Peatlands and the carbon cycle: from local processes to global implications ? a synthesis
42
0
0
Texte intégral
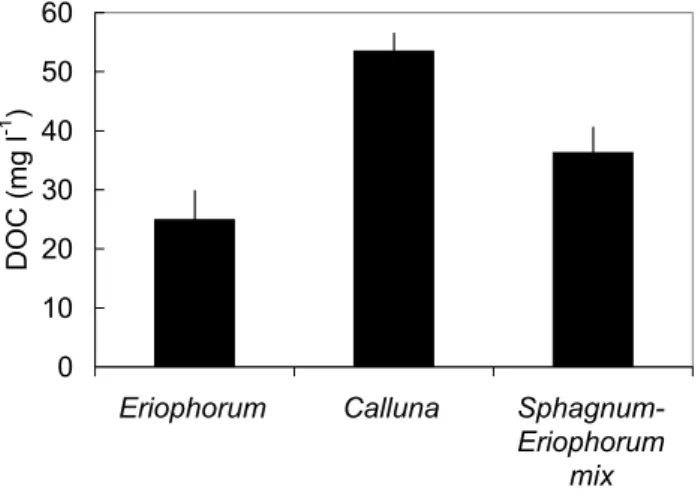
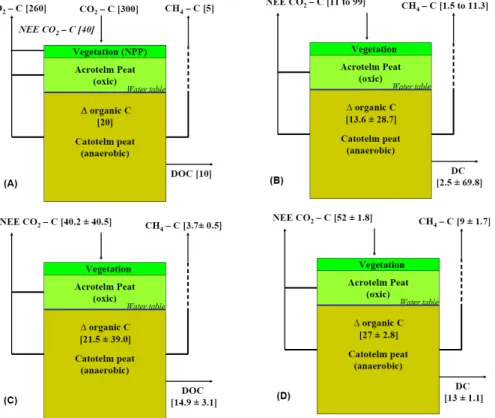
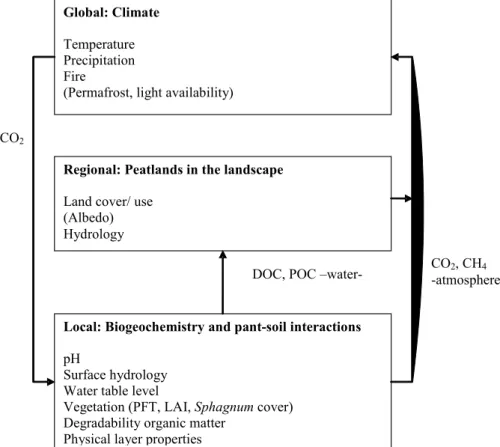
Figure


+2
Documents relatifs