Dynamique du pin blanc (Pinus strobus Linnaeus) dans
les tourbières ombrotrophes du sud du Québec
Mémoire
Sarah-Kim Lavoie
Maîtrise en sciences géographiques
Maître en sciences géographiques (M. Sc. Géogr.)
Québec, Canada
ii
Dynamique du pin blanc (Pinus strobus Linnaeus) dans
les tourbières ombrotrophes du sud du Québec
Mémoire
Sarah-Kim Lavoie
Sous la direction de :
Martin Lavoie directeur de recherche
Martin Simard codirecteur de recherche
iii
RÉSUMÉ
Plusieurs tourbières du Québec sont aujourd’hui caractérisées par un processus de boisement qui est souvent une conséquence des activités anthropiques in ou ex situ ayant affecté leurs conditions hydrologiques. Bien que le pin blanc (Pinus strobus) soit une espèce arborescente que l’on rencontre plutôt rarement dans les tourbières ombrotrophes, les individus qui y poussent semblent afficher dans plusieurs cas une bonne croissance, autant en hauteur qu’en diamètre. Les objectifs de la présente étude étaient (1) de déterminer l’époque d’établissement de l’espèce dans quelques tourbières de la région écologique de la Plaine du Saint-Laurent et les facteurs ayant favorisé cet établissement, (2) de caractériser leur croissance radiale et (3) de déterminer les facteurs qui ont régi la croissance. Cinq tourbières ont été étudiées. Des analyses dendrochronologiques ont été réalisées pour des individus sur tourbe et sur sol minéral à proximité afin de comparer la croissance entre les deux types de milieu. Des fonctions de réponse et de corrélation ont été effectuées sur les séries dendrochronologiques afin de déterminer l’influence du climat sur la croissance. Enfin, des monolithes de tourbe de surface furent l’objet d’une analyse macrofossile afin de trouver des restes de pin blanc. La présence du pin blanc en milieu tourbeux semble être un phénomène relativement récent, la colonisation ayant débuté au plus tôt un peu avant la moitié du XIXe siècle.
Cette installation récente concorde avec la présence de macrorestes de l’espèce dans les seuls premiers centimètres de la tourbe en surface. Bien qu’elle soit majoritairement inférieure à celle des individus sur sol minéral, la croissance radiale des individus sur tourbe s’avère très bonne. Le climat ne semble pas avoir influencé la croissance des individus et ce, autant sur tourbe que sur sol minéral ce qui suggère que d’autres facteurs régissent la croissance. Les précipitations moins abondantes durant certaines périodes du XXe siècle et le feu sont, selon les sites, des facteurs
iv
TABLE DES MATIÈRES
RÉSUMÉ ... iii
TABLE DES MATIÈRES ... iv
LISTE DES TABLEAUX ... vi
LISTE DES FIGURES ... vii
REMERCIEMENTS ... viii INTRODUCTION ... 1 Objectifs et hypothèses ... 6 CHAPITRE PREMIER ... 9 1 Sites à l’étude ... 9 DEUXIÈME CHAPITRE ... 15 2 Méthodologie ... 15 2.1 Travaux de terrain ... 15 2.2 Travaux de laboratoire ... 15 2.2.1 Analyses dendrochronologiques ... 15 2.2.2 Données météorologiques ... 16 2.2.3 Analyses macrofossiles... 17 TROISIÈME CHAPITRE ... 18 3 Résultats ... 18 3.1 Analyses dendrochronologiques ... 18
3.1.1 Dates d’établissement du pin blanc ... 18
3.1.2 Croissance radiale ... 20
3.1.3 Données météorologiques ... 23
3.1.4 Fonctions de réponse et de corrélation ... 25
3.2 Analyses macrofossiles ... 27
QUATRIÈME CHAPITRE ... 29
4 Discussion ... 29
4.1 Époques d’établissement ... 29
4.2 Les facteurs pouvant expliquer l’établissement et la croissance du pin blanc sur les tourbières ... 30
4.2.1 Le feu ... 30
4.2.2 Les facteurs climatiques... 31
v CONCLUSION ... 35 RÉFÉRENCES ... 37 ANNEXE 1 ... 43 ANNEXE 2 ... 48 ANNEXE 3 ... 53 ANNEXE 4 ... 72 ANNEXE 5 ... 91
vi
LISTE DES TABLEAUX
Tableau 1 : Statistiques des séries dendrochronologiques ... 20 Tableau 2 : Largeur moyenne des cernes par site en fonction du substrat ... 21
vii
LISTE DES FIGURES
Figure 1 : Aire de répartition du pin blanc en Amérique du Nord ... 1
Figure 2 : Présence du pin blanc dans des tourbières du Québec méridional : ... 4
Figure 3 : Système racinaire du pin blanc en surface dans deux tourbières du Québec méridional : .. 5
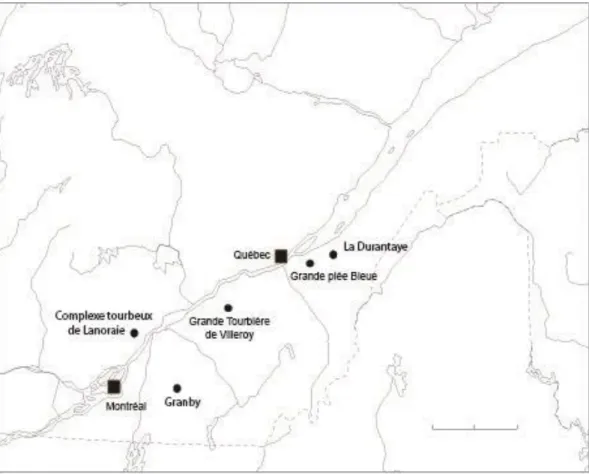
Figure 4 : Localisation des sites d’étude ... 9
Figure 5 : Vue de la Grande tourbière de Villeroy ... 10
Figure 6 : Vue de la tourbière la Grande-plée-Bleue ... 11
Figure 7 : Vue de la tourbière de La Durantaye ... 12
Figure 8 : Vue du complexe tourbeux de Lanoraie ... 13
Figure 9 : Vue de la tourbière de Granby ... 14
Figure 10 : Dates d’établissement du pin blanc sur tourbe et sur sol minéral pour chaque site ... 19
Figure 11 : Chronologies maîtresses des individus sur tourbe et sur sol minéral pour chacun des sites. ... 22
Figure 12 : Déviations climatiques aux stations météorologiques de Jean-Lesage (Québec) et de Joliette.. ... 24
Figure 13 : Fonctions de réponse et de corrélation. ... 26
Figure 14 : Diagrammes macrofossiles des restes de pin blanc et des charbons de bois des dix monolithes de tourbe. ... 28
viii
REMERCIEMENTS
Je tiens à remercier en premier lieu mon directeur Martin Lavoie ainsi que mon codirecteur Martin Simard pour l’opportunité qu’ils m’ont donnée d’avoir travaillé sur cet intéressant projet de recherche au cours des deux dernières années. Merci d’avoir cru en moi et en ce projet, d’y avoir mis temps et énergie. Je ne vous remercierai jamais assez pour ce que vous avez fait pour moi.
Des sincères remerciements à Ann Delwaide, professionnelle de recherche au Centre d’études nordiques et responsable du laboratoire de dendrochronologie, qui a mis à ma disposition son temps et ses connaissances. J’apprécie tout l’aide qu’elle a su donner, elle fut précieuse.
Je tiens également à remercier Myosotis Bourgon Desroches, professionnelle de recherche au Centre d’études nordiques et responsable du laboratoire de paléoécologie terrestre, qui m’a fort aidé tout au long de mon projet de recherche.
Merci également à M. Serge Payette, professeur au Département de biologie et à M. Michel Caillier, professeur retraité au Département des sols et de génie agroalimentaire, qui ont toujours démontré un certain intérêt pour ma recherche. Merci de votre temps et surtout de vos conseils.
Merci à M. Pierre Grondin et Mme Véronique Poirier du Ministère des forêts, de la faune et des parcs du Québec pour avoir fourni les cartes des sites potentiels. Merci aussi à M. Grondin pour avoir accepté d’être membre de mon comité d’évaluation.
Merci à M. Jean-Maurice Champagne qui nous a permis d’échantillonner les pins sur son terrain à Lanoraie et à M. Guy Chabot à qui nous a donné accès à son érablière à Granby.
Un merci spécial à Héloïse Barbel, amie et voisine de bureau qui a toujours su être présente pour moi dans les meilleurs comme dans les mauvais moments. Merci à toi, pour tout ce que tu as fait depuis que nous nous connaissons. Je te suis éternellement reconnaissante.
Un dernier merci à ma famille, mes proches, mes ami(e)s et collègues qui ont su m’épauler tout au long de cette aventure. Merci de votre inconditionnel support.
1
INTRODUCTION
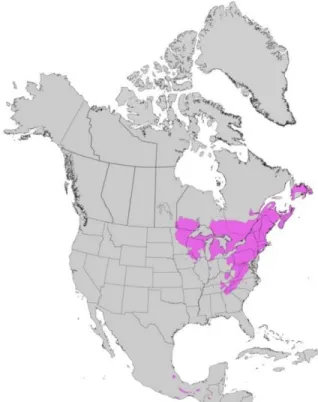
Le pin blanc (Pinus strobus Linnaeus) est une espèce arborescente présente uniquement en Amérique du Nord. Au Canada, son aire de répartition s’étend de Terre-Neuve à l’est jusqu’à la pointe sud-est du Manitoba à l’ouest (Figure 1). Aux États-Unis, il est présent dans 14 états ; son aire de répartition continue couvre les états du Minnesota, du Wisconsin et du Michigan à l’ouest et ceux de la Nouvelle-Angleterre à l’est. Elle se prolonge au sud jusqu’en Georgie dans l’axe de la chaîne de montagnes des Appalaches (Wendel et Clay Smith, 1990). Au Québec, la limite nord de la répartition continue du pin blanc se trouve au sein du domaine bioclimatique de la sapinière à bouleau jaune (Saucier et al. 2009). Elle s’étend, d’ouest en est, de l’Abitibi (secteur du lac Abitibi ; latitude 48°N) à la région de Charlevoix. Elle se prolonge ensuite vers l’est de part et d’autre de l’estuaire et du golfe du St-Laurent : du côté nord le long d’une bande littorale de quelques dizaines de kilomètres de largeur jusqu’à Baie-Comeau, incluant les secteurs de la rivière Saguenay et du lac St-Jean ; du côté sud au sein de la péninsule gaspésienne. L’espèce est aussi présente sur l’île d’Anticosti (Figure 1).
Figure 1 : Aire de répartition du pin blanc en Amérique du Nord. Source : www.arboquebecium.com
2
Il est fréquent de trouver le pin blanc sur des sols de qualité moyenne à pauvre, comme du sable ou du loam sableux bien drainé (Wendel et Clay Smith, 1990; Vlasiu et al., 2001). Il s’agit d’une espèce semi-tolérante à l’ombre (25 à 55 % d’ensoleillement) qui peut atteindre une hauteur moyenne de 30 m et un diamètre moyen de 100 cm. Sa durée de vie atteint aisément 200 ans et au maximum 400 à 450 ans. La production de graines s’amorce généralement vers l’âge de 20-30 ans ; elle culmine entre 50 et 150 ans pour ensuite diminuer. Puisque le pin blanc ne possède pas de cônes sérotineux, le feu n’est pas un facteur essentiel à la libération des graines, mais il peut néanmoins s’avérer utile pour sa régénération. En effet, cette perturbation permet, entre autres, d’ouvrir le couvert végétal, de réduire temporairement la compétition et de préparer des lits de germination adéquats (Wendel et Clay Smith, 1990; Vlasiu et al., 2001).
Les diagrammes polliniques de l’est de l’Amérique du Nord montrent que le pin blanc fut particulièrement abondant à la charnière entre l’Holocène inférieur et l’Holocène moyen, entre ~8000 et ~5000 ans avant nos jours, alors que les conditions climatiques étaient relativement chaudes (période hypsithermique; p. ex. : Liu, 1990; Richard, 1994; Anderson, 1995; Muller et Richard, 2001). Dans certaines régions comme en Abitibi et dans le nord-est de l’Ontario, sa limite septentrionale se situait alors à environ 100 km plus au nord qu’aujourd’hui (Terasmae et Anderson, 1970; Liu, 1990). La représentation pollinique du pin blanc a ensuite diminué dans les régions tempérées au profit de celle d’autres espèces comme la pruche du Canada (Tsuga canadensis) et le hêtre à grandes feuilles (Fagus grandifolia). Le pin blanc était plus abondant que de nos jours au sein des forêts précoloniales du sud du Québec. Il fut beaucoup exploité et vendu au XIXe siècle, particulièrement
dans les années 1820. La coupe sélective des tiges de grande dimension, notamment pour la production de mâts de navires, a contribué à réduire considérablement son abondance, comme en témoigne les actes notariés de l’époque (Simard et Bouchard, 1996; Brisson et Bouchard, 2003).
Au Québec, de 9 à 12 % du territoire est recouvert par les milieux humides, notamment des tourbières (Payette et Rochefort, 2001). Celles-ci sont des systèmes non équilibrés au sein desquels le taux de production de la matière organique excède le taux de décomposition. Un niveau élevé de la nappe phréatique est nécessaire au maintien des conditions anaérobiques et ainsi permettre l’accumulation de matière organique (la tourbe). Les tourbières ombrotrophes (bogs), qui sont les plus abondantes dans le sud du Québec, hébergent une flore vasculaire relativement peu diversifiée
3
en raison, entre autres, des conditions acides et des apports en éléments minéraux provenant des seules précipitations (estivales et hivernales) (Payette et Rochefort, 2001). Chez les arbres, l’épinette noire (Picea mariana) et le mélèze laricin (Larix laricina) sont les deux espèces les plus fréquentes dans les bogs, en compagnie parfois d’autres espèces comme le thuya occidental (Thuja
occidentalis) et le sapin baumier (Abies balsamea) par exemple. L’épinette noire et le mélèze laricin
peuvent former des peuplements de densité variable, de sorte que plusieurs tourbières ombrotrophes présentent une structure semi-forestière. Il arrive toutefois que l’on rencontre aussi d’autres espèces d’arbres, notamment chez les tourbières dont les conditions hydrologiques furent altérées par les activités humaines in situ (p. ex. : exploitation de la tourbe ou horticulture) ou ex situ (p. ex. : agriculture, urbanisation). La création de canaux de drainage pour ces activités a pour principale conséquence l’assèchement des conditions du sol. Les nouvelles conditions édaphiques profitent à certaines espèces de plantes vasculaires qui pourront alors s’installer. Chez les arbres, il s’agit, entre autres, du bouleau gris (Betula populifolia), de l’érable rouge (Acer rubrum), du pin gris (Pinus banksiana) et du peuplier faux-tremble (Populus tremuloides) (Lavoie et St-Louis, 1999; Pellerin et Lavoie, 2003a; 2003b; Talbot et al., 2010; Lavoie et Pellerin, 2015; Pasquet et al., 2015; Beauregard, 2017). D’autres espèces arborescentes feuillues sont aussi rencontrées à l’occasion comme le bouleau jaune (Betula alleghaniensis), le frêne noir (Fraxinus nigra) ou encore l’orme liège (Ulmus thomasii). Le boisement des tourbières semble d’ailleurs un processus en accélération qui est observé à plusieurs endroits dans l’est de l’Amérique du Nord (Pellerin et Lavoie, 2003a; 2003b; Berg et al., 2009; Talbot et al., 2010; Ireland et Booth, 2012; Pasquet et al., 2015; Pellerin et al., 2016a; 2016b), plus particulièrement depuis le début du XXe siècle. Le même phénomène s’observe
aussi en Europe; selon le cas, les espèces impliquées sont le pin sylvestre (Pinus sylvestris; Linderholm et Leine, 2004; Cedro et Lamentowicz, 2008; 2011; Lamentowicz et al., 2008; Cubizolle et al., 2009; Smiljanić et al., 2014; Hökkä et al., 2012), le pin à crochets (Pinus uncinata; Reille, 1991; Freléchoux et al., 2000), l’épicéa commun (Picea abies; Klempířová et al., 2013) ou encore le bouleau pubescent (Betula pubescens; Cubizolle et al., 2009).
Si la présence d’un couvert forestier dense sur les tourbières ombrotrophes est souvent la conséquence d’activités humaines, le déploiement massif d’arbres peut aussi être d’origine naturelle. En plusieurs endroits en Europe, des souches et des troncs de pin sylvestre ensevelis dans la tourbe ont été trouvés, datant de 4000 à 8500 ans selon les endroits (Eckstein et al., 2011). Il en est de
4
même pour des chênes dont des troncs datant de différentes époques de l’Holocène ont également été trouvés. Les périodes d’afforestation des tourbières pendant l’Holocène en Europe ont été associées à des périodes climatiques plus chaudes et plus sèches, comme celle de l’hypsithermique (Edvardsson et al., 2014).
Le pin blanc se rencontre aussi à l’occasion dans les tourbières du sud du Québec (Figure 2). Comme pour beaucoup d’autres espèces arborescentes, la tourbe ne représente pas à priori un substrat idéal pour la croissance en raison de son humidité élevée, de la présence de la nappe phréatique près de la surface et de sa pauvreté en éléments nutritifs. L’apport en eau, la texture et l’espace disponible pour les racines sont, au niveau du sol, les facteurs qui affectent le plus le potentiel de croissance du pin blanc (Wendel et Clay Simth, 1990; Vlasiu et al., 2001).
Figure 2 : Présence du pin blanc dans des tourbières du Québec méridional : Grande tourbière de Villeroy (gauche) et tourbière de La Durantaye (droite).
Source : Sarah-Kim Lavoie, 2015
Aux États-Unis, quelques travaux se sont penchés sur la présence du pin blanc en milieu tourbeux. Au New Hampshire (Lyon, 1949), un peuplement s’est établi dans une tourbière au début des années 1900. Il a été montré que les cernes de croissance les plus larges correspondaient à des années de faibles précipitations pendant la saison de croissance, probablement parce que le niveau de la nappe phréatique était plus bas et que la tourbe était plus sèche en surface, ce qui aurait favorisé la
5
croissance du pin. De plus, le système racinaire s’est développé de manière horizontale au-dessus de la nappe phréatique, soit dans les 20 premiers centimètres du dépôt. Un tel déploiement horizontal du système racinaire du pin blanc dans la tourbe a aussi été observé au Québec (Figure
3) et pour le pin à crochets en France (Bégeot et Richard, 1996). En Virginie occidentale, les mesures
du taux de croissance en hauteur du pin blanc dans un environnement tourbeux ont montré que les individus étaient plus petits que ceux croissant sur sol sableux et leurs cernes de croissance en moyenne environ deux fois plus étroits (Edens et Ash, 1969).
Figure 3 : Système racinaire du pin blanc en surface dans deux tourbières du Québec méridional : Grande tourbière de Villeroy (gauche) et Lanoraie (droite).
Source : Sarah-Kim Lavoie, 2016
Dans le cadre d’une étude récente d’une tourbière en Pennsylvanie comportant un peuplement de pin blanc, Ireland et Booth (2012) ont examiné l’évolution temporelle des concentrations en éléments nutritifs (azote et phosphore) dans la tourbe à la suite du déboisement des terres en périphérie de la tourbière. Les apports éoliens en éléments minéraux résultaient de l’utilisation d’engrais chimiques utilisés en agriculture. Selon les analyses dendrochronologiques, le pin blanc s’était établi au début des années 1900. Les analyses macrofossiles de la tourbe ont confirmé cette installation récente, les macrorestes de l’espèce n’ayant été trouvés qu’à la surface du dépôt de tourbe. La colonisation de la tourbière par le pin pourrait avoir été favorisée par un
6
enrichissement en phosphore concomitante à une baisse du niveau la nappe phréatique (déduite par l’analyse des thécamoebiens) en raison des activités agricoles environnantes (Ireland et Booth, 2012). Au Canada, la végétation de la tourbière Oil Well Bog dans le sud de l’Ontario a été étudiée par Bunting et al. (1998). Le pin blanc est présent sur des sols podzoliques en périphérie de la tourbière ainsi que dans la tourbière même. Un diagramme pollinique réalisé à partir d’une carotte sédimentaire prélevée dans la tourbière a montré une nette augmentation de la représentation pollinique du pin à partir des années 1920. Cette augmentation traduit fort probablement l’époque de son établissement sur la tourbière. Les changements survenus au sein du cortège floristique de la tourbière ont été associés à une altération des conditions hydrologiques locales. Tout comme pour la tourbière en Pennsylvanie, les terres avoisinantes ont été asséchées pour l’agriculture, ce qui a eu aussi pour effet d’abaisser la nappe phréatique et donc fort probablement de favoriser l’installation du pin.
En Pologne, Cedro et Lamentowicz (2011) ont comparé la croissance radiale d’une population de pin sylvestre établie dans une tourbière ombrotrophe avec celle d’individus de la même espèce en marge de la tourbière croissant sur substrat mésique. Les résultats furent ensuite comparés à des données climatiques à l’aide d’une fonction de réponse. Les individus sur substrat mésique montraient une croissance qui répondait aux effets du climat (température moyenne mensuelle et précipitations mensuelles). En revanche, celle des individus dans la tourbière était plutôt liée aux changements de la position de la nappe phréatique induite par les activités humaines. Le drainage à des fins d’extraction de la tourbe, qui s’opère dans cette région depuis 1880, est vraisemblablement la première cause ayant mené à l’assèchement de la tourbe et ultimement à l’établissement du pin sylvestre. Malgré les conditions locales plus sèches, les individus sur tourbe affichaient une croissance radiale nettement inférieure à celle des individus sur substrat mésique.
Objectifs et hypothèses
Au Québec, aucune étude n’a encore porté spécifiquement sur le pin blanc dans les tourbières ombrotrophes. Il s’agit d’une espèce considérée « facultative des milieux terrestres », c’est-à-dire qu’elle n’est présente qu’occasionnellement dans les milieux humides (USDA 2017). Bien qu’elle ne soit pas une espèce commune des tourbières, elle a néanmoins été répertoriée dans plus de 45
7
placettes d’inventaire forestier sur sol organique du ministère des Forêts, de la Faune et des Parcs du Québec (MFFP) pour la seule région écologique de la Plaine du Saint-Laurent (région écologique 2b) (Grondin et Poirier, 2015). Cette région écologique, qui couvre 17 000 km2, est liée de très près
à la région physiographique des basses-terres du Saint-Laurent (Gosselin, 2005). Les individus observés lors de la campagne d’échantillonnage semblent afficher dans plusieurs cas une bonne croissance, autant en hauteur qu’en diamètre. L’analyse macrofossile de carottes sédimentaires de quelques-unes de ces tourbières (Grande-plée-Bleue, Grande tourbière de Villeroy) suggère toutefois que sa présence serait un phénomène très récent puisqu’aucune pièce macrofossile de pin (p. ex. : feuilles, graines) n’a été trouvée dans les assemblages macrofossiles et ce, depuis l’origine des tourbières (Lavoie et al., 2012; Lavoie et Colpron-Tremblay, 2013). Dans le même ordre d’idées, l’examen de 86 diagrammes macrofossiles de tourbières, répartis sur l’ensemble de l’aire de répartition de l’espèce en Amérique du Nord, milite aussi en faveur du fait que le pin n’a jamais été présent de façon importante sur les tourbières dans le passé car aucun de ces diagrammes ne montre des restes de l’espèce.
Dans ce contexte, les objectifs du présent projet étaient :
(1) de déterminer l’époque ou les époques de l’installation du pin blanc dans cinq tourbières ombrotrophes de la région physiographique des basses-terres du St-Laurent et de déterminer quels furent le ou les facteurs ayant favorisé cette installation;
(2) de caractériser la croissance radiale du pin blanc en milieu tourbeux et de la comparer avec celle d’individus croissant en périphérie sur sol mésique;
(3) et de déterminer quels furent les principaux facteurs qui ont régi la croissance radiale du pin blanc dans les tourbières.
Les hypothèses qui ont été testées sont que :
(1) la présence du pin blanc dans les tourbières ombrotrophes est un phénomène relativement récent (XXe siècle) en raison de conditions hydriques locales devenues plus sèches au sein des
8
(2) les individus sur tourbe affichent une croissance radiale plus faible que ceux sur substrat mésique.
Afin d’atteindre les objectifs et de tester les hypothèses, j’ai étudié cinq tourbières réparties dans la région physiographique des basses terres du Saint-Laurent. Pour chacun des sites, des analyses dendrochronologiques ont été réalisées pour des individus croissant directement dans la tourbière, de même que pour des individus croissant sur sol minéral en périphérie dans le but de comparer leur croissance radiale. Afin de voir s’il était possible que le pin blanc se soit installé sur la tourbe depuis plus longtemps que la période temporelle livrée par la dendrochronologie, deux monolithes de tourbe de surface ont été échantillonnés dans chacune des tourbières pour examiner leur contenu en macrorestes de pin blanc. Un macroreste est une pièce végétale de taille macroscopique visible à l’œil nu conservée dans un sédiment ou un sol. En raison de son poids et de sa taille, elle n’est généralement pas dispersée sur de grandes distances et témoigne donc de la présence locale de l’espèce l’ayant produit (Birks, 2013).
9
CHAPITRE PREMIER
1
Sites à l’étude
Des sites comportant une association de pin blanc sur tourbe (T) et sur sol minéral (M) ont été visités. Cinq tourbières ombrotrophes ont été sélectionnées (Figure 4). Une des difficultés rencontrées dans le choix des tourbières était que certaines étaient situées au sein d’un paysage hautement agricole, de sorte qu’il n’y avait pas nécessairement d’individus de pin blanc croissant sur sol minéral présents en milieu forestier en périphérie des tourbières.
10
La Grande Tourbière de Villeroy (VRY-T et VRY-M), d’une superficie de 1590 ha (46°22’46’’N; 71°49’17’’O), est la plus vaste tourbière ombrotrophe de la région physiographique des basses-terres du Saint-Laurent encore peu perturbée au sud du fleuve Saint-Laurent. Elle a commencé à se former il y a 10 200 ans A.A. (avant aujourd’hui) suite au retrait de la Mer de Champlain (Lavoie et Colpron-Tremblay, 2013). La tourbe, qui atteint une épaisseur maximale de 450 cm, repose selon les endroits sur du till ou des dépôts éoliens (Avard et al., 2013; Larocque et al., 2013). La tourbière présente une structure végétale semi-forestière où l’épinette noire et le mélèze laricin sont les deux espèces arborescentes les plus abondantes. La tourbière est bordée, au nord-est, par des dunes paraboliques aujourd’hui stabilisées par la végétation (Filion, 1987). Celles-ci hébergent divers peuplements, entre autres une pessière mixte comportant l’épinette rouge (Picea rubens), l’épinette noire et le pin blanc, de même qu’une érablière à érable rouge (Payette et al., 2016). De très vieilles souches de pin blanc datant de plusieurs siècles sont aussi présentes sur les dunes (Payette et al., 2013) (Figure 5). Cette tourbière a fait l’objet d’une étude paléoécologique par Lavoie et Colpron-Tremblay (2013). L’analyse macrofossile de trois carottes sédimentaires n’avait pas révélé la présence passée du pin blanc sur la tourbière.
Figure 5 : Vue de la Grande tourbière de Villeroy : secteur de la tourbière (gauche) et secteur en périphérie sur les dunes avec les souches de pin blanc (droite).
11
La tourbière de la Grande-plée-Bleue (GPB-T) (46°46’46’’N; 71°03’19’’O), d’une superficie de 1500 ha, a commencé à se former il y a environ 9460 ans A.A. à la suite du retrait de la Mer de Goldthwait (Lavoie et al., 2012). Elle repose, selon les endroits, sur des sédiments littoraux, prélittoraux ou d’exondation mis en place en eau peu profonde. La tourbière présente une structure semi-forestière dominée par l’épinette noire et le mélèze laricin. Elle comporte aussi un réseau de 650 mares (Figure 6). Elle est ceinturée de forêts mixtes comprenant entre autres le sapin baumier et l’épinette noire. Aucun pin blanc n’est toutefois présent au sein des forêts périphériques. La tourbière est caractérisée par la présence de deux canaux de drainage aujourd’hui bloqués à des fins de restauration (Hugron et al., 2013). Des études paléoécologiques (macrorestes végétaux, thécamoebiens) ont été réalisées pour deux carottes sédimentaires dans le secteur des mares (Lavoie et al., 2012; Garneau et Lamarre, 2013). Aucun macrofossile de pin blanc n’avait alors été trouvé (Lavoie et al., 2012).
Figure 6 : Vue de la tourbière la Grande-plée-Bleue : secteur de la tourbière dominée par une structure arbustive d’éricacées (gauche) et une des mares situées au sein d’une structure semi-forestière (droite).
12
La tourbière de La Durantaye (DUR-T et DUR-M) (46°49’14’’N ; 70°50’15’’O) est constituée d’une section ombrotrophe ouverte dominée par les sphaignes, les éricacées et l’épinette noire ainsi que d’une section forestière dense caractérisée par le peuplier faux-tremble, l’aulne (Alnus sp.) et le pin blanc (Figure 7). Un canal de drainage est présent à plus de 100 m de la portion où se trouve le peuplement de pin blanc. Le site d’échantillonnage sur sol minéral est situé à 7 km de la tourbière. À cet endroit, une litière d’environ 30 cm d’épaisseur repose sur du sable fin. Les espèces dominantes sont le mélèze laricin, l’épinette noire et le pin blanc. La régénération est, quant à elle, dominée par l’érable rouge (Figure 7).
Figure 7 : Vue de la tourbière de La Durantaye : secteur forestier de la tourbière (gauche) et secteur forestier sur sol minéral situé à 7 km de la tourbière (droite).
Source : Sarah-Kim Lavoie, 2016
Le complexe tourbeux de Lanoraie (LAN-T et LAN-M) (45°57’41’’N; 73°19’06’’O) couvre une superficie de 76 km² (Pellerin et al., 2016a). L’accumulation organique aurait débuté vers 7400 ans A.A. sur des dépôts argileux de la Mer de Champlain suite à la réorganisation du système fluviatile au sein d’anciens chenaux bordés par des terrasses deltaïques sableuses (Comtois, 1982; Rosa et Larocque, 2008). L’épaisseur maximale de la tourbe est de 840 cm (Rosa et al., 2009). Les zones ombrotrophes sont dominées par l’épinette noire, le mélèze laricin, les éricacées et les
13
sphaignes tandis que les zones minérotrophes sont surtout caractérisées par l’érable rouge, l’aulne rugueux (Alnus rugosa) et le houx verticillé (Ilex verticillata) (Pellerin et al., 2016a). Les sols sableux qui bordent le complexe tourbeux, dont des dunes stabilisées, sont surtout dominés par le peuplier faux-tremble, l’érable rouge et le pin blanc (Figure 8). L’étude macrofossile de deux carottes de tourbe de surface d’un mètre d’épaisseur d’un secteur du complexe n’a révélé aucun macroreste de pin blanc (Pellerin et al., 2016a). Le contenu en azote et phosphore de même qu’une analyse des thécamoebiens des deux mêmes carottes a également été étudié (Pellerin et al., 2016a). Le site d’échantillonnage du pin blanc sur sol minéral se situe à 10 km du site d’échantillonnage sur tourbe.
Figure 8 : Vue du complexe tourbeux de Lanoraie : secteur forestier de la tourbière (gauche) et secteur forestier sur sol minéral situé à 10 km (droite).
Source : Sarah-Kim Lavoie, 2016
La tourbière de Granby (GRY-T et GRY-M) (45°21’57’’N ; 72°43’37’’O) est une tourbière ombrotrophe constituée d’une section ouverte dominée par les sphaignes, les éricacées, l’épinette noire et le pin blanc et d’une vaste section densément boisée par l’épinette noire, le mélèze laricin et le bouleau gris. La tourbière est bordée au sud par une érablière à érable à sucre (Acer
14
Figure 9 : Vue de la tourbière de Granby : secteur semi-forestier de la tourbière (gauche) et le secteur forestier sur sol minéral (droite).
15
DEUXIÈME CHAPITRE
2
Méthodologie
2.1
Travaux de terrain
Pour chacun des sites, 40 individus de pin blanc ont été échantillonnés à des fins d’analyse dendrochronologique : 20 en milieu tourbeux et 20 sur substrat minéral en périphérie de la tourbière. À la tourbière Grande-plée-Bleue, tous les individus de grand diamètre présents sur tourbe ont été échantillonnés, ce qui représentait seulement 11 individus. L’échantillonnage des individus sur sol minéral, à la même tourbière, fut impossible puisqu’aucun pin blanc n’était présent autour de la tourbière. Pour chaque individu, deux carottes ont été prélevées à l’aide d’une sonde de Pressler le plus près possible du collet (après dégagement de la tourbe dans le cas de ceux en milieu tourbeux). Deux monolithes de tourbe de surface (~20 × 20 × 20 cm), numérotés M1 et M2, ont été récoltés à l’aide d’une pelle à des fins d’analyse macrofossile afin de voir jusqu’à quelle profondeur dans le dépôt organique des restes de pin blanc étaient présents. Les monolithes ont été emballés dans une pellicule de papier cellophane puis dans du papier d’aluminium pour leur transport. Afin d’éviter leur dessèchement et le développement de moisissures, ils ont été entreposés au réfrigérateur à une température de 4ºC jusqu’au moment des analyses.
2.2
Travaux de laboratoire
2.2.1 Analyses dendrochronologiques
En laboratoire, les carottes ont été séchées à l’air libre, puis collées sur des supports en bois et sablées (grain 400). Les cernes de croissance ont été interdatés sous une loupe binoculaire
16
(grossissement de 40×) et leur largeur mesurée (précision de 2 μm) à l’aide d’une table Velmex (Velmex incorporated, Bloomfield, New York, USA). L’interdatation fut vérifiée à l’aide du logiciel COFECHA (Grissino-Mayer 2001). Des séries maîtresses furent construites pour chaque site et chaque substrat (tourbe et sol minéral) en standardisant avec une spline cubique de 9 ans et en sélectionnant les séries à l’aide de la fonction strip de la bibliothèque dplR (Bunn, 2010). Puisque qu’il n’y avait pas de pin sur sol minéral en périphérie de la tourbière Grande-plée-Bleue, la comparaison des individus sur tourbe de ce site fut effectuée avec ceux sur sol minéral du site La Durantaye.
2.2.2 Données météorologiques
Afin d’examiner l’influence possible du climat sur l’établissement du pin blanc dans les tourbières et la croissance des individus sur les deux types de substrat, les données des stations météorologiques d’Environnement et Changements Climatiques Canada (www.ec.gc.ca) les plus complètes et proches possibles des tourbières ont été utilisées. Pour les tourbières Villeroy, Grande-plée-Bleue et La Durantaye, les données de la station météorologique de l’aéroport Jean-Lesage (Québec) ont été retenues pour les analyses car il s’agissait des données les plus complètes s’échelonnant depuis 1873. Pour les sites de Lanoraie et Granby, la station météorologique de Joliette fut utilisée car elle comporte des données complètes depuis 1919. Des stations météorologiques étaient situées plus proches de certains sites, mais les périodes temporelles couvertes par ces dernières étaient relativement courtes et/ou les données incomplètes. Des fonctions de réponse et de corrélation entre les largeurs moyennes indicées des cernes de croissance et les données climatiques ont été établies (bibliothèque bootRes; Zang et Biondi, 2013). Les variables climatiques retenues pour les analyses étaient la température moyenne et les précipitations totales (1) mensuelles (mois de septembre de l’année précédente au mois d’août de l’année en cours), (2) saisonnières (juin-juillet-août, mars-avril-mai, décembre [année précédente]-janvier-février et septembre-octobre-décembre (année précédente) et (3) annuelles.
17
2.2.3 Analyses macrofossiles
Les monolithes de tourbe ont été coupés en tranches contiguës de 1 cm d’épaisseur et une couche de matière organique en surface a été éliminée pour éviter une contamination qui aurait pu se produire lors de l’échantillonnage. Les analyses macrofossiles ont été réalisées à tous les centimètres d’intervalle selon le protocole de Bhiry et Filion (2001). Un sous-échantillon d’un volume de 20 cm³ a été prélevé au centre de chaque tranche. Les sous-échantillons ont été bouillis pendant quelques minutes dans une solution d’eau et d’hydroxyde de potassium (KOH 5 %) pour défloculer la matière organique. Ils ont ensuite été tamisés à travers un tamis de maille 425 μm. Tous les macrorestes de pin blanc ont été dénombrés et identifiés. Les charbons de bois ont également été dénombrés afin de voir si l’installation du pin sur les tourbières fut liée à un événement de feu. Les résultats sont présentés sous la forme de dénombrement par unité de volume (nombre de pièces par 20 cm3).
18
TROISIÈME CHAPITRE
3
Résultats
3.1
Analyses dendrochronologiques
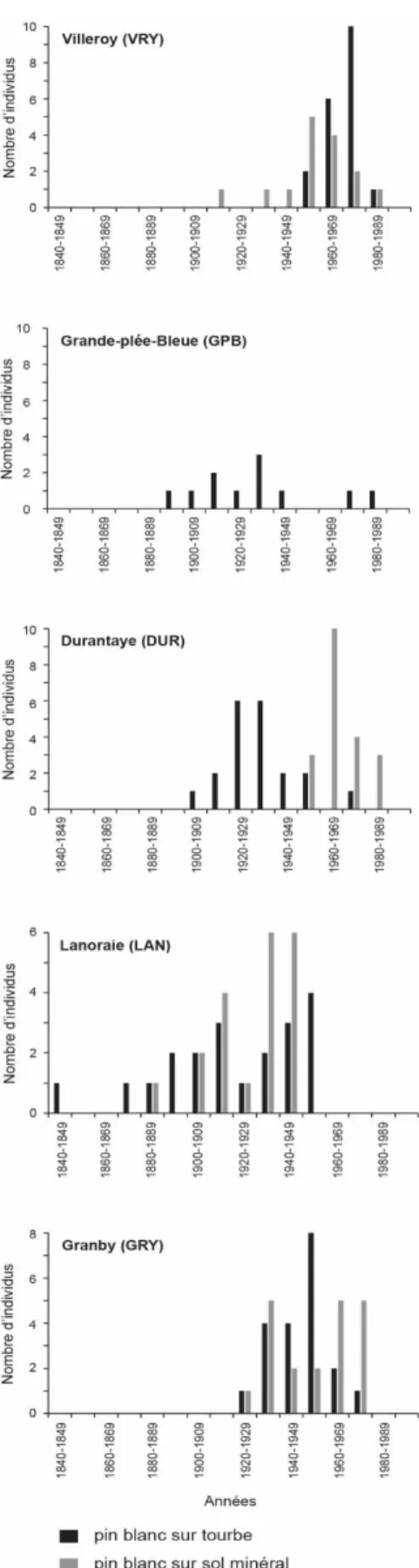
3.1.1 Dates d’établissement du pin blanc
Pour chacun des sites, la majorité des individus échantillonnés, autant ceux sur tourbe que ceux sur sol minéral, se sont établis au 20e siècle (Figure 10), à l’exception de quelques individus sur tourbe
au site Lanoraie qui datent de la seconde moitié du 19e siècle, le plus ancien s’étant établi durant la
décennie 1840. Les patrons d’établissement sont relativement différents entre les sites, autant pour ce qui est de la date du premier individu que pour la longueur de la période d’établissement. En effet, certains sites n’ont été que très récemment colonisés par le pin blanc (e.g., après 1950 à la tourbière de Villeroy et sur sol minéral à La Durantaye) alors que d’autres ont été exclusivement colonisés avant le milieu du 20e siècle (Lanoraie). De plus, l’établissement s’est fait dans certains cas sur de
relativement courtes périodes (40 ans à la tourbière de Villeroy et sur sol minéral à La Durantaye) et dans d’autres cas sur plus d’un siècle (tourbières de la Grande-plée-Bleue et de Lanoraie). Dans chacun des sites, le patron d’établissement est généralement similaire entre les deux types de milieu, à l’exception du site La Durantaye, où la colonisation de la tourbière était pratiquement terminée lorsque les premiers arbres se sont établis sur le sol minéral. Très peu de petits pins blancs (hauteur < 2 m) ont été observés en tourbière et aucun n’a été carotté pour établir leur âge. Ils étaient rares dans les sites Villeroy, Grande-plée-Bleue, La Durantaye et Granby et complètement absents à Lanoraie, bien que des cônes ouverts étaient présents au sol.
19
Figure 10 : Dates d’établissement du pin blanc sur tourbe et sur sol minéral pour chaque site. Aucun individu sur sol minéral n’a été échantillonné au site Grande-plée-Bleue.
20
3.1.2 Croissance radiale
La chronologie la plus longue sur tourbe était celle de Lanoraie (171 ans), tandis que la plus courte était celle de Villeroy (53 ans) (Tableau 1). Malgré le fait qu’elles étaient bien interdatées, les séries étaient faiblement intercorrélées (0,11 à 0,32) et leur sensitivité moyenne, qui quantifie la variabilité interannuelle de la largeur des cernes (Speer 2010), était relativement faible (< 0,2) pour tous les sites. L’EPS (Expressed Population Signal) est une mesure de la variabilité commune dans une chronologie et exprime à quel point les arbres échantillonnés représentent le peuplement en entier (Speer 2010). L’EPS était supérieur à 0,85 dans tous les sites sauf pour celui de la Grande-plée-Bleue sur tourbe où il était plus faible (0,80) et où le nombre d’arbres échantillonnés (n = 11) était d’ailleurs inférieur à celui des autres sites (n = 20).
Tableau 1 : Statistiques des séries dendrochronologiques.
Site Code Nombre
d’arbres Période couverte Nombre de cernes Inter-corrélation Sensitivité moyenne EPS*
Villeroy - tourbe VRY-T 20 1962-2014 53 0,16 0,138 0,91
Villeroy - minéral VRY-M 20 1924-2015 92 0,28 0,159 0,87
Grande-plée-Bleue - tourbe GPB-T 11 1907-2015 109 0,32 0,128 0,80
La Durantaye - tourbe DUR-T 20 1907-2016 110 0,21 0,087 0,90
La Durantaye - minéral DUR-M 20 1957-2016 60 0,22 0,094 0,93
Lanoraie - tourbe LAN-T 20 1846-2016 171 0,12 0,162 0,91
Lanoraie - minéral LAN-M 20 1888-2016 130 0,31 0,136 0,96
Granby - tourbe GRY-T 20 1931-2016 86 0,32 0,124 0,95
Granby - minéral GRY-M 20 1934-2016 83 0,11 0,116 0,85
*Expressed population signal
De manière générale, la croissance radiale des individus poussant sur sol minéral est plus élevée que celle de ceux sur tourbe (Tableau 2; Figure 11). C’est au site Lanoraie que la différence est la plus faible entre les deux types de substrat (0,58 mm), alors que c’est au site La Durantaye qu’elle est la plus élevée (2,21 mm). Parmi les sites sur tourbe, la croissance est la plus forte à Villeroy et la plus faible à Grande-plée-Bleue. Chez les individus sur sol minéral, la largeur moyenne des cernes est sensiblement la même à Villeroy et à La Durantaye alors qu’elle est plus faible à Lanoraie.
21
Tableau 2 : Largeur moyenne des cernes par site en fonction du substrat pour les années 1962-2014.
Site Largeur moyenne des cernes des individus
sur tourbe (mm)
Largeur moyenne des cernes des individus sur sol minéral
(mm) Villeroy 3,411 ± 2,976 4,558 ± 1,949 Grande-plée-Bleue 1,991 ± 1,149 n.d. La Durantaye 2,343 ± 1,507 4,554 ± 1,646 Lanoraie 2,451 ± 1,482 3,072 ± 1,755 Granby 2,2275 ± 1,109 3,668 ± 2,976
Les courbes de croissance des individus sur tourbe montrent une absence d’augmentation soudaine ou graduelle de croissance qui témoignerait d’une amélioration rapide des conditions de croissance. En effet, elles montrent que la croissance est constante comme à la Grande-plée-Bleue et Granby tandis qu’elle diminue à La Durantaye et qu’elle augmente à Villeroy (Figure 11). Il est également difficile d’établir une correspondance entre les courbes de croissance et les patrons d’établissement. Cependant, les courbes de croissance démontrent que la croissance radiale est beaucoup moins prononcée chez les individus en tourbière à l’exception du site de Lanoraie et que l’établissement de ces mêmes individus s’est produit durant le XXe siècle.
22
Figure 11 : Chronologies maîtresses des individus sur tourbe (trait plein) et sur sol minéral (tireté) pour chacun des sites. Pour le site Grande-plée-Bleue, la courbe des individus sur sol minéral du site La Durantaye a été utilisée en raison de l’absence de pin blanc sur sol minéral en périphérie de la tourbière.
23
3.1.3 Données météorologiques
Les graphiques de déviations climatiques (Figure 12) des stations météorologiques de Jean-Lesage (associée aux sites Villeroy, Grande-plée-Bleue et La Durantaye) et de Joliette (associée aux sites Lanoraie et Granby) montrent que les précipitations totales annuelles étaient, de façon générale, inférieures à la moyenne jusqu’au début de la décennie 1970. Elles ont ensuite augmenté, bien que l’on observe quelques années aux anomalies négatives, notamment à la station Jean-Lesage durant la deuxième moitié des années 1990 et au début des années 2000.
En ce qui concerne les températures estivales (mai-juin-juillet-août) à la station Jean-Lesage, elles furent supérieures à la normale pendant trois décennies (1930 à 1960) pour ensuite diminuer jusqu’au début des années 1980. À Joliette, elles ont été généralement supérieures à la moyenne pendant deux périodes : de 1935 à 1955 et depuis le début des années 1990.
24
.
Figure 12 : Déviations climatiques par rapport à la moyenne pour les précipitations totales annuelles et la température estivale moyenne (mai, juin, juillet et août) aux stations météorologiques de Jean-Lesage (Québec) et de Joliette. Les tiretés représentent l’écart-type et la ligne noire la moyenne mobile sur trois ans.
25
3.1.4 Fonctions de réponse et de corrélation
De manière générale, les fonctions de réponse et de corrélation montrent peu de relations significatives entre la croissance radiale du pin blanc et les paramètres climatiques et ce, autant sur tourbe que sur sol minéral (Figure 13). De plus, les quelques relations significatives obtenues n’étaient constantes ni à l’intérieur d’un même site (tourbière versus sol minéral), ni entre les sites pour un même type de milieu (sol minéral des cinq sites ou tourbe des cinq sites). Seule une corrélation positive entre les précipitations du mois de mai et la croissance du pin sur sol minéral aux sites Lanoraie et Granby fut obtenue.
26
Figure 13 : Fonctions de réponse et de corrélation. Les relations significatives sont indiquées par la couleur bleue (relations positives) et la couleur rouge (relations négatives).
Ptot : précipitations totales annuelles, Tmoy : température moyenne annuelle, JJA : juin-juillet-août, MAM : mars-avril-mai, DJF : décembre-janvier-février, SON : septembre-octobre-novembre. L’exposant « -1 » apposé à un mois réfère au mois de l’année précédente.
Voir le tableau 1 pour les codes d’identification des sites.
27
3.2
Analyses macrofossiles
L’analyse macrofossile des monolithes de tourbe montre que les macrorestes de pin blanc sont surtout présents et abondants dans les premiers centimètres à la surface du dépôt de tourbe (0-5 cm) et ce, pour chacun des sites (Figure 14). Il s’agit essentiellement d’aiguilles (complètes ou fragmentées) et de quelques graines. Aux sites de Lanoraie et de Granby, des pièces ont été trouvées dans l’entièreté de l’épaisseur des monolithes, surtout dans le monolithe Granby-M1, où elles sont les plus abondantes. Dans ces cas particuliers, on observe néanmoins aussi une nette augmentation de l’abondance des pièces dans les premiers centimètres. Des charbons de bois ont aussi été trouvés en quantité variable. Leur nombre est toutefois peu important dans la plupart des cas et leur représentation est inégale entre les monolithes d’un même site et ce, même si les lieux de prélèvement sont relativement proches l’un de l’autre au sein d’une même tourbière. Pour certains sites (Grande-plée-Bleue et Lanoraie), des charbons ont été trouvés dans un seul des deux monolithes. Les concentrations en charbons les plus importantes ont été trouvées au site de Villeroy (dans les premiers centimètres supérieurs des deux monolithes) et à la base des monolithes Grande-plée-Bleue – M2 et Lanoraie – M2.
28
Figure 14 : Diagrammes macrofossiles des restes de pin blanc et des charbons de bois des dix monolithes de tourbe. Les « * » représentent les monolithes pour lesquels aucun charbon de bois ne fut observé.
29
QUATRIÈME CHAPITRE
4
Discussion
4.1
Époques d’établissement
Les analyses dendrochronologiques des individus de pin blanc ont montré, conformément à notre première hypothèse, que leur établissement sur les tourbières à l’étude fut un phénomène relativement récent, les individus s’étant établis entre 1910 et 1980, outre un site (Lanoraie) où l’établissement s’est amorcé plus tôt, principalement au courant de la décennie 1840. Cet établissement concentré surtout au XXe siècle est appuyé par les analyses macrofossiles des monolithes de tourbe de tous les sites
qui montrent une présence ou une augmentation accrue de pièces macrofossiles dans les premiers centimètres en surface. Bien que ni la base des monolithes (qui se situe à 15-20 cm de profondeur selon le cas) ni l’amorce de l’augmentation marquée de l’abondance des macrorestes n’aient été datées (14C, 210Pb), les chronologies au 210Pb de l’accumulation verticale de la tourbe en surface
obtenues pour quelques tourbières ombrotrophes du sud du Québec (Lavoie et al., 2013; Pellerin et al., 2016a), dont celle de la Grande-plée-Bleue (Garneau et Lamarre, 2013), montrent que les 25-30 cm d’accumulation de tourbe de sphaignes peu décomposées à la surface représentent moins d’une centaine d’années. De ce fait, outre les deux monolithes du site de Lanoraie où la matière organique présentait un état de décomposition avancé, les monolithes des quatre autres sites sont constitués de sphaignes fraîches vivantes peu compactes correspondant à l’acrotelme des tourbières ; il est donc fort probable que l’âge de la base des monolithes soit inférieur à 100 ans. Dans le cas particulier du site de Lanoraie, qui était le plus forestier des sites à l’étude, le caractère très décomposé de la matière organique pourrait peut-être indiquer que la base des monolithes est plus ancienne.
30
L’examen qualitatif de 77 diagrammes macrofossiles, provenant de tourbières situées au sein de l’aire de répartition du pin blanc suggère également que l’espèce n’est présente de façon importante que depuis récemment (résultats non présentés), comme par exemple à la tourbière Titus bog en Pennsylvanie (Ireland et Booth, 2012). Bien que l’absence de macrorestes ne signifie pas nécessairement que l’espèce ne fut jamais présente localement sur les tourbières pendant l’Holocène, dont celles de la Grande-plée-Bleue et de Villeroy pour lesquelles deux et trois carottes sédimentaires furent respectivement l’objet d’analyses macrofossiles (Lavoie et al., 2012; Lavoie et al., 2013), les indices portent à croire que si ce fut le cas, il s’agissait sans doute d’une colonisation marginale et moins importante qu’aujourd’hui.
Un tel établissement récent du pin blanc sur tourbe s’inscrit dans le processus de boisement de plusieurs tourbières ombrotrophes du sud du Québec et de l’est de l’Ontario, processus qui a pris beaucoup d’ampleur depuis la seconde moitié du XXe siècle, autant par des espèces conifériennes
(épinette noire, mélèze, pin gris) que par des espèces feuillues (bouleau gris, érable rouge) (Jean et Bouchard, 1987; Pellerin et Lavoie, 2003a; 2003b; Talbot et al., 2010; Pasquet et al., 2015; Pellerin et al., 2016a; 2016b; Beauregard, 2017). Des études paléoécologiques en Europe ont aussi montré que la présence du pin sylvestre sur les tourbières est un phénomène récent et où aucun macrofossile ne fut trouvé dans la tourbe (Gałka et al., 2015; Lamentowicz et al., 2015a; 2015b). Des études dendrochronologiques ont également été réalisées sur des populations de pin sylvestre établies en tourbières : en Estonie, l’arbre le plus âgé avait 203 ans (Smiljanić et al., 2014) alors qu’en Lettonie le pin le plus âgé avait 98 ans (Dauškane et Elferts, 2011) et 171 ans en Suède (Linderholm, 2001).
4.2
Les facteurs pouvant expliquer l’établissement et la croissance du pin
blanc sur les tourbières
4.2.1 Le feu
Contrairement au pin gris, le pin blanc ne possède pas de cônes sérotineux ; le feu n’est donc pas obligatoire à la libération de ses graines et à sa régénération. Cette perturbation permet cependant d’ouvrir le couvert forestier, de réduire temporairement la compétition, de préparer des lits de
31
germination adéquats tout en augmentant la disponibilité en éléments nutritifs (Vlasiu et al., 2001; Pellerin et Lavoie, 2003a). Au XXe siècle, des feux d’abattis non contrôlés ont sévi en 1921, 1932 et
1933 dans le secteur du village de Val-Alain à quelques kilomètres à l’est de Villeroy. Un autre feu serait survenu en 1949 mais, selon Payette et al. (2013), il n’aurait pas atteint la tourbière. Ces auteurs ont soulevé la possibilité que le feu de 1921 soit à l’origine de la composition forestière actuelle des dunes bordant la tourbière. D’après les résultats des présentes analyses dendrochronologiques, les individus de pin blanc sur ces dunes ainsi que ceux sur tourbe se sont établis à la suite des feux de 1921 et 1949 respectivement. Les deux monolithes de tourbe, prélevés relativement proches de la bordure forestière, comportent d’ailleurs d’importantes quantités de charbons de bois en surface, ces charbons ayant sans doute été produits lors du feu de 1949. D’autres charbons de bois plus profonds dans le monolithe M-2 de la même tourbière pourraient pour leur part être associés à un des feux antérieurs à celui de 1949. Pour le site de Villeroy, il est donc possible que les individus sur tourbe puissent être issus du feu de 1949, tandis que ceux sur les dunes sont probablement issus du feu d’abattis de 1921. À la tourbière Grande-plée-Bleue, un versant de la tourbière a été affecté par le passage d’un feu d’origine anthropique au XIXe siècle qui est illustré par un mince horizon de charbons
de bois dans la tourbe (Payette, 2001). La grande abondance des charbons et le degré élevé de décomposition de la tourbe sous la couche carbonisée suggèrent qu’une couverture forestière relativement dense devait déjà être présente lors du passage du feu (Payette, 2001). Les charbons présents à la base du monolithe M-2 de cette tourbière sont peut-être associés à ce feu mais, à la lumière des données dendrochronologiques, il n’est pas possible d’établir un lien quelconque entre le feu et le pin blanc. Pour les autres sites, on ne dispose pas pour le moment d’informations concernant les feux (naturels ou anthropiques) ayant pu avoir eu lieu au XXe siècle. Bien que des charbons de
bois aient été dénombrés et qu’ils témoignent de feux ayant eu lieu à proximité et peut-être même sur les tourbières (entre autres LAN-M2 et GBY-M1), le signal n’est toutefois pas uniforme entre les monolithes d’un même site. Il est donc difficile de s’avancer sur le rôle du feu comme facteur responsable de l’établissement du pin blanc sur tourbe.
32
Les graphiques de déviations climatiques des stations météorologiques de l’aéroport Jean-Lesage (Québec) et de la ville de Joliette montrent que les précipitations annelles furent généralement inférieures à la normale pendant plusieurs décennies consécutives, soit entre 1910 et 1970. Aux deux endroits, la température estivale fut aussi plus élevée que la normale durant les décennies 1930, 1940 et 1950. Puisque les tourbières ombrotrophes sont alimentées en eau uniquement par les précipitations, il est possible que la nappe phréatique ait été généralement plus basse et les conditions de surface moins humides durant ces années en raison des moindres précipitations et d’une possible évapotranspiration plus élevée. De telles conditions auraient ainsi été en principe favorables à l’établissement du pin blanc sur tourbe. Le processus de boisement des tourbières entraîne également un effet d’assèchement en raison de l’absorption de l’eau par les racines.
Dans ce contexte, on observe qu’une grande partie des individus sur tourbe des sites de Villeroy, La Durantaye, Lanoraie et Granby se sont établis lors de périodes durant lesquelles les précipitations étaient inférieures à la normale, ce qui suggère que le climat ait pu contribuer à l’établissement du pin blanc. Au site Grande-plée-Bleue, le petit nombre d’individus échantillonnés ainsi que l’étalement des âges sur près de 100 ans ne permet pas de conclure à un rôle potentiel du climat, bien que l’analyse des thécamoebiens d’une carotte sédimentaire prélevée dans le secteur des mares (Garneau et Lamarre, 2013) montre un abaissement de la nappe phréatique au XXe siècle qui pourrait être soit de
nature climatique, soit le résultat de la création d’un canal de drainage (voir plus bas).
Le climat est certainement le facteur qui exerce la plus grande influence sur la croissance des végétaux. Dans les forêts tempérées et boréales, le facteur limitant est généralement la température puisque les précipitations sont relativement abondantes pendant la saison de croissance. Des températures plus froides ont généralement pour effet de réduire la croissance, quoique les hautes températures puissent également diminuer la croissance en augmentant l’évapotranspiration, créant ainsi un stress hydrique (Chhin et al., 2013). L’effet des précipitations sur la croissance est surtout visible lorsque les conditions de drainage sont extrêmes, c’est-à-dire dans les milieux xériques et dans ceux mal drainés tels les tourbières. Les précipitations ont alors un effet contraire sur la croissance : dans les sites xériques, la croissance est positivement reliée aux précipitations, alors que c’est l’inverse dans les sites très mal drainés (Drobyshev et al., 2010). Les présents résultats ont démontré que la croissance du pin blanc était effectivement plus faible en milieu tourbeux comparativement à celle sur
33
substrat minéral. Cependant, les fonctions de réponse et de corrélation n’ont détecté aucune influence des variations interannuelles de la température ou des précipitations sur la croissance. Ceci pourrait donc suggérer que climat n’a pas été un facteur limitant pour la croissance du pin blanc dans les sites échantillonnés.
4.2.3 Le drainage
Le drainage des tourbières est un autre facteur ayant pu contribuer, dans certains cas, à l’établissement du pin blanc. Le drainage, par l’intermédiaire de la création de canaux, a pour conséquence de diminuer la hauteur de la nappe phréatique et d’assécher la tourbe, pouvant du coup favoriser leur boisement (Pellerin et Lavoie, 2003; Talbot et al., 2010). La distance sur laquelle les canaux de drainage ont un impact sur la nappe phréatique et ultimement sur la composition floristique est difficile à estimer et varie d’un site à l’autre. Cet impact dépend de plusieurs facteurs propres à chacun des sites dont la topographie de surface de la tourbière, l’âge des canaux ou encore le nombre et la distance entre les canaux. Différentes études menées sur ce sujet suggèrent des impacts sur l’acrotelme se faisant sentir sur des distances variant de 30 à 200 m selon le cas (Jutras et al, 2007 ; Landry et Rochefort, 2012).
Entre 1950 et 1960, le gouvernement du Québec avait approuvé le drainage de grandes tourbières, dont celle de la Grande-plée-Bleue (Doyon, 2016). Des canaux ont alors été creusés, en plus d’approfondir et d’élargir ceux déjà existants afin d’augmenter le débit d’évacuation de l’eau. Cependant ces travaux se sont avérés inefficaces et n’ont pas contribué à l’assèchement de la tourbière. Par contre, le drainage a eu pour conséquence un léger boisement essentiellement le long des canaux sur quelques mètres de largeur seulement (Doyon, 2016). Comme cela fut mentionné plus haut, l’analyse des thécamoebiens d’une carotte sédimentaire prélevée dans le secteur des mares de cette tourbière non loin de l’extrémité du plus large canal, montre une baisse de la profondeur de la nappe phréatique au cours des dernières décennies (Garneau et Lamarre, 2013), mais il demeure difficile de lier cette baisse à l’effet du canal. Il est clair que ce dernier n’a pas contribué à l’établissement des pins échantillonnés car, d’une part, les âges des 11 individus ne montrent pas de signal particulier associé à l’époque de la création du canal et, d’autre part, ceux-ci sont situés
34
beaucoup trop loin du canal, soit à près de 500 mètres, pour qu’un impact ait pu se faire sentir. Au site de Lanoraie, des canaux étaient présents dans le secteur échantillonné de la tourbière. Bien que l’année de leur création est inconnue, l’âge des individus échantillonnés (avant 1950) suggère que la plupart d’entre eux étaient déjà présents avant que les canaux ne soient creusés. Des canaux sont aussi présents à la tourbière de La Durantaye, mais à plus de 100 m du peuplement étudié. Ils semblent relativement récents et leur effet fut sans doute négligeable compte tenu du fait que la tourbière montre des conditions de surface très humides et la présence d’un épais couvert de sphaignes vivantes. Enfin, aucun canal de drainage n’a été repéré dans les portions des tourbières de Villeroy et de Granby où les actuels peuplements de pins sont présents.
35
CONCLUSION
Dans le cadre de cette recherche, cinq tourbières ombrotrophes situées dans le sud du Québec et où le pin blanc est présent en tourbière et à proximité sur sol minéral furent étudiées. La présence d’îlots de pin blanc sur tourbe, dont la croissance autant en hauteur qu’en diamètre semble excellente, est en principe inhabituelle puisque la tourbe n’est pas un substrat à priori favorable pour cette espèce. Les analyses dendrochronologiques et macrofossiles réalisées dans la présente étude suggèrent que la colonisation des tourbières par le pin blanc est un phénomène relativement récent ayant surtout eu lieu au XXe siècle. De plus, bien qu’inférieure à celle sur sol minéral, la croissance radiale des individus
en tourbière s’avère néanmoins excellente. Des précipitations inférieures à la normale dans les décennies 1930 à 1970 ainsi que des températures supérieures durant les années 1930 à 1960 ont peut-être contribué à l’amorce du processus de colonisation. Cependant les fonctions de réponse et de corrélation suggèrent que le climat n’aurait pas régi la croissance radiale subséquente et ce, autant pour les individus sur tourbe que ceux sur sol minéral. Bien que le (ou les facteurs) à l’origine de l’installation du pin blanc au sein des tourbières étudiées demeure encore à être élucidé, il semble que sa présence sur les tourbières relève d’une analyse de cas par cas puisqu’il ne semble pas y avoir un seul facteur qui puisse être généralisé à l’ensemble des sites. Seul le cas de la Grande Tourbière de Villeroy apparait clair et où un feu d’abattis ayant eu lieu en 1949 est vraisemblablement à l’origine de l’actuel peuplement de pin blanc au sein de la tourbière.
Des facteurs autres que climatiques ont probablement permis au pin blanc d’afficher une excellente croissance radiale en tourbière. Puisque ces écosystèmes sont généralement des milieux ouverts ou semi-forestiers, l’intensité lumineuse au sol y est forte, ce qui fait que la lumière n’est pas limitante. Puisque les tourbières de cette recherche sont situées dans une trame agricole, des données concernant les concentrations en éléments chimiques (azote, phosphore) dans la partie supérieure de la tourbe correspondant aux deux derniers siècles pourraient permettre de voir si l’établissement et la croissance de l’espèce auraient été favorisés par des apports éoliens externes de ces nutriments. Conjointement à cela, une analyse de photographies aériennes multidates pourrait être réalisée : il serait alors possible de cartographier les modifications de l’utilisation des terres en périphérie des sites d’étude (p. ex. : sur une distance de 500 m) sur plusieurs décennies afin de voir si celles-ci sont en lien
36
avec l’établissement du pin et, de façon plus générale, au phénomène de boisement des tourbières. La présence de quelques jeunes individus dans quatre des cinq tourbières étudiées semble indiquer que les conditions ayant influencé l’établissement et la croissance des arbres sont encore présentes aujourd’hui. Il serait intéressant de prélever des jeunes individus pour produire une analyse de tige afin de vérifier si la croissance en hauteur suit le patron de croissance générale de l’espèce. Enfin, puisque l’azote est un élément limitant pour les végétaux, les pluies acides azotées pourraient aussi avoir un impact sur la croissance. Quoi qu’il en soi, plusieurs possibilités de recherches sont possibles afin d’ajouter de nouvelles connaissances et de compléter cette recherche.
37
RÉFÉRENCES
Anderson, T.W., 1995. Forest changes in the Great Lakes region at 5-7 ka BP. Géographie physique
et Quaternaire, 49: 99-116.
Avard, K., Larocque, M. & Pellerin, S., 2013. Perturbations des tourbières de la région de Bécancour, Centre-du-Québec, entre 1966 et 2010, Le Naturaliste canadien, 137: 2-15.
Beauregard, P., 2017. Dynamique du bouleau gris (Betula populifolia) dans les tourbières ombrotrophes de la Montérégie (Québec). Mémoire de M.Sc., Université Laval, Québec.
Bégeot, C. & Richard, H., 1996, L’origine récente des peuplements de Pin à crochets (Pinus uncinata Miller ex Mirbel) sur la tourbière de Frasne et exploitation de la tourbe dans le Jura, Acta Botanica
Gallica, 143 : 47-53.
Berg, E.E., McDonnell Hillman, K., Dial, R. & DeRuwe, A., 2009. Recent woody invasion of weatlands on the Kenai Peninsula Lowlands, south-central Alaska: a major regime shift after 18 000 years of wet
Sphagnum-sedge peat recruitment, Canadian Journal of Forest Research, 39: 2033-2046.
Bhiry, N. & Filion, L., 2001. Analyse des macrorestes végétaux. Dans Payette, S. & Rochefort, L. (éditeurs). Écologie des tourbières du Québec-Labrador. Les Presses de l’Université Laval, Québec, Québec, pp. 259-273.
Birks, H.H., 2013. Plant macrofossil – Introduction In Elias, S.A. (editor). Encyclopedia of Quaternary
Science. Elsevier, Edinburgh, pp. 593-612.
Bunn, A.G., 2010. Statistical and visual crossdating in R using the dplR librairy, Dendrochronologia, 28: 251-258.
Bunting, M.J., Morgan, C.R., Van Bakel, M. & Warner, B.G., 1998. Pre-European settlement conditions and human disturbance of a coniferous swamp in southern Ontario, Canadian Journal of Botany, 76: 1779-1779.
Cedro, A. & Lamentowicz, M., 2008. The last hundred years’ dendroecology of scots pine (Pinus
sylvestris L.) on a Baltic bog in the northern Poland: Human impact and hydrological changes, Baltic Forestry, 14: 26-33.
Cedro, A. & Lamentowicz, M., 2011. Contrasting responses to environmental changes by pine (Pinus
sylvestris L.) growing on peat and mineral soil: An example from Polish Baltic bog, Dendrochronologia,
29: 211-217.
Chhin, S., Chumack, K., Dhal, T., David, E.T., Kurzeja, P., Magruder, M. & Telewski, F.W., 2013. Growth-climate relationships of Pinus strobus in the floodway versus terrace forest along the banks of the Red Cedar River, Michigan, Tree-Ring Research, 69: 37-47.
38 Journal of Earth Sciences, 19 : 1938-1952.
Cubizolle, H., Thebaud, G., Seytre, L., Porteret, J. & Tourman, A., 2009-2010. Les causes du développement de l’arbre sur les tourbières du Massif Central oriental (France) au cours du XXe siècle,
Dans Cubizolle, H., Hölzler, A., Duchamp, L. & Müller, F. (éditeurs). Ecologie et protection des tourbières - Ökologie und Schutz der Moore, Annales scientifiques de la Réserve de Biosphère
transfrontalière, Vosges du Nord-Pfälzerwald, pp. 140-170.
Dauškane, I. & Elferts, D., 2011. Influence of climate on scots pine growth on dry and wet soils near lake Engure in Latvia, Estonian Journal of Ecology, 60: 225-235.
Doyon, J.P., 2016. Grande-plée-Bleue : une réserve écologique en devenir [en ligne]
https://jpdinitiateurgpb.com/
Drobyshev, I., Simard, M., Bergeron, Y. & Hofgaard, A., 2010. Does soil organic layer thickness affect climate-growth relationships in the black spruce boreal ecosystem? Ecosystems, 13: 556-574. Eckstein, J., Leuschner, H.H. & Bauerochse, A., 2011. Mid-Holocene pine woodland phases and mire development-significance of dendroeological data from subfossil trees from northwest Germany,
Journal of Vegetation Science, 22: 781-794.
Edens, D. & Ash, S., 1969. The development of a white pine stand in a bog environment at Cranberry Glades, West Virginia, Castanea, 43: 204-210.
Edvardsson, J., Poska, A., Van der Putten, N., Rundgren, M., Linderson, H. & Hammarlund, D., 2014. Late-Holocene expansion of a south Swedish peatland and is impact on marginal ecosystems: Evidence from dendrochronology, peat stratigraphy and palaeobotanical data, The Holocene, 24: 466-476.
Filion, L., 1987. Holocene development of parabolic dunes in the central St. Lawrence lowland Québec,
Quaternary Research, 28: 196-209.
Freléchoux, F., Buttler, A., Schweingruber, F.H. & Gobat, J-M., 2000. Stand structure, invasion, and growth dynamics of bog pine (Pinus uncinata var. rotundata) in relation to peat cutting and drainage in the Jura Mountains, Switzerland, Canadian Journal of Forest Research, 30: 1114-1126.
Gałka, M., Miotk-Szpiganiwicz, G., Marczewska, M., Barabach, J., van der Knaap, W.O. & Lamentowicz, M., 2015. Palaeoenvironmental changes in Central Europe (NE Poland) during the last 6200 years reconstructed form a high-resolution multi-proxy peat archive, The Holocene, 25: 421-434. Garneau, M. & Lamarre, A., 2013. Dynamique paléohydrologique de la tourbière de la Grande-plée-Bleue. Rapport final présenté au Ministère du Développement Durable, de l’Environnement, de la Faune et des Parcs du Québec, 31 p.
Gosselin, J., 2005. Guide de reconnaissance des types écologiques de la région écologique 2b – Plaine du Saint-Laurent. Ministère des Ressources naturelles et de la Faune du Québec, Direction des inventaires forestiers, Division de la classification écologique et productivité des stations.