Doctoral College:
Life and Health Sciences
DOCTORAL THESIS
Presented by
Ilias MARMOUZI
Speciality: Medical Biology, Human and Experimental
Pathology, and Environment
Laboratory: Pharmacology and Toxicology
Theme
Nutritional Characteristics, Biochemical Composition
and Pharmacological Properties of Moroccan Oat
Varieties and their Endophyte Chaetomium Globosum
Publicly defended by the 31/10/2017 in the presence of the jury:
Prof. Yahia CHERRAH Mohammed V University in Rabat-FMPR President
Prof. Abdelaziz BENJOUAD International University of Rabat-UIR Reporter Prof. Mourad ERRASFA University Sidi Mohamed Ben Abdellah, Fès-USMBA Reporter
Prof. Katim ALAOUI Mohammed V University in Rabat -FMPR Reporter
Prof. Youssef BAKRI Mohammed V University in Rabat-FSR Examinator
Prof. Abdelhalim MESFIOUI University Ibn Tofail, Kénitra-FSK Examinator Prof. My El Abbes FAOUZI Mohammed V University in Rabat-FMPR Supervisor
MOHAMMED V UNIVERSITY IN RABAT
FACULTÉ DE MÉDECINE ET DE PHARMACIE
À ma chére épouse Aisha, en témoignage de mon amour et mon affection (Error).
À mes chers parents (El Arbi Marmouzi & Malika Ben Dehhane), Pour ces longues années de soutien inconditionnel, pour leur confiance permanente. Pour tous les sacrifices et les encouragements qu’ils ont consentis pour moi, pour que nous puissions poursuivre nos études. J’en suis conscient et très reconnaissant.
À mon frère Salaheddine et ma sœur Samia, En témoignage de mon affection fraternelle, de ma profonde tendresse et reconnaissance, je vous souhaite une vie pleine de bonheur et de succès. Que vos rêves soient en tous exaucés.
À mes grands-mères Fatna El ouad et Aïcha Blanchit et ma tante Fatima Ben Dehhane, Qui m’ont accompagné par leurs prières, douceur, que Dieu, le tout puissant, vous protège et vous garde. À la mémoire de mes grands-parents Allal & Bouaaza, Vous demeurez toujours présents à nos esprits. Tellement vous nous manquez ce jour-là. Aucune dédicace ne pourra traduire notre profond amour et gratitude pour tout l’amour et l’affection que vous nous avez offerts. Que le paradis soit votre céleste demeure.
À la grande famille Marmouzi, À la grande famille Ben Dehhane, À la grande famille Attar,
À mes ami(e)s, particulièrement Ayoub Sabri, Meryem El Jemli, Fatima Lakhdar, Fouzia Mamouch, Abdul-Malik Abudunia, Mourad Kharbach, Anouar Hamda, El Arbi Aziane, Rim Bousalham et Inssaf Berkiks, et à tous ceux qui me sont chers, À toutes les personnes qui ont participé à l’élaboration de ce travail à tous ceux que j’ai omis de citer.
AKNOWLEDGEMENTS
It is a great pleasure to get this opportunity to express my deepest appreciation and gratitude to all those that were directly or indirectly involved in the completion of this doctoral thesis.
My deepest gratitude should go to my supervisor Prof. My El Abbes Faouzi for giving me such a great opportunity to work on this promising field. It has been a real pleasure to work with him. He has always been available to advise me and demand a high quality of work in all my endeavors. He encouraged and expected me to think more independently about our experiments and results. I am very grateful for his patience, motivation, and determination.
I would like to express my cordial thanks and gratitude to Prof. Peter Proksch for giving me the opportunity to pursue my research fellowship at his institute, as well as for his valuable recommendations, his fruitful discussions, his unlimited support and for the excellent working facilities at the Institute of Pharmacological Biology and Biotechnology, Heinrich- Heine-Universität, Düsseldorf.
I would like to deeply thank Prof. Yahia Cherrah for his guidance, his continuous encouragement, and his kind advices during my thesis. I would like to acknowledge Prof. Bouchra Meddah for her contribution in the design and initiation of this research project.
My specials thanks to Dr. Fatima Zahrae Kabbaj, for her constructive advises and help in fungi isolation, and to Dr. Sherif Esayed Ebada, for sharing his expertise in NMR data interpretation.
I am deeply grateful to all members of the jury for agreeing to read the manuscript and to participate in the defense of this thesis. I would like to express my profound gratitude to my professors and colleagues from the Department of Pharmacology, University Mohammed V, Rabat, Morocco, especially Prof. Amina
Zellou, Prof. Abdelaziz Bouklouze, Prof. Katim Alaoui, Prof. Soufiane Derraji and
Prof. Rachid El Jaoudi.
My gratitude and all my regards to Prof. Jamal Taoufik (Director of the Doctoral College “Sciences de la Vie et de la Santé” and Vice Doyen of Pharmacy at the Faculty of Medicine and Pharmacy) for carrying out the success of our educational program. My sincere gratitude and thankfulness to the professors of ongoing seminars at the doctoral college “CedocSVS”; for their coaching, assiduity and their sense of responsibility.
I would like to thank Prof. Werner E. G. Müller, Institute of Physiological Chemistry and Pathobiochemistry, University of Mainz, for carrying out the cytotoxicity
assays, and Prof. Rainer Kalscheuer and Dr. Nidja Rehberg, Institute of Pharmaceutical Biology and Biotechnology, Heinrich-Heine University, for carrying out the antibacterial and antitubercular assays.
Many thanks to technical assistants of : - the Institute of Pharmacological Biology and Biotechnology, Heinrich-Heine-Universität, Düsseldorf (Mrs. Miljanovic Simone, Mrs. Katja Friedrich, Mrs. Waltraud Schlag) who never gave up providing assistance and continuously supplied me with the laboratory equipment and solvents during my laboratory work. - the Laboratory of Pharmacology and Toxicology, University Mohammed V in Rabat (Mrs. Hayat Houari and Mr. Ahmed Lafrouhi), Hassan II Agronomy and Veterinary Institute, Rabat (Mr. Hassan Mounakhil), National Institut for Agricultural Reasearch (Mr. Abdenbi Saadi) for their sense of responsibility and their ongoing assistance.
Many thanks to Dr. Said Gharby, Dr. Mohamed Benzine, Dr. Malika Haddam
and Dr. Hicham Harhar for Oil analysis, Dr. Miloud El Karbane and Dr. Mourad Kharbach for chemical analysis, Dr. Hanae Ouaddari for mineral analysis, Dr. Karym El Mostafa for cell cultures, Prof. Mohammed Achaaban for advices, guidance and help
with dissection, Dr. Laila Jahidi Ghazali Laboratory for histological experiments and comments, Dr. Ali Berraaouan for pancreas protocols, Prof. Fatima Affifi and Prof.
Violet Kasabri for enzymes protocols, Dr. Fatima Lakhdar, Dr. Nadia El Madani and
Dr. Abdellatif Bouayyadi for providing reagents and Dr Taha Kamli for rheological measurement.
I would like to extend my thanks to my past and present colleagues and lab-mates and all the others for the nice multicultural time I spent with them, for their help and assistance whenever I needed it: Bouchra Faridi, Rini Muharini, Tatyana Medvedeva, Chong Dat Pharm, Hendrik Niemann, Artha Kuci, Catalina Peres Hemphil, Maha El Hamdani, Huiqin Chen, Antonius Ola, Dhana Tommy, Yang Liu, Mi Young Chung, Lena Hammerschmidt, Andreas Marmann, Mustapha El Amrani, Imke Form, Robert Bara, Noureddine Touach, Karima Sayah, Nadjat Bouchmaa, Imane Arous, Kaddafi Ali, Anas Lakhouit, Laila Chemlal and Mohammed Kaabaoui. Especially, I treasure my time with Meryem El Jemli, Sabrina Keil, Nihal Aktas, Mariam Moussa, Georgios Daletos, Gideon Alade and Sergi Akone for the nice and productive period I spent with them.
FORWARD
This thesis started as a collaborative project between the Pharmacology and Toxicology Laboratory of Faculty of Medicine and Pharmacy- Rabat, and the National Institut of Agricultural Research. The project is financially supported by the CNRST (Centre National de Recherche Scientifique et Technique, Morocco (RS/2011/27)). The main part of this thesis was done at the Pharmacology and
Toxicology Laboratory. Other parts of this work were prepared at the following departments:
Institut für Pharmazeutische Biologie und Biotechnologie, Heinrich-Heine-Universität Düsseldorf, Germany.
Unité d'Anatomie Comparée-CNRST-URAC 49, Institut Agronomique et Vétérinaire Hassan II, Rabat, Morocco.
Animalerie Centrale de la Faculté de Médecine et de Pharmacie de Rabat, Morocco.
Service de Physicochimie, Laboratoire de Controle de Qualité de Médicament, division de Médicament et Pharmacie, Morocco.
I gratefully acknowledge the funding sources received towards my PhD from DAAD - Deutscher Akademischer Austauschdienst (Service allemand d’échange académique).
Special thanks should also be given to Prof. Peter Proksch of University of Düsseldorf and Prof. Khalid El Allali, Hassan II institut. Thank you for always making me feel so welcome in your labs, for your encouragement and supervisory role.
LIST
OF
PUBLICATIONS
Thesis publications
Ilias Marmouzi, El Mostafa Karym, Nezha Saidi, Bouchra Meddah, Mourad Kharbach, Azlarab Masrar, Mounya Bouabdellah, Layachi Chabraoui, Khalid El Allali, Yahia Cherrah, My El Abbes Faouzi. In vitro and in vivo antioxidant and anti-hyperglycemic activities of Moroccan Oat cultivars. Antioxidants 2017, 6(4), 102; doi:10.3390/antiox6040102.
Ilias Marmouzi, My El Abbes Faouzi, Nezha Saidi, Yahia Cherrah, Nidja Rehberg, Sherif S. Ebada, Weaam Ebrahim, Rainer Kalscheuer, Peter Proksch. Bioactive Secondary Metabolites from Chaetomium globosum, an Endophyte from the Moroccan Plant
Avena sativa. Chemistry of Natural compounds, 2017, 53: 1208;
doi:10.1007/s10600-017-2242-6.
Ilias Marmouzi, Samira Serragui, Nezha Saidi, Mohammed Bouksaim, Yahia Cherrah, My El Abbes Faouzi. Effect of Drying, Polarity and Molecular Weight on Oat Chemical content and Bioactivity. Journal of Biologically Active Products from Nature, 2017, 331-340.
Ilias Marmouzi, Nezha Saidi, Bouchra Meddah, Mohammed Bouksaim, Said Gharby, Miloud El Karbane, Samira Serragui, Yahia Cherrah, My El Abbes Faouzi. Nutritional characteristics, biochemical composition and antioxidant activities of Moroccan Oat varieties. Journal of Food Measurement and Characterization, 2016, 10, 1, 156-165.
Data
New secondary metabolites of Chaetomium globosum. Ilias Marmouzi, Fatima Zahra Kabbaj, Andréas Marmann, Bouchra Meddah, Nezha Saidi, Yahia Cherrah, Abdessamad Debbab, My El abbes Faouzi and Peter Proksch. Chaetomium globosum genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, strain TR3. GenBank: HG423571.1
LIST OF COMMUNICATIONS
Oral communications
29 May-1June 2016: Ilias Marmouzi, El Mostafa Karym, Nezha Saidi, Bouchra Meddah, Mohammed Bouksaim, Azlarab Masrar, Mounya Bouabdellah, Layachi Chabraoui, Khalid El Allali, Yahia Cherrah, My El Abbes Faouzi. Antihyperglycemic and antioxidant effect of new Moroccan hybrid oat in diabetic model, and inhibitory effect on digestive enzymes. CIPAM 2016 - 6th International Congress of Aromatic and Medicinal Plants, Coimbra, Portugal.
10-10 March 2016: Ilias Marmouzi, My El Abbes Faouzi, Nezha Saidi, Yahia Cherrah, Nidja Rehberg, Sherif S. Ebada, Weaam Ebrahim, Rainer Kalscheuer, Peter Proksch. Antitubercular secondary metabolites from Chaetomium globosum. 5th
Spanish-Moroccan Symposium on Organic Chemistry, Casablanca, Morocco.
15-16 December 2014: Ilias Marmouzi, Nezha Saidi, Bouchra Meddah, Mohammed Bouksaim, Yahia Cherrah, My El Abbes Faouzi. Effect of hybridization on the chemical composition, antioxidant and antihyperglycemic properties in new Moroccan oat lines. Premier Congrès International Substances Naturelles & Modélisation: Applications Thérapeutiques, Environnementales & Développement Durable, Taza, Morocco.
20-21 December 2013: Ilias Marmouzi, Bouchra Meddah, Mohamed Bouksim, Nezha Saidi, Yahia Cherrah, Abdessamad Debbab, My El Abbes Faouzi. Phytochemical content, antioxidant activity and antihyperglycemic effect of two morrocan oat varities. Deuxieme Journées Jeune Chercheurs - Société Marocaine de Chimie Thérapeutique, Kénitra, Morocco.
5-7 December 2013: Ilias Marmouzi, Nezha Saidi, Bouchra Meddah, Fatima Zahra Kabbaj, Peter Proksch, Yahia Cherrah, Abdessamad Debbab, My El Abbes Faouzi. Cytotoxic epipolythiodoxopiperazines derivatives from chaetomium globosum. The third annual meeting of the Middle Eastern association for cancer research “Cancer science from bench to clinic”, Rabat, Morocco.
14-16 November 2013: Ilias Marmouzi, Nezha Saidi, Bouchra Meddah, Peter Proksch, Yahia Cherrah, Fatima Zahra Kabbaj, Abdessamad Debbab, My El Abbes Faouzi. Cytotoxic secondary metabolites from the endophytic fungus Chaetomium
globosum isolated from Avena sativa growing in Morocco. Symposium international
de plante aromatique et médicinale. (SIPAM5) Marrakech, Morocco.
Posters
13 - 14 May 2016: Ilias Marmouzi, Bouchra Meddah, Nezha Saidi, Yahia Cherrah, My El Abbes Faouzi. Oat Phytochemistry and Antidiabetic Activity. Colloque National sur le Diabète et la Nutrition, Faculty of Medecine and Pharmacy, Rabat, Morocco.
20-21 December 2013: Ilias Marmouzi, Nezha Saidi, Bouchra Meddah, Fatima Zahra Kabbaj, Peter Proksch, Yahia Cherrah, My El Abbes Faouzi, Abdessamad Debbab. Metabolites secondaires d’un champignon endophyte de l’avoine. Deuxième Journées Jeune chercheurs - Société Marocaine de Chimie Thérapeutique, Kénitra, Morocco.
14-16 November 2013: Ilias Marmouzi, Nezha Saidi, Bouchra Meddah, Mohamed Bouksim, Nezha saidi, Samira Serragui, Yahia Cherrah, Fatima Zahra Kabbaj, Abdessamad Debbab, My El Abbes Faouzi. Antioxidant activity of Oat methanolic extracts and fractions. SIPAM5, Marrakech, Morocco.
SUMMARY
GENERAL INTRODUCTION 5
LITERATURE REVIEW 7
UNIT 1. PHYSIOPATHOLOGY OF DIABETES AND OXIDATIVE STRESS 8
1. Glucose homeostasis and regulation 9
1.1. Pancreatic regulation of glucose 9
2. Defects in glucose homeostasis 15
2.1. Diagnosis of hyperglycaemia 15
2.2. Physiopathology of hyperglycemia 16
3. Diabetes and Oxidative stress 17
3.1. Oxidative stress 17
3.2. Glucose induced toxicity and oxidative stress 17
UNIT 2. FOOD INTAKE AND GLUCOSE REGULATION 20
1. Food antioxidants and glucose regulation 21
2. Oat compounds and glucose regulation 22
UNIT 3. BIOACTIVE SECONDARY METABOLITES FROM PLANT’S
ENDOPHYTES 24
1. Endophytes chemodiversity 25
2. Natural products isolation from endophytes 26
3. Plant endophyte interactions 26
CONTEXT & OBJECTIVES 28
PLANT MATERIAL 29
STATISTICAL ANALYSIS 29
EXPERIMENTAL PART 30
CHAPTER 1. OAT CHEMICAL COMPOSITION 31
I. Introduction 32
II.Material and methods 32
1. Proximate analysis 32
2. Scanning electron microscopy (SEM) 32
3. Rheological measurement 33
4. Mineral content 33
5. Fatty acids and sterols analysis 33
6. HPLC-RI of Sugars content 34
7. HPLC-FD 34
8. Spectrophotometric and HPLC-MS analysis of phenolic compounds 35
2
CHAPTER 2. OAT ANTIOXIDANT EFFECT 45
I. Introduction 46
II.Material and methods 47
1. Free radical scavenging activity by DPPH 47
2. Trolox equivalent antioxidant capacity 47
3. Determination of reducing/antioxidant power (FRAP) 47
4. Antioxidant effect in Tetrahymena pyriformis cell culture 48
III. Results and discussion 49
CHAPTER 3. OAT IN VITRO ANTIDIABETIC ACTIVITY 52
I. Introduction 53
II.Material and methods 54
1. α-amylase inhibition assay 54
2. α-glucosidase inhibition assay 54
3. Inhibitory Kinetics 55
III. Results 55
IV. Discussion 56
CHAPTER 4. OAT IN VIVO ANTIDIABETIC AND ANTIOXIDANT ACTIVITIES 58
I. Introduction 59
II.Material and methods 60
1. Animals 60
2. Acute Toxicty 60
3. Experimental diabetes 60
4. Experimental design 61
5. Oral glucose tolerance test 61
6. Behavioral assays 62
7. Hematological and biochemical analysis 62
8. Key enzymes and markers of oxidative stress 62
9. Histological study of pancreas 65
III. Results 66
IV. Discussion 72
CHAPTER 5. SECONDARY METABOLITES OF OAT ENDOPHYTIC FUNGUS
CHAETOMIUM GLOBOSUM 77
I. Introduction 78
II.Material and methods 78
1. Pure fungal strain isolation 78
2. Taxonomic identification 79
3 4. Extraction and fractionation of fungi grown on solid rice medium 80
5. Isolation of secondary metabolites 81
6. Structure elucidation of the isolated metabolites 83
7. Bioassays of isolated compounds 86
III. Results and discussion 88
CONCLUSIONS AND PERSPECTIVES 98
REFERENCES 100
4
TABLES
Table 1. Antidiabetic mechanisms of Food ingredients 21
Table 2. Selected publications on preclinical antidiabetic activities of Oat derived compounds
23
Table 3. Proximate analysis of Moroccan Oat varieties 36
Table 4. Viscosity of hybrid and parent Oat varities 38
Table 5. Mineral composition in mg/Kg 39
Table 6. Fatty acids composition (%) of Moroccan Oat varieties 40
Table 7. Sterols content (%) of Moroccan Oat varieties 41
Table 8. Sugar contents in g/Kg dw by HPLC-RI method 42
Table 9. α-tocopherol contents in mg/Kg dw by HPLC-FD method 42
Table 10. Phenolic and flavonoid contents of Oat varieties 43
Table 11. Phenolic composition of hybrid Oats and parent (Amlal) in mg/g 44
Table 12. Antioxidant activity of Moroccan Oat varities 50
Table 13. Changes in fasting blood glucose and metabolic parameters 68
Table 14. Hematological and biochemical analysis of diabetic animals 69
Table 15. Levels of antioxidant enzymes and oxidative stress markers in liver and kidney
70
Table 16. Behavioral analysis using OFT and EPM 72
Table 17. Composition the malt agar medium 79
Table 18. Analytical HPLC parameters 83
Table 19. Semi-preparative HPLC parameters 83
Table 20. ESIMS parameters 85
Table 21. Antitubercular and Antibiotic Activities of the Active Compound Chetomin (MIC in μM)
90
Table 22. Cytotoxic Activity of Isolated Compounds Against Murine Lymphoma L5178Y cell line.
90
Table 23. H (500 MHz) and 13C NMR (125 MHz) Data of Chetomin (1) and Dethio-tetra (methylthio) Chetomin (2) in (DMSO-d6, δ, ppm, J/Hz)
94
Tablle 24. 1H (500 MHz) and 13C NMR (125 MHz) Data of Chetosemindusins B (3) and C (4) in (CD3OD, δ, ppm, J/Hz)
5
FIGURES
Figure 1. Anatomical and functional organization of the pancreas. 9 Figure 2. The interplay of the pancreas with the brain, liver, gut as well as
adipose and muscle tissue
12
Figure 3. Principles mechanisms leading to hyperglycemia 16 Figure 4. Hyperglycemia, through different mechanisms 18
Figure 5. Oat seeds morphology 22
Figure 6. Chemical structure of the anticancer drug Taxol 25 Figure 7. Steps to isolate natural products from biological material 26 Figure 8. Beneficial properties of endophytes 27
Figure 9. Surface morphology of Oat starch 38
Figure 10. HPLC-RI chromatograms of sugars 41
Figure 11. HPLC-FD chromatogram of α-tocopherol in Oat extract. 42 Figure 12. HPLC-MS chromatogram of phenolic compounds in Oat extract 43 Figure 13. Oat protective effect on antioxidant enzymes under oxidative
stress in T. pyriformis.
51
Figure 14. α-amylase and α-glucosidase inhibitory activities 55 Figure 15. Oral glucose tolerance before and after treatment period 67 Figure 16. Effect of diabetes and Oat treatment on histological alterations in
the pancreas
72
Figure 17. Pure fungal strain of Chaetomium globosum on a petri dish 79 Figure 18. Large scale of the endophytic fungus on rice medium. 81 Figure 19. Chromatogramm of the ethyl acetate extract of Chaetomium
globosum.
89
6
ABBREVIATIONS
AD: Alzheimer diseaseADF: Acid detergent fibre ADL: lignin content
AGEs: Advanced glycation end products AI: Atherosclerosis index
AlCl3: Aluminium chloride
ALT: Alanine aminotransferase ANOVA: Analysis of variance
AOAC: Association of Official Analytical Chemists AST: Aspartate aminotransferase
ATF-6: Activating transcription factor 6 ATP: Adenosine triphosphate
AUC: Areas under curve AV: Avenanthramides
BNDF: Brain-derived neurotrophic factor cAMP: Cyclic adenosine monophosphate CAT: Catalase
CCK: Cholecystokinin
CHOP: C/EBP homologous protein DNS: Dinitrosalicylic acid
DPPH: 2,2’-Diphenyl-1-picrylhydrazyl hydrate EI-MS: Electron Impact Mass Spectrometry EPM: Elevated plus maze
ERAD: ER-associated degradation ESIMS: Electron spray ionization mass
spectrometry
EtOAc: Ethyl acetate
FAMEs:fatty acid methyl esters FAs: fatty acids
FBG: Fasting blood glucose
FOXO1: forkhead transcription factor FRAP: ferric ions reducing antioxidant power G-1-P: Glucose-1-phosphate
GIP: Glucose dependent insulinotropic peptide GLP: Glucagon-like peptide
GLUT4: Glucose transporter 4 GPCRs: G-Protein-Coupled Receptors GPx: Glutathione Peroxidase GRP: Gastrin-Releasing Peptide GSH: Glutathione
GSK: Glycogen synthase kinase H2O2: Hydrogen peroxide
HbA1C: Glycated hemoglobin HDL: High density lipoprotein HGB: Hemoglobin
HPLC: high-performance liquid chromatography HR-MS: High-resolution mass spectrometry IC50: 50% inhibitory concentration values
IFG: impaired fasting glucose IGT: impaired glucose tolerance IL-6: Interleukin 6
In: Inoculated
INRA: National Institute for Agricultural
Research
IPF-1: Insulin Promotor Factor 1 IRE-1: inositol-requiring enzyme ITS: internal transcribed spacer JNK: Jun N-terminal kinase,
K+-channels: KATP-channels LDH: Lactate dehydrogenase LDL: Low density lipoprotein
MCH: Melanin concentrating hormone MDA: Malondialdehyde
MIC: Minimal Inhibitory Concentrations MUFA: monounsaturated fatty acids Na2CO3: sodium carbonate
NaNO2: sodium nitrite
NaOH: sodium hydroxide NDF: Neutral detergent fibre NF-kB: Nuclear Factor-Kappa B Ni: noninoculated
NMR: Nuclear magnetic resonance spectroscopy NmU: Neuromedin U
NO: nitric oxide NPY: neuropeptide Y O2. : Superoxide
OECD: Organization of Economic Co-operation
and Development
OFR: Overnight fasted rats OFR: Overnight fasted rats OFT open field test
OGTT: Oral Glucose Tolerance Test OS: Oxidative stress
PACAP: pituitary adenylate cyclase-activating
polypeptide
PBS: phosphate buffer solution PERK: PKR-like ER kinase PKC: Protein kinase C
p-NPG: p-Nitrophenyl-α-D-glucopyranoside
POMC: pro-opiomelanocortin PP: pancreatic polypeptide
PPYG: Proteose-peptone yeast Glucose defined
medium
PUFA: polyunsaturated fatty acids RBC: Red blood corpuscles count RBC: red blood corpuscles ROS: reactive oxygen species SD: standard deviation SM: Sec1/Munc18-like
SNAP-25: synaptosomal-associated protein of
25 kDa
SNAREs: N-ethylmaleimide-sensitive receptor
proteins
SOD: superoxide dismutase
STZ-NA: Streptozotocinenicotinamide T1DM: Type 1 Diabetes Mellitus T2DM: Type 2 Diabetes Mellitus TFC: Total Flavonoids Contents TG: Triacylglycerols
TPC: Total Phenolic Contents UFA: unsaturated fatty acids
VAMP2: Vesicle Associated Membrane Protein VDCCs: Voltage-Dependent Ca+-Channels VIP: Vasoactive Intestinal Peptide VLC: Vacuum liquid chromatography WBC: White Blood Corpuscles
2
ABTRACT
Oat has been recognized as a healthy food for a long time. However, they are still underutilized in Moroccan gastronomy. The current work aimed to evaluate the in vitro and in vivo antidiabetic and antioxidant activities of Moroccan hybrid Oats, and to establish the functional chemical variability among their seeds.
The physicochemical analysis including, minerals, sugars, fatty acids, sterols, and phenolic compounds were investigated. Additionally, starch morphology and Oat viscosity were analyzed. HPLC-MS revealed the presence of eleven phenolic acids, including vannilic, ferulic, hydroxybenzoic, caffeic, salicylic, syringic, sinapic, p-coumaric, caffeic and chlorogenic acids, and pyrogallol. The potassium was the main mineral in Oat seeds; also, the oleic, linoleic, and palmitic acids were the dominant fatty acids. The major sterols were β-sitosterol, Δ-5-avenasterol and campesterol.
Improvement of Oat lines via introgression is an important process for food biochemical functionality. The current work demonstrated the protective effect of aqueous extracts from hybrid Oat line (F11-5) and its parent (Amlal), on hyperglycemia induced oxidative stress and the inhibitory effect on digestive enzyme. The Oat extract (2000 mg/kg) ameliorated the glucose tolerance, decreased FBG and oxidative stress markers (SOD, CAT, GPx, GSH & MDA) in rats liver and kidney. Moreover, Metformin and Oat intake prevented anxiety, hypercholesterolemia and atherosclerosis in diabetic rats. In
vivo anti-hyperglycemic effect of Oat extracts has been confirmed by their inhibitory
activities on α-amylase (723.91 μg/mL & 1027.14 μg/mL) and α-glucosidase (1548.12 μg/mL & 1803.52 μg/mL) enzymes by mean of a mixed inhibition.
Finally, the endophytic fungal strain Chaetomium globosum (HG423571) which was isolated from Oat inner and healthy tissues was subject of secondary metabolites isolation, in order to evaluate their bioactivity.
3
RESUMÉ
Reconnue pour son innocuité et ses bienfaits depuis des decennies, l'avoine est encore sous exploitée en gastronomie marocaine. Le travail actuel visait à évaluer les activités antidiabétiques et antioxydantes in vitro et in vivo des variétés hybrides d’avoine marocaine et à établir la variabilité chimique fonctionnelle entre leurs graines.
L'analyse physico-chimique, y compris les minéraux, les sucres, les acides gras, les stérols et les composés phénoliques a été étudiée. En outre, la morphologie de l'amidon et la viscosité de l'avoine ont été reportées. L’analyse HPLC-MS a révélé la présence de onze acides phénoliques, y compris les acides : vannilique, ferulique, p-hydroxybenzoïque, caféique, salicylique, syringique, sinapique, p-coumarique, chlorogénique et le pyrogallol. Parmi les mineraux de l’avoine, le potassium était majoritaire; aussi, parmi les acides gras : l’oléiques, linoléiques et palmitiques ont dominé. Les stérols majeurs ont été le β-sitostérol, l'Δ-5-avenasterol et le campestérol. L'amélioration des lignes d'avoine par croisement est un processus d'intérêt pour la fonctionnalité biochimique et alimentaire. Le travail actuel a démontré l'effet protecteur des extraits aqueux de la ligne d'avoine hybride (F11-5) et de son parent (Amlal), sur le stress oxydatif induit par l'hyperglycémie et l'effet inhibiteur sur les enzymes digestive. L'extrait d'avoine (2000 mg / kg) a amélioré la tolérance au glucose, les niveaux de glucose à jeun et les marqueurs de stress oxydant (SOD, CAT, GPx, GSH et MDA) au niveau du foie et du rein chez des rats diabétique. En outre, l’administration de la metformine et de l’avoine a pu prévenir le comportement anxieux, l'hypercholestérolémie et l'athérosclérose chez les rats diabétiques. L'effet anti-hyperglycémique in vivo des extraits d'avoine a été confirmé par leurs activités inhibitrices sur l’α-amylase (723.91 μg/mL et 1027.14 μg/mL) et l’α-glucosidase (1548.12 μg / mL et 1803.52 μg/mL) avec un type d’inhibition mixte.
Enfin, Chaetomium globosum (HG423571), un champignon endophytique, isolée à partir de tissus internes et sains d'avoine a été sujet d'isolement de métabolites secondaires. L'objectif de cette étude était d'extraire, d'explorer et de caractériser les produits naturels produits par cet endophyte et d'évaluer ses activités pharmacologiques.
4
ص خل
م
نافوشلا
وأ
يبشع تابن وه لاطرخلا
و
عون
نم
بوبحلا
لما
ورعر
اذغلا هتميقب
،ةيئ
لاإ
هنأ
املق
يف مدختسي
يبرغلما خبطلما
.
دهت
انتسارد
ىلإ
مييقت
ةيلاعرف
ا
مك يبرغلما نافوشل
لم ردص
تاداض
الأ
و ودسك
نم ةياقو
ا
يركسل
و ،
مييقتل كلذك
تاريغتلا
اميكلا
و
ةي
نيب
روذبلا
.
تلا ةسارد للاخ نم
دوجو نيبت يئايميكويزيفلا ليلح
نداعرم
،
تايركس
،
ضامحأ
ةينهد
،
لوريتسوتياف
و
م
تابكر
ةيلونيف
.
لذ ىلإ ةفاضلإاب
مت ،ك
ةظحلام
ا
اشنل
الإ بوكسوركيلما ةطساوب
ينورتكل
،
و
ذك
ى
لدعرم
ةجوزللا
.
يفاركوتاموركلا ليلحتلاب قلعرتي اميفو
HPLC-MS
دقف
أ
ضف
ى
رعرتلل
لع
ى
إ
رشع ىدح
مح
ض
ا
يلونيف
ا
يهو
كيلينافلا
،
كيلريفلا
،
كيوزنب ي سكورديهلا
،
لا
،كيفاك
يسلا ،كيلسلاسلا
رن
ج
ي
،كيبانيسلا ،ك
كيراموكلا
،
كينيجورولكلا
،
و
لولاغوريبلا
.
ظحول امك
أ
ن
م
ندعر
مويساتوبلا
وه
لايثمت رثكالأ
.
لإ ةفاضإ
ى
كيتيملبلاو ،كيلونيللا ،كيلوالأ ضامحأ ،كلذ
،
تناك
روذبلا يف ادجاوت رثكالأ ةينهدلا ضامحالأ
.
أ
يف ام
ام
صخي
لا
لوريتسوتياف
لاف
اتيب
-
لوريتسوتيس
لا و
اتلد
و لوريتسانيفأ
لا
لوريتسبماك
نلجس
أ
لع
ى
بسنلا
.
إ
ةيلمع ن
لا
نيسحت
ينيجلا
ل
نافوشل
لاب
نيجهت
ةماه ةيلمع يه
لا يف
نيوكت
يئايميكويبلا
يفيظولا و
.
سارد
انت
ت حضوت
أ
نيجهلا نافوشلل ةيئالما تاصلختسلما ريث
(F11-5)
و
يربلا
(Amlal)
ع
ل
ى
لا
لماوعر
د ِسكؤلما
ةجتانلا و
نع
إ
مدلا يف ركسلا عافتر
و
ىذك
هريثأت
ا
طبثلما
ل
يمضهلا زاهجلا تاميزن
.
أ
تبث
نافوشلا صلختسم
(
0222
غلم
/
غلك
)
يف هتيلعاف
نيسحت
ايلاخلل زوكلجلا ليمحت
،
ةبسن ضيفخت
مدلاب ركسلا
،
ريثأتلاو
إ
ىلع اباجي
أ
و تاميزن
اتن
ئ
ج
الأ
ودسك
)
(SOD, CAT, GPx, GSH, MDA
لكو دبك يف
ى
نارئفلا
.
لا ةفاضإ
ى
،كلذ
أ
تبث
نافوشلا
و
هتيلعاف نيمروفتيلما
ام
نم ليلقتلا يف
أ
ضارع
إو ،قلقلا
نييارشلا بلصتو لورتسلوكلا عافتر
دنع
ةباصلما نارئفلا
يركسلاب
.
دنع
ةسارد
تاصلختسم طاشن
نافوشلا
ىلع
أ
مضهلا تاميزن
،
لا
طيبثت انضح
تانيتوربلا هذه ةفيظول تابكرلما
،
تناك ثيح
ةيلاعرفلا زيكارت
افلأ يلاتلاك
زيليمأ
(
307.9.
مارغوركيم
(م
ي
غ
)/
و لم
.203..1
م
ي
غ
/
لم
)
و
فلأ
ا-لج
ي
ديزوكو
زا
(
.118..0
م
ي
غ
/
و لم
.827.10
م
ي
غ
/
لم
).
اريخأ
لزع دعربو
ةيرطفلا ةللاسلا
Chaetomium globosum (HG423571)
ةميلسلا ةيلخادلا ةجسنالأ نم
عرضخ نافوشلل
ت
مييقتل ،ةيوناثلا اهتابكرم
لا
طاشن
تا
يويحلا
ة.
تاملكلا حيتافم
:
الأ لماوع، يركسلا ءاد
، نافوشلا، ودسك
توبان يلخاد5
GENERAL
INTRODUCTION
Glucose in chronic high levels can alter or disrupt the functionality and structures organs, including the pancreatic islet. The resulting glucose toxicity can undergo multiple mechanisms of action and biochemical pathways, e.g. glucose autooxidation, protein kinase C activation, methylglyoxal formation and glycation, hexosamine metabolism, sorbitol formation, and oxidative phosphorylation (Robertson, 2004). These pathways share in common the occurence of reactive oxygen species (ROS) that, in excess and over time, results in chronic oxidative stress, and subsequently defective insulin gene expression and insulin secretion as well as increased apoptosis (Robertson, 2004). Glucose toxicity and hyperglycemia is the promoting factor of retinopathy, kidney failure, neuropathies, and macrovascular disease in diabetes (Ceriello, 2006). (Robertson, 2004) suggested that interrupting the overproduction of superoxide by the mitochondrial electron-transport chain would normalize the pathways involved in the development of diabetic complications. In fact, conventional antioxidants scavenge ROS in a stechiometric manner. Food compounds that act as superoxide dismutase (SOD) or catalase (CAT) mimetics have theoriticaly the advantage of scavenging ROS continuously such as native enzymes (Ceriello, 2003). Such compounds normalize endothelial dysfunction and improve diabetes-induced decreases in endoneurial blood flow and motor nerve conduction velocity (Ceriello, 2003). The significance of cereals as a source of unique antioxidant compounds and low-glycemic index diets is expanding (Brand-Miller; Hayne, Petocz, 2003; Wolever et al., 1992). The major cereal grains include wheat, rice, and maize, with oats, rye, barley, buckwheat, sorghum, and millet as minor grains. They are grown for their highly nutritious edible seeds which are often referred to as grains. Grains are composed of starchy endosperm, germ, and bran (including aleurone). Whole grains are important components of the human diet as shown by the inclusion in the Food Guide Pyramid and US Dietary Guidelines (National Research Council, 1989). Whole grains especially cereal grains have been found to be associated in the prevention of Type 2 Diabetes Mellitus (T2DM) (Murtaugh et al., 2003; Venn and Mann, 2004). Among cereals, Oats (Avena sativa L.) contain bioactive
6 components that have a range of positive health benefits, including effects on lipidemic and glycemic control (Jenkins et al., 2002; Ripsin, 1992), as well as satiety (Beck et al., 2009). The Oat soluble fiber may promote satiety, by slowing down digestion resulting in increased gastric retention and feelings of fullness (Jiménez-Cruz et al., 2006). The presence of soluble fiber has also been shown to alter the secretion of gastrointestinal hormones (Behall et al., 2005) and aid body weight regulation (Schuster et al., 2015). Indeed given the high β-glucan content of Oats it could be that release of this polymer significantly alters intestinal viscosity, or has a similar influence on gastric residence time because although energy density affects gastric emptying, it is also affected by viscosity (Camps et al., 2016). In fact the antidiabetic activities of Oat β-glucans were largely established. However, Oats supply more than just fiber. The contributions of the unique antioxidant composition from Oat have not been fully studied. This grain contains numerous bioactive phytochemicals, which are structurally diverse secondary metabolites synthesized by plants (Singh et al., 2013). For example, oats are a good source of phenolic acids, flavonoids, carotenoids, vitamin E, and phytosterols (Bahraminejad et al., 2008; Dimberg et al., 2005; Singh et al., 2013). In addition, Oats produce two unique types of phytochemicals: avenanthramides (AV) (Dokuyucu et al., 2003) and steroidal saponins (Sang and Chu, 2017). Nevertheless, the protective effect of Oat antioxidants against glucose induced oxidative stress is not sufficiently described. On the other hand, improvement of Oat lines via introgression is an important process for food biochemical and pharmacological functionality. Thus, our study investigated the chemical variability in Moroccan hybrid Oat grains compared to a parent variety. Further the in vitro inhibitory effect on digestive enzymes was characterized and the in vivo antioxidant effect in a diabetic model was evaluated. Moreover, numerous plants metabolites were isolated from their endophytic fungi, suggesting a genetic transfer from plants to their endophytic communities. This hypothesis promotes the questions about the contribution of fungal metabolites in plant bioactivities, and weither or not they can constitue alternatives at the industrial level. In this focus, our work aim to investigate the chemical composition of an Oat endophytic fungus (Chaetomium globosum).
7
LITERATURE
REVIEW
- Unit 1. Physiopathology of Diabetes and Oxidative
Stress
Glucose homeostasis and regulation
Defects in glucose homeostasis
- Unit 2. Food intake and Diabetes
Food antioxidants and glucose regulation
Oat compounds and glucose regulation
- Unit 3. Bioactive Secondary Metabolites from
Plants Endophytes
Endophytes chemodiversity
Natural products isolation from endophytes
8
9 1. Glucose homeostasis and regulation
Glucose is an essential nutrient for the human body and a major energy source for many cells. The low blood concentrations of glucose can cause seizures, loss of consciousness, and death. On the other hand, toxicological manifestations such as blindness, renal failure, vascular disease etc, can result from long lasting elevation of glucose concentrations; therefore, blood glucose concentrations need to be maintained within narrow limits at a steady-state level, a process called “glucose homeostasis”(Grayson et al., 2013).
1.1. Pancreatic regulation of glucose
Located behind the stomach within the left upper abdominal cavity and is partitioned into head, body and tail (Röder et al., 2016), the pancreas has key roles in the regulation of macronutrient digestion and hence metabolism/energy homeostasis by releasing various digestive enzymes and pancreatic hormones.
Figure 1. Anatomical and functional organization of the pancreas. The acinar cells mediate the exocrine functions of the pancreas by secreting digestive enzymes into the upper small intestine via the pancreatic duct. Various hormones from the pancreatic islets assure the endocrine functions. Micrograph of pancreatic islets LM× 760 (Röder et al., 2016)
The acinar-or exocrine-cells secrete the pancreatic juice containing digestive enzymes, such as amylase, pancreatic lipase and trypsinogen, into the ducts, that is, the main pancreatic and the accessory pancreatic duct (Figure 1). In contrast, pancreatic hormones are released in an endocrine manner, that is, direct secretion into the blood stream. The islets of langerhans, formed by clustered
10 endocrine cells, is a small island-like structures within the exocrine pancreatic tissue that account for only 1–2% of the entire organ (Chandra and Liddle, 2009). The endocrine cells are: glucagon-producing α-cells (Brissova et al., 2005) (15–20%); amylin-, C-peptide and insulin-producing β-cells (65–80%); pancreatic polypeptide (PP)-producing γ-cells (Katsuura et al., 2002) (3–5%); somatostatin producing δ-cells (3–10%); and ghrelin-producing ε-cells (Wierup et al., 2002) (<1%). Each of the hormones has distinct functions.
Glucagon and insulin plays an opposing and balanced functions in glucose homeostasis; while glucagon increases blood glucose levels, the insulin decreases them (Göke, 2008). Somatostatin inhibits both, glucagon and insulin release (Hauge-Evans et al., 2009), whereas PP regulates the exocrine and endocrine secretion activity of the pancreas (Batterham et al., 2003; Katsuura et al., 2002). Although the islets have a similar cellular composition among different species, that is, human, rat and mouse, their cytoarchitecture differs greatly. Human islets exhibit interconnected α- and β-cells, while in rodents, they are primarily composed of β-cells located in the center with other cell types in the periphery, (Brissova et al., 2005; Cabrera et al., 2006). Through its various hormones, the pancreas maintains blood glucose levels within a very narrow range of 4-6mM.
Hepatic glycogenolysis is promoted by α-cells glucagon when blood glucose levels are low. During prolonged fasting, the hormone drives hepatic and renal gluconeogenesis to increase endogenous blood glucose levels (Freychet et al., 1988). On the opposite, insulin secretion from β-cells is stimulated by elevated exogenous glucose levels (Komatsu et al., 2013). It enables the insulin-dependent uptake of glucose into these muscle and adipose tissues and hence lowers blood glucose levels. Furthermore, it promotes glycogenesis, lipogenesis and the incorporation of amino acids into proteins; thus, insulin is an anabolic hormone, contrasting the catabolic activity of glucagon (Röder et al., 2016). The external signals, such as nutrient intake or stress, induce an endocrine secretion
via humoral, neural or hormonal signaling pathways. The stimulus-secretion
coupling process translates these signals into hormonal release. This process is known as the stimulus-dependent exocytosis of a particular substance, such as glucose-stimulated β-cell insulin release (Ashcroft et al., 1994; Komatsu et al.,
11 2013). Glucose uptake is facilitated by the glucose transporter GLUT2 (SLC2A2) located on the surface of the β-cells, and once inside the cell, it undergoes glycolysis, thereby generating adenosine triphosphate (ATP), resulting in an increased ATP/ADP ratio. This altered ratio then leads to the closure of ATP-sensitive K+-channels (KATP-channels).
Insulin is stored in large dense-core vesicles that are recruited to the proximity of the plasma membrane following a stimulation (Röder et al., 2016). The key molecules that mediate the fusion of the insulin-containing large dense-core vesicles are the synaptosomal-associated protein of 25 kDa (SNAP-25), syntaxin-1 and synaptobrevin 2 (or vesicleassociated membrane protein VAMP2), all of which belong to the superfamily of the soluble N-ethylmaleimide-sensitive factor attachment protein (SNAP) receptor proteins (SNAREs). Together with the Sec1/Munc18-like (SM) proteins they form the so-called SNARE complex (Thurmond, 2007). In addition synaptotagmins, are highly expressed in neurons and endocrine cells, and were shown to participate in Ca2+-dependent exocytosis processes.
The insulin secretion from β-cells includes a great variety of modulators that trigger, potentiate or inhibit glucose-stimulated insulin secretion, primarily through G-protein-coupled receptors (GPCRs). The most traditional external factor that initiates insulin secretion is glucose. In addition to its trigger function, glucose also induces pathways that amplify insulin secretion through metabolism-cAMP (cyclic adenosine monophosphate) coupling or the incretin hormones glucagon-like peptide (GLP)-1 and glucose dependent insulinotropic peptide (GIP) (Henquin, 2014).
1.2. Influence of organs on pancreatic secretions a. The brain
The brain via the brain–islet axis is one of the interacting organs with insulin secretion. In fact, the pancreas is highly innervated with both, parasympathetic and sympathetic nerve fibers from the autonomic nervous system (Röder et al., 2016). At the same time, insulin receptors are widely distributed within the brain, including the hypothalamus, cerebral cortex, cerebellum and hippocampal formation in humans, as well as the olfactory and limbic areas, hypothalamus
-12 particularly the periventricular nucleus and the arcuate nucleus- hippocampus and the choroid plexus in rat brains. The hypothalamic brain derived neurotrophic factor (Figure 2) via efferent nerves modulates the glucagon secretion, whereas the melanocortin system directly reduces basal insulin levels by stimulating sympathetic nerve fibers via α-adrenoceptors (Röder et al., 2016).
Figure 2. The interplay of the pancreas with the brain, liver, gut as well as adipose and muscle tissue. BNDF, brain-derived neurotrophic factor; CCK, cholecystokinin; GIP, glucose-dependent insulinotropic peptide; GLP-1, glucagon-like peptide 1; GRP, gastrin-releasing peptide; IL-6, Interleukin 6; MCH, melanin concentrating hormone; NPY, neuropeptide Y; PACAP, pituitary adenylate cyclase-activating polypeptide; POMC, pro-opiomelanocortin; VIP, vasoactive intestinal peptide (Röder et al., 2016).
The various neuropeptides exert their effects through various pathways, including the extracellular signal-regulated kinase (ERK)/Akt pathway, and modulation of Ca2+-influx (melanin concentrating hormone) (Pissios et al., 2007), cAMP and, to a lesser extent, PI3K signaling (VIP and PACAP) (Borboni et al., 1999; Straub and Sharp, 1996), muscarinic/β-adrenoceptors signaling, PI3K/PKC signaling and Ca2+-mobilization from intracellular stores (gastrin-releasing peptide) (Gregersen and Ahrén, 1996; Pettersson and Ahrén, 1987). Likewise, insulin release is stimulated by the so-called cephalic phase insulin response (Bellisle et al., 1985); the neural mechanism underlying this phase was
13 found to include cholinergic and non-cholinergic processes as well as the dorsal vagal complex located in the medulla oblongata (Röder et al., 2016).
b. The liver
The liver has a key role in glucose homeostasis by storing (glycogenesis) or releasing (glycogenolysis/gluconeogenesis) glucose upon interaction with insulin and glucagon, respectively (Figure 2). The binding of glucagon to its hepatic GPCR evokes a signaling cascade, eventually resulting in the activation of PKA, which in turn stimulates two processes; one promotes glycogenolysis/gluconeogenesis and the other inhibits glycolysis/glycogenesis (Hers, 1990; Jiang and Zhang, 2003). Glycogenolysis is a multistep process that includes the PKA-mediated phosphorylation of phosphorylase kinase, cleavage of glucose-1-phosphate (G-1-P) from glycogen by activated glycogen phosphorylase a and the conversion of G-1-P into G-6-P, eventually resulting in phosphate and free glucose (Röder et al., 2016). Furthermore, insulin inactivates glycogen phosphorylase and glycogen synthase kinase (GSK)-3 through the PI3K pathway, which in turn activates glycogen synthase (Akpan et al., 1974; Miller and Larner, 1973; Stalmans et al., 1974).
The liver also excert its effect on insulin by repressing the expression of the phosphoenolpyruvate carboxykinase by disrupting the association of cAMP response element-binding protein and RNA polymerase (Duong et al., 2002); whereas the suppression of G-6-P requires PKBα/Akt and forkhead transcription factor (FOXO1) (Nakae et al., 2001; Schmoll et al., 2000), whose expression was shown to be diminished by the inhibition of GSK-3 (Lochhead et al., 2001). Moreover, hepatocyte-derived factors conversely influence the pancreas and/or insulin secretion. For instance, the loss of HNF1α results in an almost abolished insulin secretion, likely due to a decreased response to intracellular calcium (Röder et al., 2016).
c. The gut
Insulin secretion is potentited by gut released hormones GLP-1 and GIP upon nutrient ingestion binding to their respective receptors on pancreatic β-cells (Figure 2). Furthermore, these hormones exert pancreatic effects, such as
GLP-1-14 stimulated insulin gene expression, incretin induced β-cell neogenesis, proliferation and survival, the prevention of β-cell apoptosis in general and in response to glucolipotoxicity. The GLP-1 extrapancreatic actions includes the suppression of endogenous glucose production/glycogenolysis, glucagon secretion, appetite, a delay in gastric emptying and improved β-cell insulin sensitivity and glucose disposal, whereas GIP positively affects lipid and bone metabolism (Eckel et al., 1979; Egan et al., 2002; Röder et al., 2016; Song et al., 2007). In addition to incretins, limostatin and Neuromedin U (NmU) or decretins are secreted during fasting to suppress insulin release. Furthermore, the gastrin and cholecystokinin (CCK) are gastrointestinal hormones that interact with the pancreas. Finally, the gut microbiota is an important factor related to metabolic disorders. Obesity and T2DM patients display alterations in the composition of their microbiota that may initiate and/or promote these disorders.
d. The adipocytes/myocytes
The insulin interaction with adipose and muscle tissues is broadly based on facilitating insulin-dependent glucose uptake through the GLUT4 (Khan and Pessin, 2002; Kohn et al., 1996; Zisman et al., 2000), and the modulation of insulin release through the adipose and muscle tissue secretions of adipokines and myokines (Figure 2). The most famous adipokine leptin (Kieffer et al., 2000), acts on its receptors in the hypothalamic arcuate nucleus to inhibit food intake and control whole body homeostasis (Elmquist et al., 1999). However, the stimulation of the pancreatic leptin receptor (Ob-R) mRNA (Kieffer et al., 1996) can cause reduction in insulin secretion (Röder et al., 2016). Furthermore, leptin was shown to suppress insulin gene expression (Seufert et al., 1999; Seufert et al., 1999), representing a negative feedback loop. Conversely, insulin enhances ob gene expression and leptin secretion. Likewise, insulin modulates the expression of adiponectin, another well-known adipokine, the abundance of its receptor in adipose and muscle tissue as well as its secretion (Röder et al., 2016; Sattar and Sattar, 2012; Tsuchida et al., 2004). Adiponectin is not only involved in glucose and fatty acid metabolism (Yamauchi et al., 2002) but it also prevent β-cell apoptosis and induces insulin gene expression and release; Other adipokines, such as apelin, chemerin, omentin, resistin and visfatin, were also
15 shown to directly interact with insulin, whereas retinol-binding protein 4, tumor necrosis factor-α and vaspin are indirectly linked to insulin (Clarke and Mohamed-Ali, 2005). In addition fibroblast growth factor-21 is a widely expressed protein with a broad mode of action, including the regulation of carbohydrate and fatty acid metabolism (Potthoff et al., 2009) and may be considered as a myokine due to its secretion from muscle cells (Ribas et al., 2014).
2. Defects in glucose homeostasis 2.1. Diagnosis of hyperglycaemia
Hyperglycemia or high blood glucose happens when the body has too little insulin or when the body cannot use it properly (Szablewski, 2011). There are two specific types of hyperglycemia. First the fasting hyperglycemia with blood glucose levels greater than 90 – 130 mg/dL (5 – 7.2 mmol/L) after fasting for at least 8 hours. The postprandial (after-meal hyperglycemia) is characterized by blood glucose higher than 180 mg/dL (10 mmol/L). Early signs and symptoms of hyperglycemia include the following: increased thirst, headaches, difficulty concentrating, blurred vision, frequent urination, fatigue, weight loss, blood sugar more than 180 mg/dL (10 mmol/L), and high levels of sugar in the urine. Prolonged hyperglycemia in diabetes may result in vaginal and skin infections, slow-healing cuts and sores, decreased vision, nerve damage causing painful cold or insensitive feet, stomach and intestinal problems. The pre-diabetes status can be manifested in two forms: impaired glucose tolerance (IGT) and impaired fasting glucose (IFG). IGT is a transition phase between normal glucose tolerance and diabetes. It is a combination of impaired secretion of insulin and reduced insulin sensitivity (insulin resistance) (DeFronzo and Abdul-Ghani, 2011). IGT is diagnosed when: 1) plasma glucose, two hours after consuming 75 g glucose, appears to be superior to 7.8 mmol/L (normal level) but remains inferior to 11.1 mmol/L. The level of plasma glucose is measured by means of an Oral Glucose Tolerance Test (OGTT) (Standards of Medical Care in Diabetes, 2014). The procedure typically involves testing glucose levels after an eight hour fasting period, and measuring it again two hours after drinking a glucose solution. Generally, if the test shows blood glucose levels in the 140 and 199
16 mg/dL range, two hours after the drink, this could signify IGT. IGT is often affiliated with several other similar related risk factors such as high blood pressure (hypertension), increased LDL-cholesterol (Low Density Lipoprotein), reduced HDL-cholesterol (High Density Lipoprotein). A person has IGF when fasting plasma glucose is 100 to 125 mg/dL. This level is higher than normal but less than the level indicating a diagnosis of diabetes. DM is characterized by recurrent or persistent hyperglycemia and is diagnosed by demonstrating any one of the following: fasting plasma glucose level ≥ 7.0 mmol/L (126 mg/dL), plasma glucose ≥ 11.1 mmol/L (200 mg/dL) two hours after a 75 g oral glucose load as in a glucose tolerance test, symptoms of hyperglycemia and casual plasma glucose ≥ 11.1 mmol/dL, glycated hemoglobin (HbA1C) ≥ 6.5% (Szablewski, 2011).
2.2. Physiopathology of hyperglycemia
The cellular and molecular mechanisms responsible for T2DM are highly linked to the regulation and homeostasis of glycaemia. A normoglycaemia is maintained by the balanced interplay between the action and secretion of the insulin, a key hormone for glucose regulation. A decrease in insulin action is accompanied by upregulation of insulin secretion (and vice versa) in normal pancreatic β-cell. A curvilinear relation exist between normal β-cell function and insulin sensitivity (Bergman, 1989; Stumvoll et al., 2005).
17 Some exceptions, occurs when β-cell function is inadequately low for a specific degree of insulin sensitivity. Thus, β-cell dysfunction is a critical component in the pathogenesis of T2DM. When insulin action decreases such in obesity, the β-cell increases their functionality and activity. However, at the same time, concentrations of blood glucose at fasting and 2h after glucose load will increase mildly (Stumvoll et al., 2003). These small increases can over time become damaging and leads to β-cell dysfunction mainly because of glucose toxicity. Thus, even with hypothetical unlimited β-cell reserve, insulin resistance paves the way for hyperglycaemia and T2DM (Figure 3).
3. Diabetes and Oxidative stress 3.1. Oxidative stress
Oxidative stress occurs in a cellular system when the production of free radical moieties exceeds the antioxidant capacity of that system. The oxidized or nitrosylated products of free radical attack have decreased biological activity, leading to loss of energy metabolism, cell signaling, transport, and other major functions. These altered products also are targeted for proteosome degradation, further decreasing cellular function. Accumulation of such injury ultimately leads a cell to die through necrotic or apoptotic mechanisms. Several free radical species are normally produced in the body to perform specific functions. Superoxide (O2_.), hydrogen peroxide (H2O2), and nitric oxide (NO) are three free ROS that are essential for normal physiology, but are also believed to accelerate the process of aging and to mediate cellular degeneration in disease states (Mahjoub and Roudsari, 2012).
3.2. Glucose induced toxicity and oxidative stress
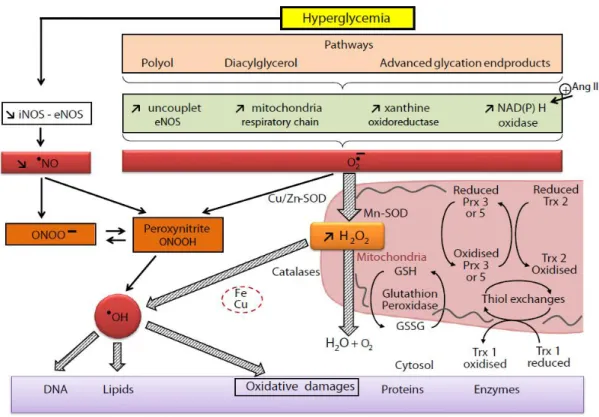
Hyperglycemia resulting from uncontrolled glucose regulation is widely recognized as the causal link between diabetes and diabetic complications (Rolo and Palmeira, 2006). Brief episodes of hyperglycemia cause tissue damage by mechanisms involving repeated acute changes in cellular metabolism (Figure 4). However, exposure to high glucose also causes cumulative changes in long-lived macromolecules, which persist despite restoration of euglycemia. A large amount of data emphasize four key metabolic pathways as being major
18
contributors to hyperglycemia induced cell damage (Robertson, 2004): (1) increased polyol pathway flux; (2) increased advanced glycation end product (AGE) formation; (3) activation of protein kinase C (PKC) isoforms; and (4) increased hexosamine pathway flux. ROS produced during normal metabolic processes are removed rapidly with the help of various endogenous detoxifying enzymes.
Figure 4. Hyperglycemia, induced oxidative stress. (Rochette et al., 2014)
While normal cellular ROS concentrations are necessary for proper functioning of cells, excessive, non-physiological concentrations of ROS result in oxidative stress. ROS such as O2•− and hydroxyl radicals (HO•), and hydrogen peroxide (H2O2), are highly reactive and can cause damage to biological macromolecules such as DNA, proteins, and lipids (Storz, 2011). Major sources of oxidative stress during diabetes include glucose autooxidation, overproduction of ROS by mitochondria, non-enzymatic glycation, and the polyol pathway (Giacco, 2011; Wohaieb and Godin, 1987). Under normal physiological conditions, ROS are eliminated efficiently by the body antioxidant defense mechanisms. Hyperglycemia and diabetes are thought to cause impairment of the endogenous antioxidant defense system (Bonnefont-Rousselot et al., 2000). Consequently, the mitochondrial production of ROS exceeds the capacity of the normal
19 antioxidant defenses. Mitochondrial overproduction of ROS in hyperglycemia has been postulated to cause redox imbalance, oxidative insults, mitochondrial dysfunction, and cell death (Green et al., 2004; Suomalainen and Kaukonen, 2001). The overwhelmed free radicals can damage DNA integrity, membrane lipids and protein function by oxidation and lead to functional abnormalities, apoptosis or necrosis (Wold et al., 2005). Chronic exposure of the β-cell to supraphysiologic concentrations of glucose causes defective insulin gene expression accompanied by marked decreases in insulin content and abnormal insulin secretion (Robertson et al., 1992). The clinical state of diabetes mellitus is often accompanied by elevated blood levels of cholesterol, triglyceride, and free fatty acids. This gave rise to the consideration that deteriorating beta cell function in diabetic patients might be caused by chronic exposure to high concentrations of lipids, a concept termed the lipotoxicity hypothesis (LeRoith et al., 2004). Prolonged exposure of pancreatic beta cells to fatty acids has been reported to inhibit insulin gene expression (Briaud et al., 2001).
20
UNIT 2. FOOD INTAKE AND GLUCOSE REGULATION
21 1. Food antioxidants and glucose regulation
The primary goal in the treatment of diabetes is to maintain blood glucose levels within the normal physiological range. It is recognized that diet plays a major role in the incidence and management of diabetes. Medicinal plants, herbs, and their derivatives have been used for centuries in the traditional treatment of diabetes, and are being studied intensively for their mechanism of action and active principles (Aggarwal, 2011; Marles and Farnsworth, 1995). The consumption of various foods has also been shown in animal and human studies to have positive impacts on blood glucose regulation (Table 1)(Lacroix and Li-Chan, 2014; Robertson, 2012).
Table 1. Antidiabetic mechanisms of Food ingredients (Lacroix and Li-Chan, 2014)
Antidiabetic
properties Mechanisms Examples of food plants and compounds
Reduction of glucose uptake
Inhibition of
carbohydrate-hydrolyzing enzymes Flavonoids and tannins (many fruits and vegetables), sorghum, sardine, and egg white proteins Inhibition of intestinal glucose
transport Polyphenols (tea), strawberry, and apple extracts Inhibition of hepatic glucose output Chlorogenic acid (green coffee)
Delay of intestinal glucose absorption resulting from the formation of viscous gels that slow down the diffusion of glucose in the lumen
Soluble fibers (oat, barley), guar gum, high-viscosity hydroxypropylmethylcellulose
Enhancement of
incretin effect Stimulation of GLP-1 secretion triggered by short-chain fatty acids released during bacterial
fermentation of fiber or by proteins and peptides
Fermentable fibers, starches, and sugar alcohol (e.g. cornstarch, lactitol, Viscofiber®), corn and milk proteins,
and meat hydrolysate
Inhibition of DPP-IV Peptides or hydrolysates of milk, rice bran, egg, corn, and fish proteins
Enhancement of insulin sensitivity
Inhibition of PTP1B Cinnamon, oxidized catechins (tea)
Activation of PPAR- γ Unsaturated fatty acids, isoflavones genistein, and daidzein (soy), many herbs and spices (e.g. thyme, oregano, rosemary), 20(S)-protopanaxatriol, psi-baptigenin, hesperidin, apigenin, chrysin, and kaempferol Enhancement of
insulin action Mimicking of insulin functions Cinnamon, allspice, bay leaves, nutmeg, clove, epigallocatechin gallate, epicatechin gallate, tannins, theaflavins (tea), conglutin- γ(lupin), tannic acid 1,2,3,4,6-penta-O-galloyl-D-glucopyranose
Enhancement of
insulin secretion Stimulation of insulin secretion Anthocyanins (e.g. cyanidin-3-glucoside), delphinidin-3-glucoside and pelargonidin-3 galactoside (fruits), S-allyl cysteine sulfoxide and diallyl trisulfide (garlic), and bitter gourd
Alleviation of
oxidative stress Prevention of oxidant formation or radical chain propagation via the activation of Nrf2 mediated ARE
Flavonols (fruits, vegetables, tea, wine), lipoic acid, vitamin E,β-carotene, and vitamin C
Over the last few decades, numerous studies have been carried out to try and identify the antidiabetic constituents in these food products and their mechanisms of action. Dietary components, including soluble fiber, phenolic
22 compounds and peptides, have been shown to display various antihyperglycemic properties, such as inhibitory activity against digestive enzymes, insulin secretagogue effects, and activation of receptors involved in glucose metabolism. Although not as potent as synthetic drugs, the antidiabetic constituents from food products could have the advantage of causing less undesirable side effects and thus, could represent an interesting complementary approach in the management of diabetes (Lacroix and Li-Chan, 2014).
2. Oat compounds and glucose regulation
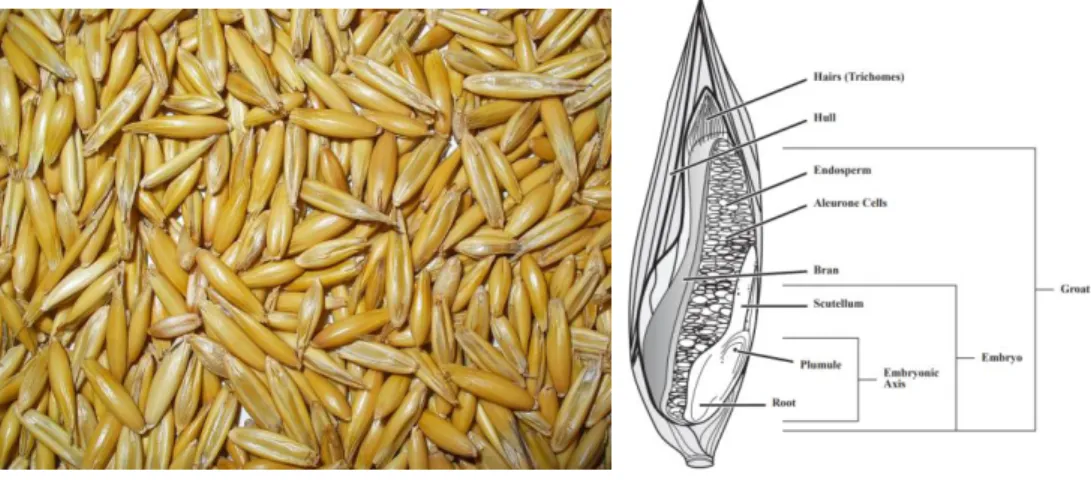
The complex and intimate association between increased oxidative stress and increased inflammation in related disorders such as T2DM makes it difficult to establish the temporal sequence of the relationship (Rochette et al., 2014). In diabetic patients, the increase in oxidative stress is associated with a decline in cellular antioxidant defenses. Several attempts have been made to reduce oxidative stress-dependent cellular changes in patients with diabetes by antioxidant supplementation.
Figure 5. Oat seeds morphology.
Oat (Figure 5) plays a role in modulating the metabolic effects observed after fiber-rich meals. As a soluble fiber with viscous characteristics, Oat β-glucans modifies properties of chyme in the upper part of the gastrointestinal tract affecting gastric emptying, gut motility, and nutrient absorption, which are reflected in lower postprandial glycemic and insulin responses (Daou and Zhang, 2012).