0.25 mmol/l of IBMX, and cAMP levels in the media were determined. The hormones used were rec hCG, V-LH, WT-LH and WT-LH (CHO), i.e. recombinant LH produced in CHO cells (Organon). The results are the means ± SEM of four independent experiments.. Figure 4. Effects of rec WT- and V-LH on StAR mRNA expression in mLTC-1 cells. Cells were stimulated for 6 h with increasing doses of rec LHs (0–100 µg/l), total RNA was extracted from the different stimulation groups and subjected to RT–PCR analysis of StAR mRNA expression as described in Materials and Methods. The variation in RT–PCR efficiency was evaluated by co-amplifying ribosomal L19 protein gene mRNA with each sample. (A) A representative autoradiogram showing StAR mRNA expression in control cells (C) and in the presence of increasing concentrations (0–100 µg/l) of rec WT-LH (WT) and V-LH (V). The arbitrary densitometric units (A.D.U.) of StAR mRNA expression in each band were quantified and normalized for intensity of the corresponding L19 bands (B). The values are the means ± SEM of three independent experiments. The asterisks represent significant differences over WT-LH at the specific concentrations indicated. *, P < 0.05; **, P < 0.01.. activity of V-LH on steroidogenesis, expression of the StAR gene, shown to be acutely involved in regulating steroid biosynthesis, was investigated. Figure 4 illustrates dose-dependent induction of StAR mRNA expression with increasing concentrations of the rec LHs (0–100 µg/l), and ∼2-fold higher biopotency of V-LH compared with WT-LH. These data corroborate the higher potency of V-LH in stimulation of P production (Fig. 2), and the interdependence of StAR expression and steroidogenesis. The in vitro bioactivity of rec WT- and V-LH was further assessed by determining cAMP production, in comparison to hCG (CR-127) and human rec LH produced in CHO cells [WT-LH (CHO), Organon], utilizing mLTC-1 cells (Fig. 5). Dose-dependent cAMP responses were observed with increasing hormone concentrations (0–1000 µg/l), and the maximum response to V-LH was found to be ∼25% higher than that evoked by WT-LH (Fig. 5). The ED50s of the different hormone preparations did not show significant differences, ranging between 12–17 µg/l.. Figure 6. Circulating half-lives of the different LH/hCG preparations (Fig. 5 legend) in vivo. Chronically catheterized adult male rats were treated with cetrorelix, followed by intravenous injections of the hormones (750 IU/rat), as described in Materials and Methods. Venous blood samples were obtained at indicated times (0–300 min), and serum hormone levels were determined. The data represented are the means ± SEM of six to nine rats per group.. In vivo half-lives of the rec LH/hCG preparations, and their effects in ovulation induction To characterize the eventual differences in in vivo behavior of the different LH/hCG preparations, we evaluated their circulating clearance rates after injection into rat circulation. Figure 6 demonstrates that the half-life of V-LH was much faster (T1/2; 5–9 min) in comparison to WT-LH (12–22 min), WT-LH (CHO) (25–34 min) or hCG (50–70 min). The difference between the rec WT-LH forms prepared on HEK 293 and CHO.
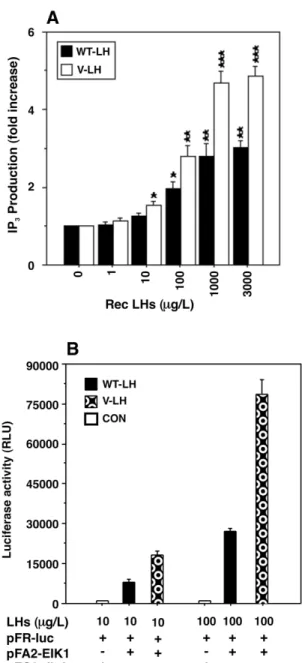
(6) 306. Human Molecular Genetics, 2002, Vol. 11, No. 3. Figure 7. In vivo biopotency of rec WT- and V-LH, as monitored by ovulation induction. Adult female Sprague–Dawley rats primed with PMSG were injected with increasing doses of rec WT- and V-LH (0–100 IU/rat), as described in Materials and Methods. The number of ovulated oocytes was determined in the oviducts ( ± SD). The numbers of rats ovulated out of the total are shown in parentheses on the top of each bar.. cells apparently reflects the differences in glycosylation between the two cell types, demonstrating sulfated and sialylated LHs, respectively. Since V-LH appeared to have higher in vitro bioactivity than WT-LH, we next assessed whether the same difference prevailed in vivo in induction of ovulation of adult female rats primed with PMSG. The data presented in Figure 7 demonstrate that rats injected with increasing amounts of WT- and V-LH (0–100 IU/l) did not show significant differences in ovulation induction. Ovulation was absent or observed in a part of the animals with the lower dose of the rec LHs (up to 25 IU/l), while at the highest dose of 100 IU/l, both LH forms were capable of ovulating all animals with quantitatively similar yield of oocytes. Receptor binding of the rec LHs To corroborate the in vitro findings, we next analyzed ligandbinding activities of the rec LHs utilizing intact mLTC-1 cells. Displacement of specific [125I]iodo-hCG binding increased gradually with increasing concentration of unlabeled hormones (rec LHs and hCG), and 50% displacement was observed with all hormones at ∼45 µg/l (Fig. 8A). In fact, no significant differences were observed with WT- and V-LH in displacing [125I]iodo-hCG binding. The equilibrium dissociation constants (Kds) of these hormones, as calculated from Scatchard plots (Fig. 8B), were between 0.39 and 0.42 nmol/l, also indicating that the mutational changes in V-LH did not affect its affinity to the LHR. Influence of the rec LHs on inositol phosphate (IP3) production and on ElK1 trans-reporting MAPK activity The biopotency of V-LH in signal transduction was evaluated further in terms of IP3 production. HEK 293 cells transfected. Figure 8. Displacement of [125I]iodo-hCG binding with increasing concentrations of the rec LHs and hCG. (A) mLTC-1 cells (2.5 × 105 cells/incubation) were incubated with [125I]iodo-hCG ( ∼150 000 c.p.m.) in the presence of varying amounts (0–1000 µg/l) of hormones, as demonstrated in Materials and Methods. Specific binding was calculated by subtracting non-specific from total binding. (B) The affinity of WT-LH, V-LH, WT-LH (CHO) and hCG binding to the mLTC-1 cell LH/hCG receptor, using [125I]iodo-hCG as tracer. The affinities (Kds) were determined by Scatchard analysis as described in Materials and Methods. The Kd values were found to be 0.39, 0.41, 0.40 and 0.42 nmol/l for WT-LH, V-LH, WT-LH (CHO) and hCG, respectively. These experiments were repeated three times, and data from a representative experiment are presented.. with human LHR demonstrated a dose-dependent increase in IP3 production in response to increasing concentrations (0–3000 µg/l) of the rec LHs (Fig. 9A). The maximum response was 1.8 ± 0.15-fold higher with V-LH in comparison to WT-LH. Furthermore, the effects of WT- and V-LH were studied on the MAPK signaling pathway. HEK 293 cells were transfected with the ElK1 trans-activator plasmids followed by stimulation with two concentrations of the rec LHs (10 and 100 µg/l). The results presented in Figure 9B demonstrate that V-LH augmented 2–3-fold higher stimulation of MAPK-mediated luciferase activity than WT-LH at the hormone concentrations used..
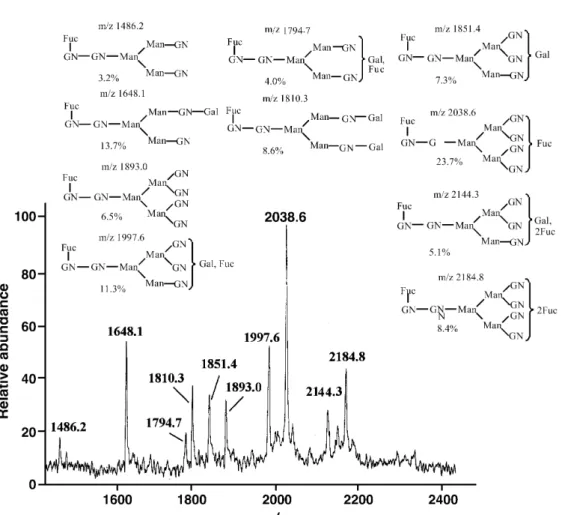
(7) Human Molecular Genetics, 2002, Vol. 11, No. 3. 307. and the glycans were analyzed using both positive and negative ion matrix-assisted laser desorption ionization–mass spectrometry (MALDI–MS). The positive ion MS profile of WT-LH showed 10 prominent ions, and the corresponding N-linked structures are presented in Figure 10A. As illustrated in Figure 10B, MS analysis of the same sample by negative ion extraction showed a different set of eight glycoforms. Positive ion MALDI–MS analysis of the V-LH sample suggested the presence of 13 glycans, together with their N-linked structures (Fig. 11A). Interestingly, mass analysis with negative ion extraction of V-LH glycans yielded a significantly different finding with a total of 23 N-linked glycans (Fig. 11B). Separate positive and negative ion MALDI-MS analysis usually provides the greatest sensitivity for independent assessment of neutral and acidic glycans, but the relative abundance between the glycoforms cannot be accurately assessed. A relative comparison of all glycotypes (acid and neutral) can be achieved by methylation which blocks the ionic character of individual species, but this chemical step requires larger amounts of sample, not available for this assay. These results document that mutations in the LHβ gene clearly affect high mannose processing in the V-LH, which might be associated with the functional consequences documented both in vivo and in vitro. DISCUSSION. Figure 9. Effects of rec WT- and V-LH on IP3 production, and activity of the MAPK pathways. (A) HEK 293 cells were transiently transfected with pcDNAneo-hLHR using the lipofectamine method. 24 h after transfection, cells were labeled with [3H]inositol as described in Materials and Methods. Cells were stimulated (60 min) with increasing concentrations of the rec LHs (0–3000 µg/l) in the presence of 12.5 mmol/l LiCl, and IP3 levels in the cell extract were determined. The results are the means ± SEM of three independent experiments. The asterisks represent significant differences in comparison to control. *, P < 0.05; **, P < 0.01, ***, P < 0.001. (B) HEK 293 cells were transfected with ElK1 trans-reporting plasmids. 36 h later, cells were stimulated for 8 h with two different concentrations of the rec LHs (10 and 100 µg/l). Luciferase activity in the cell lysate was determined. The results represent the means ± SEM of four independent experiments. pFR-luc, reporter plasmid; pFA2-ElK1, pathway-specific fusion trans-activator plasmid; pFC2-dbd, GAL4 DNA binding domain (dbd) that lacks the specific activation domain (EIK1).. Mass spectrometric (MS) analysis of the carbohydrate structures of WT- and V-LH To elucidate further the structural differences of V-LH and WT-LH, we carried out MS analysis of their N-linked glycans. These residues were enzymatically released from the protein. Gonadal development, maturation and reproductive functions are intricately regulated by the two gonadotropins, LH and FSH (1,40,41). Structural alterations, caused by mutations in genes of gonadotropins or their cognate receptors, have been found in abnormalities of reproductive function (14,15). Also milder genetic alterations, due to polymorphisms of these genes, have been demonstrated to have phenotypic effects in both sexes (24–31,42). The present investigation was focused on exploration of the structure–function relationships of WT- and V-LH using recombinant forms of these hormones. The biochemical and functional differences of these two rec LH forms provide a clear molecular basis for the clinical observations on their differential phenotypic effects. Besides the amino acid alterations caused by the two point mutations, the clear differences in their glycosylation, demonstrating incomplete processing of the N-linked high mannose glycans (Man5,8,9) in rec V-LH, apparently contribute to the functional differences observed between WT- and V-LH in vivo and in vitro. To elucidate the physiological relevance of the LHβ mutations, rec WT- and V-LH were produced in HEK 293 cells, by cotransfecting the common α-subunit gene with either WT- or V-LHβ gene. This procedure has been shown not to affect the assembly of α- and β-subunits or secretion of the dimer of hCG and LH in CHO cells (43). Purification of the rec LHs was carried out by red-sepharose CL-6B affinity, immunoaffinity and FPLC Mono Q anion exchange chromatography. Among the procedures employed, Mono Q anion exchange chromatography was critical for achieving pure rec LHs. SDS–PAGE analysis of the purified materials revealed a sharp band of ∼31 kDa with both LH preparations, in accordance with the known molecular size of LH of 32–33 kDa (1,33). The apparent slight difference in molecular weight may be due to the pattern of glycosylation, N-acetyl-neuraminic acids and sulfate contents (2,33,36,44)..
(8) 308. Human Molecular Genetics, 2002, Vol. 11, No. 3. Figure 10. Positive ion matrix-assisted laser desorption ionization (+MALDI) mass spectrum (MS) following profile of (M+Na)+ with PNGase-F release. The N-linked glycan structures of WT-LH are presented based on ions detected (A). The matrix used was DHB. Negative ion MALDI mass spectrum (–MALDI-MS) of WT-LH, with profile of (M-H)– following PNGase-F release (B, opposite). The N-linked glycan structures are proposed based on ions detected. The matrix used was THAP.. Although the in vitro bioactivity of V-LH in terms of mLTC-1 cell P production was higher, the distribution pattern of isoforms of the rec LHs was rather similar at pI range 6.6–7.8 (data not shown), in agreement with previous findings (33,45). The degree of purity of the rec LHs obviated potential interference of contaminating molecules, and provided the opportunity to characterize eventual effects of the two point mutations on carbohydrate composition. The MS glycan profiles observed with the rec LHs were in general agreement with those of other pituitary glycoprotein hormones, including differences in galactose, GalNAc, sialic acid and sulfate contents (2,33–36,44,45). However, very clear differences were observed in the profiles of WT-LH and V-LH, especially in the abundance of high mannose glycans (Man5,8,9), which might be related to the functional differences observed. The mutations may have altered Golgi trafficking where the resultant structure is the consequence of a different set of glycosyltransferases. It was surprising to note that the two amino acid changes in the LH chain had such dramatic effects on quality of glycosylation. The conformational changes at the N-terminus on V-LHβ, i.e. amino acid substitutions of the two hydrophobic residues at. codon 8 and 15 from Trp to Arg and Ile to Thr, respectively, could induce greater N-terminal hydrophilicity that might affect functional properties of the hormone. The Trp8Arg substitution in the V-LH molecule has been shown to be predominantly responsible for the altered functional characteristics of V-LH in vitro and in vivo (31). A recent study demonstrated the presence of several point mutations in the V-LHβ promoter sequence which was ∼50% more active than the WT-LHβ promoter along with several differences in response to hormonal stimulation (46). These characteristics of the WT- and V-LHβ promoters probably contributed to the difference observed in their physiological functions. The occurrence of V-LH in women has been shown to correlate with reproductive abnormalities, including menstrual disorders, PCOS, protection from breast cancer, and subfertility (Introduction). Our results show that V-LH binds to LHR with similar affinity to WT-LH, as demonstrated in monitoring Kd value (∼0.4 nmol/l) and in ovulation induction, suggesting that receptor–ligand interaction is not altered. However, V-LH demonstrated increased bioactivity in P and cAMP accumulation in mLTC-1 cells, while the ED50 values were found to be between 8 and 17 µg/l in both cases. The rec LH forms.
(9) Human Molecular Genetics, 2002, Vol. 11, No. 3. behaved in vitro and in vivo in identical fashion with hormones present in circulation of homozygotes for the WT- or V-LHβ alleles (22). The increased steroidogenic response to V-LH in terms of P accumulation was further confirmed by determining StAR gene expression, which has been shown to be intimately associated with steroid hormone biosynthesis (47). The increased bioactivity of V-LH in mLTC-1 cells, as monitored by their cAMP, P and StAR mRNA responses, was further confirmed in non-steroidogenic cells. In accordance, the HEK 293 cells expressing human LHR cDNA demonstrated higher response of IP3 production and MAPK activation to V-LH than to WT-LH. The Ile to Thr alteration introduces a new consensus site for N-glycosylation at Asn13 of the LHβ protein. Recent studies demonstrated that Asn13 could be glycosylated in V-LH, resulting in four terminally sulfated carbohydrate side chains in comparison to three in the WT-LH, and the altered function could be attributed to a number of factors (31,48). The hormonal potency may depend on glycan sulfation and/or sialylation of Asn-linked oligosaccharides affecting the net charge on the molecule, and these mutations may alter the determinants critical for GalNAc and SO4 addition. Subsequently,. 309. the importance of the Asn13 in hCGβ and bovine LHβ has been shown for hormonal binding and signal transduction (49,50). Previous reports from this (22,51) and other (31) laboratories provide evidence that the bio-immuno ratio of V-LH homozygotes is higher than that of WT-LH controls. Our current data also show that V-LH has a shorter half-life (5–9 min) in rat circulation, in comparison to WT-LH (12–22 min), WT-LH in CHO cells (25–34 min) or hCG (50–70 min), which are in general agreement with the previous findings on serum LH from WT- or V-LH homozygotes (22). The faster clearance rate of V-LH was unexpected since the mutational changes in V-LHβ bring it structurally closer to hCGβ, which has a longer circulating clearance rate in rats (52). It has been demonstrated that rec glycoproteins produced in CHO cells possess sialylated carbohydrates that affect the circulating half-life (33,37,39,53). Indeed, our studies clearly demonstrated that the in vivo half-life of rec WT-LH produced in CHO cells was clearly longer than that produced in HEK 293 cells, which is a consequence of glycan capping with neuraminyl or sulfate residues in the latter (37,39). These data are consistent with terminally sulfated oligosaccharides in pituitary LH, which are known to be involved in its rapid clearance from the circulation (12,32). Moreover, the higher valency of sulfated carbohydrate moieties.
(10) 310. Human Molecular Genetics, 2002, Vol. 11, No. 3. Figure 11. Positive ion MALDI mass spectrum (+MALDI-MS) of V-LH, following the profile of (M+Na)+ with PNGase-F release. The biantenary structures of N-linked glycans proposed are based on ions detected in MS (A). The matrix used was DHB. Negative ion MALDI mass spectrum (–MALDI-MS) of V-LH, and profile of (M-H)– following PNGase-F release. The N-linked glycan structures of V-LH are presented (B, opposite). The matrix used was THAP.. in V-LH (four versus three in WT-LH) is the likely explanation for its shorter half-life, since it would probably be more actively eliminated from circulation by the hepatic receptor for sulfated glycoproteins (12,54). Several studies have demonstrated that sulfation could occur on numerous glycan moieties other than GalNAc (55–58); however, its consequences in case of V-LH require further investigation. The mutations of glycoprotein hormones so far reported have concerned with inactivating alterations. A deletion in the second and third nucleotides of Val61 in the FSHβ gene is associated with primary amenorrhea (59), while a nonsense mutation in TSHβ caused familial hypothyroidism leading to infertility (60). A point mutation (Gln54Arg) in the LHβ gene has been shown in a male patient with hypogonadism (18). In contrast, the alterations in LHβ gene in the present study showed activation of the different signal transduction pathways in vitro. Suganuma and co-workers (19,25). observed that four out of five homozygotes associated with menstrual disturbances including these mutations exhibited hypersecretion of LH following gonadotropin-releasing hormone (GnRH) administration, a situation closely resembling PCOS (61). In the present findings, the high mannose structures observed in V-LH may suggest incomplete maturation of glycan chains, altering the molecule’s surface topology and subsequent functional roles. Since all signaling pathways of LH action were similarly affected by V-LH, i.e. no effect on ED50, but clear increase in maximal response, we can conclude that the divergence between magnitude of the different pathways is not affected by the mutational alterations of V-LH. This is in contrast to recent findings on a constitutively activating mutation of the LHR, where a specific alteration appears to activate the cAMP and IP3 pathways to different extents (62)..
(11) Human Molecular Genetics, 2002, Vol. 11, No. 3. Collectively, purification of the synthesized rec forms WTand V-LH has provided an opportunity to evaluate the structure–function relationships of the latter hormone. The increased in vitro bioactivity of V-LH with shorter in vivo half-life may affect gonadal function at various levels and be responsible for the multiple, albeit mild, reproductive disorders detected in carriers of the V-LHβ allele. MATERIALS AND METHODS Plasmids and transfections The mutations in exon 2 of the LHβ gene (including three exons and two introns) at codons 8 and 15, from Trp (TGG) to Arg (CGG) and Ile (ATC) to Thr (ACC), respectively, were constructed by site-directed mutagenesis (Fig. 1). Mutational changes were confirmed by sequencing with fluorescent dye termination reaction (Prism Ready Reaction Dye Termination Cycle Sequencing Kit) using automated sequencer (Perkin-Elmer, Foster City, CA). The full-length LHβ gene (1511 bp), a BglII–BamHI fragment, was. 311. inserted into eukaryotic expression vector PM2 (Fig. 1), downstream of the Harvey murine sarcoma virus long terminal repeat as described previously (31,63,64). The PM2 containing 2.4 kb α-minigene (a BamHI fragment) has two coding regions (exons 2+3 and 4), a single intron and polyadenylation signals 3′ to exon 4 (63,65). HEK 293 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM)-Ham’s F-12 (1:1; Life Technologies Inc., Gaithersburg, MD), supplemented with 10% fetal calf serum containing antibiotics (6). Transfection studies were carried out at 65–75% confluency of cells by lipofectamine (Life Technologies Inc.), using a 2:1 ratio of the α-subunit minigene (31,63–65) with either WT- or V-LHβ genes. Following 48 h of transfection, rec LH levels were determined in media by immunofluorometric assay (IFMA, Delfia LH Spec, Wallac, Turku, Finland). For generation of clonal lines, cells were trypsinized and split into selection medium containing 800 µg/l of geneticin (G418, Sigma Chemical Co., St Louis, MO). Following 3–4 weeks, resistant colonies were picked, expanded and screened for the level of LH production..
(12) 312. Human Molecular Genetics, 2002, Vol. 11, No. 3. For studying the effects of rec LHs on IP3 response, HEK 293 cells were transiently transfected (as above) with the full-length human LH receptor (hLHR) cDNA, a NheI–BamHI fragment (2.1 kb) (6), cloned into pcDNAneo 3.1 (Invitrogen, San Diego). The pathdetect EIK1 trans-reporting plasmids (Stratagene) were used for evaluating the MAPK pathway. Synthesis of the rec WT- and V-LH and their immunoassay The clonal lines of choice, WT-78 and V-24, were subsequently cultured in the presence of 300 mg/l of G418. The culture was initiated in DMEM/F-12 growth medium supplemented with 360 mg/l of L-glutamine (Merck, Darmstadt, Germany). The glucose and lactate levels were frequently determined in the media. Finally, serum-free medium was used containing 0.5 µg/l insulin, 5 mg/l transferrin and 1 µmol/l of 2,3-dehydro2-deoxy-N-acetyl-neuraminic acid. The production of rec LHs in the media was determined by a solid phase, two-site IFMA based on a direct sandwich technique (Delfia LH Spec). In the first step, hLH reacted with the monoclonal antibodies directed against a specific antigenic site of the hLHβ subunit, and in the second step, europiumlabeled antibodies specific to another antigenic epitope of the hLHβ subunit were reacted with hLH or its β-subunit bound to the solid phase antibody. The standard for the assay was calibrated with the WHO 2nd International Reference Preparation for pituitary LH immunoassay (80/552). The concentrations of rec LHs used in the subsequent experiments were calibrated using this assay. The LH Spec assay detects WT- and V-LH with similar stoichiometry (23,31). Purification of the rec LHs The media collected from the WT- and V-LH expressing cells were first concentrated using the Pellicon™-2 ultrafiltration (Millipore Corporation, Bedford, MA) system. The concentrated rec LHs were further purified using the following steps: (i) Red-Sepharose CL-6B chromatography. The first step of purification was carried out with red-sepharose CL-6B column (Amersham Pharmacia Biotech, Uppsala, Sweden). The column (1.8 × 32 cm) was equilibrated with sodium phosphate buffer (25 mmol/l, pH 6.8) in a cold room (4–6°C), at a flow rate of 20 ml/h. Before loading, the materials (WT- and V-LH) were passed through a sterile 0.2 µm filter (Gelman Sciences, Ann Arbor, MI), equilibrated by dialysis with eluting buffer containing NaCl (50 mmol/l). The materials were loaded onto the column and allowed to bind for 60–90 min, and washed thereafter with starting buffer, as described previously (33). Bound proteins were eluted with different concentrations of KCl (0.5, 1.0, 2.0 and 3.0 mol/l), and the fractions collected were measured for their LH immunoreactivity, pooled, dialyzed and processed for the next step of purification. (ii) Immunoaffinity chromatography. This step was performed using cyanogen bromide (CNBr)-activated Sepharose 4 Fast Flow matrix (Amersham Pharmacia). The coupling of monoclonal anti-hLH (5303; Medix Biochemica, Kauniainen, Finland) to matrix was carried out following the manufacturer’s instructions, with minor modifications. Briefly, the coupling was performed at 4°C for 12–16 h, and anti-LH. tagged sepharose was then loaded onto the HR 10/10 column (Amersham Pharmacia). The pooled fraction obtained on red-sepharose CL-6B chromatography was concentrated and equilibrated with buffer A (10 mmol/l sodium phosphate containing 0.6% saline, pH 7.2) using centrifugal concentrators (Centriprep-10; Amicon Inc., Beverly, MA), loaded onto the column, and allowed to bind for 4–6 h at 4°C. Bound LH was eluted with buffer B, i.e. buffer A containing 3.0 mol/l sodium iodide (NaI) as a chaotropic agent. The immunoreactive peak fractions were pooled, dialyzed against 10 mmol/l PBS pH 7.2, lyophilized and stored at –70°C. (iii)FPLC. The immunoaffinity pooled fraction was reconstituted in Tris–HCl buffer (20 mmol/l, pH 7.2), desalted and subjected to FPLC Mono Q anion exchange chromatography (Amersham Pharmacia). The column (HR 5/5) was equilibrated with buffer A (20 mmol/l Tris–HCl pH 6.5), at a flow rate of 0.5 ml/min. The adsorbed materials were eluted with buffer B, i.e. buffer A containing 1.0 mol/l NaCl. The immunoreactive peak fractions were pooled, lyophilized and stored at –70°C for further studies. SDS–PAGE The purities of WT- and V-LH during the process of purification were assessed by PhastGel® gradient system using SDS buffer strips (Amersham Pharmacia). The samples (5 ng) were prepared into sample buffer containing 10 mmol/l Tris–HCl, 1.0 mmol/l EDTA pH 8.0, treated with 5% SDS, 10% β-mercaptoethanol, and heated at 80–100°C for 5 min, followed by electrophoresis, run with low molecular weight protein markers. Following completion of the run, gels were silver stained using an automatic development program (Amersham Pharmacia). Assessment of in vivo bioactivity of the rec LHs in ovulation induction and circulatory clearance rate Mature female Sprague–Dawley rats (180–230 g), were housed under controlled lighting schedule (14 h light and 10 h darkness), fed commercial diet and water ad libitum. The rats primed with gonadotropin (PMSG, 10 IU/100 g body weight; Sigma) were injected in the presence of increasing doses of the rec LHs (0–100 IU/rat). A control group was maintained with physiological saline. Following 24 h, the rats were killed with CO2 asphyxia, and the numbers of ovulated oocytes were counted in oviducts (33). Circulating half-lives of WT- and V-LH were determined in adult Sprague–Dawley male rats in comparison to hCG and WT-LH produced in CHO cells [WT-LH (CHO); Organon, Oss, The Netherlands]. A technique of chronical intravenous catheterization was used, as described previously (66). The rats were first treated with the GnRH antagonist cetrorelix (ASTA, Frankfurt am Main, Germany; 100 µg/rat), followed by intravenous injection of the LH/hCG preparations (750 IU/rat). At specified times (0–300 min), blood samples were collected, serum separated, and hormone levels were measured (33,66). The animals were maintained in accordance with guidelines of the Turku and Geneva University Ethical Committees for the Use and Care of Experimental Animals, and appropriate permissions were obtained for the experiments..
(13) Human Molecular Genetics, 2002, Vol. 11, No. 3. Efficacy of the rec WT- and V-LH in receptor binding Radiolabeling of hCG (CR-127; biological potency 14 900 IU/mg, NIH, Bethesda, MD), and [125I]iodo-hCG binding studies were carried out using intact mLTC-1 cells, as described previously (6). The specific activities of the labeled hormone batches were found to be between 27 and 35 Ci/g. For displacement studies, 2.5 × 105 cells were incubated with [125I]iodo-hCG (∼150 000 c.p.m.) either in the absence (total) or presence (non-specific) of increasing amounts of unlabeled hCG (CR-127) or rec LHs (0–1000 µg/l). The reaction was terminated by placing the tubes on ice after overnight incubation, and followed by 3 ml ice-cold mixture of Dulbecco’s PBS (10 mmol/l, pH 7.2)-BSA (0.1%). Specific binding was determined by subtracting non-specific binding from total binding in a γ-spectrometer (1260 Multigamma II, Wallac). The affinities (Kds) of these hormones were determined by converting the binding inhibition data into Scatchard plots (6). Isolation of RNA, and RT–PCR analysis Total RNA was extracted from different stimulation groups following the single step acid guanidinium thiocyanatephenol-chloroform extraction method (67). The isolation and amplification of mouse (mLTC-1) StAR cDNA was carried out as previously described (68,69). The primer pairs used were: StAR sense 5′-GACCTTGAAAGGCTCAGGAAGAAC-3′ (bases 51–27) and antisense 5′-TAGCTGAAGATGGACAGACTTGC-3′ (bases 931–908). The variation in RT–PCR efficiency was determined by L19 ribosomal protein gene co-amplified in each sample using specific primer pairs (69). RT and PCR of the target genes were sequentially run in the same assay tube with 2 µg of total RNA, using a previously described procedure (68,69). Briefly, the reaction was started at 50°C for 15 min (RT), followed by denaturation at 97°C for 5 min. The PCR was then run for 17 cycles, defining denaturation at 96°C for 1.5 min, annealing at 55°C for 1.5 min and extension at 72°C for 5 min (PTC-200, Peltier Thermal Cycler; MJ Research, Waltham, MA). A final cycle of extension at 72°C for 15 min was included. The PCR products were examined in 1.2% agarose gel. The gels were then vacuum dried and exposed to Fuji X-ray film (Tokyo, Japan) at 4°C for 1–3 h. The relative mRNA levels of StAR and L19 were quantified by phosphorimaging and densitometry (Tina 2.0 Package, Straubenhardt, Germany). Effects of the rec LHs on IP3 production, and on pathdetect ElK1 trans-reporting assay HEK 293 cells were transiently transfected with human LHR expressing plasmid pcDNAneo-hLHR (see above). Briefly, 24 h after transfection, cells (2.5–3 × 105 cells/well in 6-well plate) were labeled for 24–30 h using 2 µCi/ml of [3H]-inositol (NEN Life Sciences Products Inc., Boston, MA). After washing, cells were stimulated for 45–60 min with increasing concentrations of WT- and V-LH (0–3 × 104 µg/l) in the presence of 12.5 mmol/l LiCl. The incubation was terminated by addition of ice-cold perchloric acid to a 2.5% final concentration. The supernatants were equilibrated with 1.5 M KOH and recollected by centrifugation. IP3 levels in the cell lysate were determined in AG-1 anion exchange column (Bio-Rad, Hercules, CA), as. 313. previously demonstrated (70). The radioactivity in different fractions was counted in a β-spectrometer. The effects WT- and V-LH were assessed on the MAPK pathway employing trans-reporting assay (Stratagene). In brief, HEK 293 cells were transfected with trans-activator plasmids using lipofectamine (Gibco BRL). Following 36 h, cells were stimulated for 8 h with two different concentrations of WT- and V-LH (10 and 100 µg/l), and luciferase activity in the cell lysates was determined (chemiluminescence), using 1251 luminometer (BioOrbit, Turku, Finland). Mass spectrometry The FPLC purified rec LHs were analyzed by MALDI-MS. Spectra were taken on a TofSpec-E time-of-flight mass spectrometer (Micromass, Manchester, England) equipped with delayed extraction. These MALDI-MS measurements were carried out using a nitrogen laser (337 nm, 4 ns pulse) operated in either the reflectron (20 kV accelerating voltage) or linear mode (25 kV accelerating voltage), as described previously (71,72). Both positive and negative ions were accelerated to 20 kV after 80 ns delay. The positive ion extraction was carried out with 2,5-dihydroxybenzoic acid (DHB) matrix, whereas for negative ion extraction the matrix used was 2´,4´,6´-trihydroxyacetophenone (THAP). Determination of progesterone and cAMP The culture and maintenance of mLTC-1 mouse Leydig tumor cells (73) were carried out in Waymouth’s growth medium, as described previously (68,69). Cells were stimulated either for 2 h (cAMP) or 6 h (P) in the presence of 0.25 mmol/l 3-isobutyl1-methyl xanthine (IBMX), with varying concentrations of hormones as specified elsewhere. cAMP and P levels in the media were assessed by specific RIAs (74,75). Data analysis The data presented are the means ± SEM, unless otherwise indicated. The statistically significant differences were analyzed by one-way analysis of variance (ANOVA), followed by Fisher’s protected least significant differences test using the Statview program (Abacus Concepts Inc., Berkeley, CA) fitted with the Macintosh computer. P < 0.05 was considered statistically significant. ACKNOWLEDGEMENTS The authors would like to thank Dr I.Boime (Washington University School of Medicine, St Louis, MO), for the generous gift of α-minigene construct. We would like to thank M.Vehniäinen (Department of Biochemistry, University of Turku), for providing us with the ultraconcentration system. Our special thanks to H.Peuravuori (Department of Pathology, University of Turku) for his active cooperation in relation to the FPLC system. The cordial cooperation of Dr P.Pakarinen is also gratefully acknowledged. Finally, we thank Ms Riikka Kytömaa and Ms Tarja Laiho for their superb technical assistance. This work was supported by grants from the Academy of Finland, The Sigrid Jusélius Foundation and the Foundation for the Finnish Cancer Societies to I.T.H., and the NIH grant GMS RO1-54045 to V.N.R..
(14) 314. Human Molecular Genetics, 2002, Vol. 11, No. 3. REFERENCES 1. Pierce,J.G. and Parsons,T.F. (1981) Glycoprotein hormones: structure and function. Annu. Rev. Biochem., 50, 465–495. 2. Green,E.D. and Baenziger,J.U. (1998) Asparagine-linked oligosaccharides on lutropin, follitropin, and thyrotropin. Distribution of sulfated and sialylated oligosaccharides on bovine, ovine and human pituitary glycoprotein hormones. J. Biol. Chem., 263, 36–44. 3. McFarland,K.C., Sprengel,R., Phillips,H.S., Kolher,M., Rosemblit,N., Nicolics,K., Segaloff,D.L. and Seeburg,P.H. (1989) Lutropin/ choriogonadotropin receptor: an unusual member of the G protein-coupled receptor family. Science, 245, 494–499. 4. Camp,T.A., Rahal,J.O. and Mayo,K.E. (1991) Cellular localization and hormonal regulation of follicle-stimulating hormone and luteinizing hormone receptor messenger RNAs in the rat ovary. Mol. Endocrinol., 5, 1405–1417. 5. Segaloff,D.L. and Ascoli,M. (1993) The lutropin/choriogonadotropin receptor…4 years later. Endocr. Rev., 14, 324–347. 6. Zhang,F.-P., Rannikko,A.S., Manna,P.R., Fraser,H.R. and Huhtaniemi,I.T. (1997) Cloning and functional expression of the luteinizing hormone receptor complementary deoxyribonucleic acid from the marmoset monkey testis: Absence of sequences encoding exon 10 in other species. Endocrinology, 138, 2481–2490. 7. Gudermann,T., Birnbaumer,M. and Birnbaumer,L. (1992) Evidence for dual coupling of the murine luteinizing hormone receptor to adenylate cyclase and phosphoinositide breakdown and Ca2+ mobilization. Studies with the cloned murine luteinizing hormone receptor expressed in L cells. J. Biol. Chem., 267, 4479–4488. 8. Cameron,M.R., Foster,J.S., Bukovsky,A. and Wimalasena,J. (1996) Activation of mitogen-activated protein kinases by gonadotropins and cyclic adenosine 5′-monophosphates in porcine granulosa cells. Biol. Reprod., 55, 111–119. 9. Sairam,M.R. (1989) Role of carbohydrates in glycoprotein hormone signal transduction. FASEB J., 3, 1915–1926. 10. Furuhashi,M., Suzuki,S., Tomoda,Y. and Suganuma,N. (1995) Role of the Pro-Leu-Arg motif in glycosylation of human gonadotropin α-subunit. Endocrinology, 136, 2270–2275. 11. Jia,X.-C., Bo,M., Tanaka,T., Ny,T., Boime,I. and Hsueh,A.J.W. (1991) Expression of human luteinizing hormone (LH) receptor: interaction with LH and chorionic gonadotropin from human but not equine, rat, and ovine species. Mol. Endocrinol., 5, 759–768. 12. Baenziger,J.U., Kumar,S., Brodbeck,R.M., Smith,P.L. and Beranek,M.C. (1992) Circulatory half-life but not interaction with the lutropin/chorionic gonadotropin receptor is modulated by sulfation of bovine lutropin oligosaccharides. Proc. Natl Acad. Sci. USA, 89, 334–338. 13. Dufau,M.L. (1988) Endocrine regulation and communicating functions of the Leydig cell. Annu. Rev. Physiol., 50, 483–508. 14. Jameson,J.L. (1996) Inherited disorders of the gonadotropin hormones. Mol. Cell. Endocrinol., 125, 143–149. 15. Themmen,A.P.N. and Huhtaniemi,I.T. (2000) Mutations of gonadotropins and gonadotropin receptors: elucidating the physiology and pathophysiology of pituitary-gonadal function. Endocr. Rev., 21, 551–583. 16. Zhang,F.-P., Poutanen,M., Wilbertz,J. and Huhtaniemi,I. (2001) Normal prenatal but arrested postnatal sexual development of luteinizing hormone receptor knockout (LuRKO) mice. Mol. Endocrinol., 15, 172–183. 17. Lei,Z.M., Mishra,S., Zou,W., Xu,B., Foltz,M., Li,X. and Rao, Ch.-V. (2001) Targeted disruption of luteinizing hormone/human chorionic gonadotropin receptor gene. Mol. Endocrinol., 15, 184–200. 18. Weiss,J., Axelrod,L., Whitecomb,R.W., Harris,P.E., Crowley,W.F. and Jameson,J.L. (1992) Hypogonadism caused by a single amino acid substitution in the β-subunit of luteinizing hormone. N. Engl. J. Med., 326, 179–183. 19. Furui,K., Suganuma,N., Tsukahara,S., Asada,Y., Kikkawa,F., Tanaka,M., Ozawa,T. and Tomoda,Y. (1994) Identification of two point mutations in the gene encoding luteinizing hormone (LH) β-subunit, associated with immunologically anomalous LH variants. J. Clin. Endocrinol. Metab., 78, 107–113. 20. Pettersson,K., Mäkelä,M.M., Dahlén,P., Lamminen,T., Huoponen,K. and Huhtaniemi,I. (1994) Genetic polymorphism found in the LH β-gene of an immunologically anomalous variant of human luteinizing hormone. Eur. J. Endocrinol., 130 (Suppl. 2), 65. 21. Okuda,K., Yamada,T., Imoto,H., Komatsubara,H. and Sugimoto,O. (1994) Antigenic alteration of an anomalous human luteinizing hormone. 22.. 23.. 24.. 25.. 26.. 27.. 28.. 29.. 30.. 31.. 32.. 33.. 34.. 35.. 36.. 37.. 38.. 39.. caused by two chorionic gonadotropin-type amino-acid substitutions. Biochem. Biophys. Res. Commun., 200, 584–590. Haavisto,A.M., Pettersson,K., Bergendahl,M., Virkamäki,A. and Huhtaniemi,I. (1995) Occurrence and biological properties of a common genetic variant of luteinizing hormone. J. Clin. Endocrinol. Metab., 80, 1257–1263. Nilsson,C., Pettersson,K., Millar,R.P., Coerver,K.A., Matzuk,M.M. and Huhtaniemi,I.T. (1997) World wide frequency of a common genetic variant of luteinizing hormone: an international collaborative research. International Collaborative Research Group. Fertil. Steril., 67, 998–1004. Huhtaniemi,I.T. and Pettersson,K.S.I. (1999) Alterations in gonadal steroidogenesis in individuals expressing a common genetic variant of luteinizing hormone. J. Steroid Biochem. Mol. Biol., 69, 281–285. Suganuma,N., Furui,K., Furuhashi,M., Asada,Y., Kikkawa,F. and Tomoda,Y. (1995) Screening of the mutations in the luteinizing hormone β-subunit in patients with menstrual disorders. Fertil. Steril., 63, 989–995. Raivio,T., Huhtaniemi,I., Anttila,R., Siimes,M.A., Hagenäs,L., Nilsson,C., Pettersson,K. and Dunkel, L, (1996) The role of luteinizing hormone- β gene polymorphism in the onset and progression of puberty in healthy boys. J. Clin. Endocrinol. Metab., 81, 3278–3282. Cramer,D.W., Pettersson,K.S., Barbieri,R.L. and Huhtaniemi,I.T. (2000) Reproductive hormones, cancers, and conditions in relation to a common genetic variant of luteinizing hormone. Hum. Reprod., 15, 2103–2107. Rajkhowa,M., Talbot,J.A., Jones,P.W., Pettersson,K., Haavisto, A.-M., Huhtaniemi,I. and Clayton,R.N. (1995) Prevalence of an immunological LH β-subunit variant in a UK population of healthy women and women with polycystic ovary syndrome. Clin. Endocrinol., 43, 297–303. Tapanainen,J.S., Koivunen,R., Fauser,B.M.J., Taylor,A.E., Clayton,R.N., White,D., Franks,S., Anttila,L., Pettersson,K. and Huhtaniemi,I.T. (1999) A new contributing factor to polycystic ovary syndrome: The genetic variant form of luteinizing hormone. J. Clin. Endocrinol. Metab., 84, 1711–1715. Ramanujam,L.N., Liao,W.X., Roy,A.C., Loganath,A., Goh,H.H. and Ng,S.C. (1999) Association of molecular variants of luteinizing hormone with menstrual disorders. Clin. Endrocrinol., 51, 243–246. Suganuma,N., Furui,K., Kikkawa,F., Tomoda,Y. and Furuhashi,M. (1996) Effects of the mutations (Trp8→Arg and Ile15→Thr) in human luteinizing hormone (LH) β-subunit on LH bioactivity in vitro and in vivo. Endocrinology, 137, 831–838. Smith,P.L., Kaetzel,D., Nilson,J.H. and Baenziger,J.U. (1990) The sialylated oligosaccharides of recombinant bovine lutropin modulate hormone bioactivity. J. Biol. Chem., 265, 874–881. Hakola,K., Van der Boogaart,P., Mulders,J., de Leeuw,R., Schoonen,W., Van Heyst,J., Swolfs,A., Casteren,J.V., Huhtaniemi,I. and Kloosterboer,H. (1997) Recombinant rat luteinizing hormone; production by Chinese hamster ovary cells, purification and functional characterization. Mol. Cell. Endocrinol., 128, 47–56. Green,E.D., Boime,I. and Baenziger,J.U. (1986) Differential processing of Asn-linked oligosaccharides on pituitary glycoprotein hormones: implications for biologic function. Mol. Cell. Biochem., 72, 81–100. Amoresano,A., Siciliano,R., Orru,S., Napoleoni,R., Altarocca,V., DeLuca,F., Sirna,A. and Pucci,P. (1996) Structural characterization of human recombinant glycohormones follitropin, lutropin and choriogonadotropin expressed in Chinese hamster ovary cells. Eur. J. Biochem., 242, 608–618. Smith,P.L., Skelton,T.P., Dharmesh,S.M., Beranek,M.C., MacPhail,L., Broze,G.J.,Jr and Baenziger,J.U. (1992) The asparagine-linked oligosaccaharides on tissue factor pathway inhibitor terminate with SO4-4GalNAc β 1, 4GlcNAc β 1,2 Mana α. J. Biol. Chem., 267, 19140–19146. Grossmann,M., Szkudlinski,M.W., Tropea,J.E., Bishop,L.A., Thotakura,N.R., Schofield,P.R. and Weintraub,B.D. (1995) Expression of human thyrotropin in cell lines with different glycosylation patterns combined with mutagenesis. Characterization of a novel role for oligosaccharides in the in vitro and in vivo bioactivity. J. Biol. Chem., 270, 29378–29385. Green,E.D., Morishima,C., Boime,I. and Baenziger,J.U. (1985) Structural requirements for sulfation of aspergine-linked oligosaccharides of lutropin. Proc. Natl Acad. Sci. USA, 82, 7850–7854. Joshi,L., Murata,Y., Wondisford,F.E., Szkudlinski,M.W., Desai,R. and Weintraub,B.D. (1995) Recombinant thyrotropin containing a β-subunit chimera with the human chorionic gonadotropin-β carboxy-terminus is biologically active, with a prolonged plasma half-life: role of.
(15) Human Molecular Genetics, 2002, Vol. 11, No. 3. 40. 41.. 42.. 43.. 44.. 45.. 46.. 47. 48.. 49.. 50.. 51.. 52.. 53.. 54.. 55.. 56.. 57.. carbohydrate in bioactivity and metabolic clearance. Endocrinology, 136, 3839–3848. Gharib,S.D., Wierman,M.E., Shupnik,M.A. and Chin,W.W. (1990) Molecular biology of the pituitary gonadotropins. Endocr. Rev., 11, 177–199. Catt,K.J. and Dufau,M.L. (1991) Gonadotropic hormones: Biosynthesis, secretion, receptors, and actions. In Yen,S.S.C. and Jaffe,R. (eds), Reproductive Endocrinology, 3rd edn. W.B. Saunders Co., Philadelphia, PA, pp. 105–155. Perez Mayorga,M., Gromoll,J., Behre,H.M., Gassner,C., Nieschlag,E. and Simoni,M. (2000) Ovarian response to follicle-stimulating hormone (FSH) stimulation depends on the FSH receptor genotype. J. Clin. Endocrinol. Metab., 85, 3365–3369. Matzuk,M.M., Spangler,M.M., Camel,M., Suganuma,N. and Boime,I. (1989) Mutagenesis and chimeric genes define determinants in the β-subunits of human chorionic gonadotropin and lutropin for secretion and assembly. J. Cell Biol., 109, 1429–1438. Kaetzel,D.M., Browne,J.K., Wondisford,F., Nett,T.M., Thomason,A.R. and Nilson,J.H. (1985) Expression of biologically active luteinizing hormone in Chinese hamster ovary cells. Proc. Natl Acad. Sci. USA, 82, 7280–7283. Talbot,J.A., Mitchell,R., Hoy,A.M., Lambert,A., Gauher,A., Hilton,B., McLoughlin,J.D. and Robertson,W.R. (1996) Recombinant human luteinizing hormone: a partial physiochemical, biological and immunological characterization. Mol. Hum. Reprod., 2, 799–806. Jiang,M., Pakarinen,P., Zhang, F.-P., El-Hefnawy,T., Koskimies,P., Pettersson,K. and Huhtaniemi,I. (1999) A common polymorphic allele of the human luteinizing hormone β-subunit gene: additional mutations and differential function of the promoter sequence. Hum. Mol. Genet., 8, 2037–2046. Stocco,D.M. (2001) StAR protein and the regulation of steroid hormone biosynthesis. Annu. Rev. Physiol., 63, 193–213. Suzuki,S., Furuhashi,M. and Suganuma,N. (2000) Additional N-glycosylation at Asn (13) rescues the human LH β-subunit from disulfide-linked aggregation. Mol. Cell. Endocrinol., 160, 157–163. Matzuk,M.M., Keene,J.L. and Boime,I. (1989) Site specificity of the chorionic gonadotropin N-linked oligosaccharides in signal transduction. J. Biol. Chem., 264, 2409–2414. Kaetzel,D.M., Vergin,J.B., Clay,C.M. and Nilson,J.H. (1989) Disruption of N-linked glycosylation of bovine luteinizing hormone β-subunit by site-directed mutagenesis dramatically increases its intracellular stability but does not affect biological activity of the secreted heterodimer. Mol. Endocrinol., 3, 1765–1774. Pettersson,K., Ding,Y.-Q. and Huhtaniemi,I. (1992) An immunologically anomalous luteinizing hormone variant in a healthy woman. J. Clin. Endocrinol. Metab., 74, 164–171. Kalyan,N.K. and Bahl,O.P. (1983) Role of carbohydrate in human chorionic gonadotropin: effect of deglycosylation on the subunit interaction and its in vitro and in vivo biological properties. J. Biol. Chem., 258, 67–74. Skzudlinski,M.W., Thotakura,N.R., Bucci,I., Joshi,L.R., Tsai,A., East Palmer,J., Shiloach,J. and Weintraub,B.D. (1993) Purification and characterization of recombinant human thyrotropin (TSH) isoforms produced by Chinese hamster ovary cells: the role of sialylation and sulfation in TSH bioactivity. Endocrinology, 108, 1490–1503. Fiete,D., Srivastava,V., Hindsgaul,O. and Baenziger,J.U. (1991) A hepatic reticuloendothelial cell receptor specific for SO4-4GalNAc β 1,4GlcNAc β 1,2Man α that mediates rapid clearance of lutropin. Cell, 67, 1103–1110. Kamerling,J.P., Rijkse,I., Maas,A.A., van Kuik,J.A. and Vliegenthart,J.F. (1988) Sulfated N-linked carbohydrate chains in porcine thyroglobulin. FEBS Lett., 241, 246–250. Freeze,H.H., Willies,L., Hamilton,S. and Koza-Taylor,P. (1989) Two mutants of Dictyostelium discoideum that lack a sulfated carbohydrate antigenic determinant synthesize a truncated lipid-linked precursor of N-linked oligosaccharides. J. Biol. Chem., 264, 5653–5659. Spiro,R.G. and Bhoyroo,V.D. (1988) Occurrence of sulfate in the asparagine-linked complex carbohydrate units of thyroglobulin. Identification and localization of galactose 3-sulfate and. 58.. 59.. 60.. 61. 62.. 63.. 64.. 65.. 66.. 67.. 68.. 69.. 70.. 71.. 72.. 73. 74.. 75.. 315. N-acetylglucosamine 6-sulfate residues in the human and calf proteins. J. Biol. Chem., 263, 14351–14358. Hoogewerf,A.J. and Bensadoun,A. (1991) Occurrence of sulfate in an asparagine-linked complex oligosaccharide of chicken adipose lipoprotein lipase. J. Biol. Chem., 266, 1048–1057. Mattews,C.H., Borgato,S., Beck-Peccoz,P., Adams,M., Tone,Y., Gambino,G., Casagrande,S., Tedeschini,G., Benedetti,A. and Chatterjee,V.K.K. (1993) Primary amenorrhoea and infertility due to a mutation in the β-subunit of follicle-stimulating hormone. Nat. Genet., 5, 83–86. Dacou-Voutetakis,C., Feltquate,D.M., Drakopoulou,M., Kourides,I.A. and Dracopoli,N.C. (1990) Familial hypothyroidism caused by a nonsense mutation in the thyroid-stimulating hormone β-subunit gene. Am. J. Hum. Genet., 46, 988–993. Lobo,R.A., Kletzky,O.A. and DiZerega,G.S. (1982) Elevated serum bioactive LH concentrations in women with PCO. Fertil. Steril., 37, 301–302. Liu,G., Duranteau,L., Carel,J.C., Monroe,J., Doyle,D.A. and Shenker,A. (1999) Leydig-cell tumors caused by an activating mutation of the gene encoding the luteinizing hormone receptor. N. Engl. J. Med., 341, 1731–1736. Matzuk,M.M., Krieger,M., Corless,C.L. and Boime,I. (1987) Effects of preventing O-glycosylation on the secretion of human chorionic gonadotropin in Chinese hamster ovary cells. Proc. Natl Acad. Sci. USA, 84, 6354–6358. DeFeo,D., Gonda,M.A., Young,H.A., Cheng,E.H., Lowy,D.R., Scolnick,E.M. and Ellis,R.W. (1981) Analysis of two divergent rat genomic clones homologous to the transforming gene of Harvey sarcoma virus. Proc. Natl Acad. Sci. USA, 78, 3328–3332. Matzuk,M.M. and Boime,I. (1988) The role of aspergine-linked oligosaccharides of the α-subunit in the secretion and assembly of human chorionic gonadotropin. J. Cell Biol., 106, 1049–1059. Pierroz,D.D., Aebi,A.C., Huhtaniemi,I.T. and Aubert,M.L. (1999) Many LH peaks are needed to physiologically stimulate testosterone secretion: modulation by fasting and NPY. Am. J. Physiol., 276, E603–E610. Chomczynski,P. and Sacchi,N. (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem., 162, 156–159. Manna,P.R., Tena-Sempere,M. and Huhtaniemi,I.T. (1999) Molecular mechanisms of thyroid hormone-stimulated steroidogenesis: Involvement of steroidogenic acute regulatory (StAR) protein. J. Biol. Chem., 274, 5909–5918. Manna,P.R., Pakarinen,P., El-Hefnawy,T. and Huhtaniemi,I.T. (1999) Functional assessment of the calcium messenger system in cultured mouse Leydig tumor cells: Regulation of human chorionic gonadotropin-induced expression of the steroidogenic acute regulatory protein. Endocrinology, 140, 1739–1751. Tena-Sempere,M., Manna,P.R. and Huhtaniemi,I. (1999) Molecular cloning of the mouse follicle-stimulating hormone receptor complementary deoxyribonucleic acid: Functional expression of alternatively spliced variants and receptor inactivation by a C566T transition in exon 7 of the coding sequence. Biol. Reprod., 60, 1515–1527. Reinhold,B.B., Hauer,C.R., Plummer,T.H. and Reinhold,V.N. (1995) Detailed structural analysis of a novel, specific O-linked glycan from the prokaryote Flavobacterium meningosepticum. J. Biol. Chem., 270, 13197–13203. Stroud,M.R., Handa,K., Salyan,M.E.K., Ito,K., Levery,S.B., Hakomori,S., Reinhold,B.B. and Reinhold,V.N. (1996) Monosialogangliosides of human myelogenous Leukemia HL60 cells and normal human leukocytes. Characterization of E-selectin binding fractions, and structural requirements for physiological binding to E-selectin. Biochemistry, 35, 770–778. Rebois,R.V. (1982) Establishment of gonadotropin-responsive murine Leydig tumor cell line. J. Cell Biol., 94, 70–76. Harper,J. and Brooker,G. (1975) Femtomol sensitive radioimmunoassay for cyclic-AMP and cyclic-GMP after 2′-O-acetylation by acetic anhydride in aqueous solution. J. Cyclic Nucleotide Res., 4, 207–218. Vuorento,T., Lahti,A., Hovatta,O. and Huhtaniemi,I.T. (1989) Daily measurements of salivary progesterone reveal a high rate of anovulation in healthy students. Scand. J. Clin. Lab. Invest., 49, 395–401..
(16) 316. Human Molecular Genetics, 2002, Vol. 11, No. 3.
(17)
Figure
+5
![Figure 8. Displacement of [ 125 I]iodo-hCG binding with increasing concentrations of the rec LHs and hCG](https://thumb-eu.123doks.com/thumbv2/123doknet/14890250.648886/6.892.94.398.143.449/figure-displacement-iodo-hcg-binding-increasing-concentrations-lhs.webp)

Documents relatifs