Dynamics and statics of actin assemblies
Texte intégral
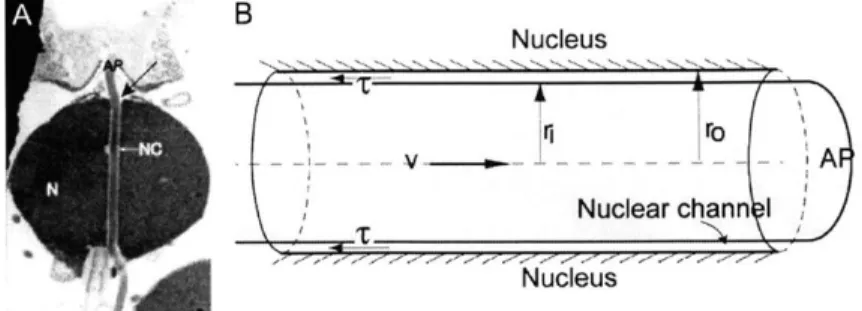
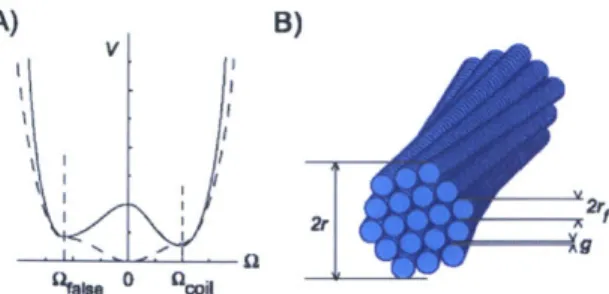
Figure
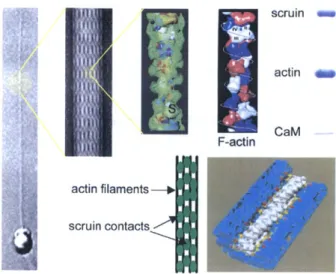


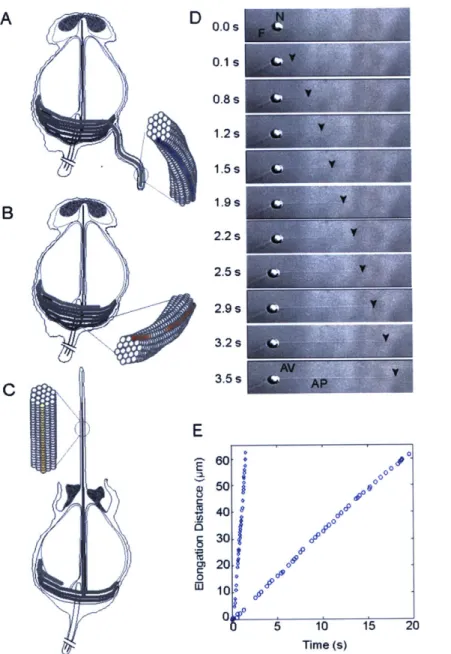
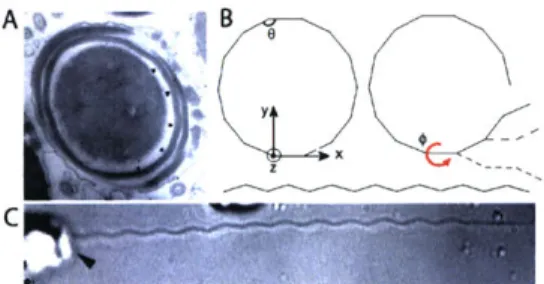
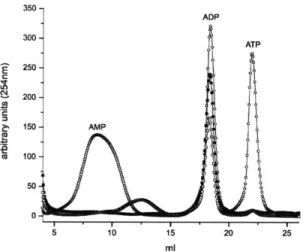


Documents relatifs
In the present paper, based on recent data describing PEBP1 relationships with various molecular targets, we reinforce the idea that the likely principal functionality
The study focuses on two aspects: the first seeks to know how frequently these principles are used and the second stresses the importance of the availability of
Lorsqu'une modification se produit dans leur teneur en humidité, de nombreux matériaux de construction changent de dimensions et ces changements peuvent être plus
Number of incoming (outgoing) links of all the nodes attached to a sector, divided by the total number of links—banks (green), broker-dealers (purple), insurance companies (blue)
À l’aide d’une méthode automatisée de reconnaissance d’entités nommées et d’une analyse bibliométrique détaillée dans la section suivante, cette étude s’attache à
Taken together, these accumulated data motivate our revised definition of ‘good health’ to include a sense of purpose for a more meaningful life. It is simply amazing that
In recent years several authors have described interesting dynamical behavior of SIR epidemiological models in which the population can be portioned into two age structured
In carcinoma cells, consistent with the role of L-plastin in controling actin dynamics, the PKC activator PMA induced the translocation of L-plastin to de novo actin
