Structure and function of mitochondrial small heat shock protein 22 in Drosophila
melanogaster
Thèse
Afrooz Dabbaghizadeh
Doctorat en biologie cellulaire et moléculaire Philosophiae Doctor (Ph. D.)
Québec, Canada
Structure and function of mitochondrial small heat shock protein 22 in
Drosophila melanogaster
Thèse
Afrooz Dabbaghizadeh
Sous la direction de :
Résumé
Les petites protéines de choc thermique (sHsps) ont été découvertes initialement chez
Drosophila. Les membres de cette famille sont des chaperons moléculaires sont présents
dans la plupart des organismes eucaryotes et procaryotes et certains virus. En plus d’être induites en réponse à la plupart des stresseurs dont un choc thermique, elles sont également exprimés en absence de stress. Les sHsps forment des structures dynamiques s'assemblant en oligomères et elles sont essentielles durant les conditions de stress en empêchant l'agrégation des protéines dénaturées et en favorisant leur repliement par des chaperons moléculaires dépendants de l'ATP.
Le génome de Drosophila melanogaster code pour 12 sHsp, qui ont des profils d'expression développementaux, des localisations intracellulaires diverses et des spécificités de substrats distincts. DmHsp22 est jusqu'à présent la seule sHsp localisée dans les mitochondries avant et après un choc thermique. Elle est préférentiellement régulée lors du vieillissement et en réponse à la chaleur et aux stress oxydants. La surexpression de DmHsp22 augmente la durée de vie et la résistance au stress et sa régulation négative est préjudiciable. C'est un chaperon efficace, qui pourrait être impliqué dans la réponse mitochondriale au dépliement protéique (UPRMT). Cependant, le mécanisme exact de son action est mal compris.
Structurellement, DmHsp22 forme une population d'oligomères semblable aux nombreux sHsps de métazoaires et différente de DmHsp27. L'alignement des séquences de la région ACD de DmHsp22 avec des sHsp de drosophile et d'autres organismes a démontré la présence de trois résidus d'arginine hautement conservés dans ce domaine. Une forte conservation de ces résidus suggère leur implication possible dans la structure et la fonction de DmHsp22. La substitution des résidus d'arginine hautement conservés dans les sHsps de mammifères est associée à certaines pathogenèses et déclenche des changements de conformation des protéines ainsi que l'agrégation des protéines intracellulaires. La mutation de l'arginine en glycine au niveau de trois résidus hautement conservés d'ACD dans DmHsp22 (R105, R109, R110) résulte en une population oligomérique qui, dans le
IV
Bien que DmHsp22 ainsi que les mutants aient été caractérisés comme des chaperons efficaces in vitro, les mécanismes d'action exacts dans les mitochondries et l'information sur le comportement protecteur nécessitent la détermination du réseau d’interaction in vivo. Nous avons utilisé la technique capture d’immunoaffinité (CIA) pour récupérer 60 protéines qui interagissent spécifiquement avec DmHsp22 in vivo pendant le traitement normal et thermique, dans le surnageant des cellules de mammifères exprimant la DmHsp22. L’CIA effectuée sur la fraction mitochondriale a permis d’identifies 39 protéines qui interagissent spécifiquement avec DmHsp22.
La combinaison de l’IAC avec l'analyse par spectroscopie de masse de mitochondries de cellules HeLa transfectées avec DmHsp22 a conduit à l'identification de partenaires de liaison à DmHsp22 dans des conditions de normales et de choc thermique. L'interaction entre DmHsp22 et deux autres chaperons mitochondriaux a été validée par immunobuvardage. Notre approche a montré que les cellules HeLa exprimant DmHsp22 augmentent la consommation d'oxygène mitochondrial et les teneurs en ATP, ce qui confère un nouveau rôle à DmHsp22 dans les mitochondries. En outre, l'activité d’une luciférase exogène a légèrement augmenté dans les cellules HeLa exprimant DmHsp22 après que l'activité enzymatique ait été réduite à la suite de l'exposition à la chaleur.
En résumé, ce projet a permis de caractériser la structure oligomérique de DmHsp22 et un certain nombre de mutants dans le domaine alpha cristallin tout en fournissant un rôle potentiel mécanistique dans l’homéostase mitochondriale. La détermination du réseau mitochondrial de DmHsp22 suggère son importance dans cette organelle non seulement en tant que chaperon moléculaire, mais aussi en tant que protéine impliquée dans plusieurs fonctions cellulaires significatives.
Abstract
The small heat shock proteins (sHsps) were first discovered in Drosophila. Members of this family are molecular chaperones and are present in most eukaryotic and prokaryotic. Although, they are induced in response to most of the stressors including heat shock, they are also expressed in absence of stress. SHsps form dynamic structures that assemble into oligomers which are essential during stress conditions by preventing aggregation of denatured proteins and promoting their folding by ATP dependent molecular chaperones.
Drosophila melanogaster genome encodes 12 sHsps, that have developmental
expression patterns, diverse intracellular localizations and distinct substrate specificities. DmHsp22 is up to now the only sHsp localized in mitochondria before and after heat shock. It is preferentially regulated during ageing and in response to heat and oxidative stresses. Over-expression of DmHsp22 increases lifespan and resistance to stress and its down-regulation is detrimental. It is an efficient chaperone and could be involved in the mitochondrial unfolding protein response (UPRMT). However, the exact mechanism of its action is poorly understood.
Structurally, DmHsp22 forms one population of oligomers similar to the many metazoan sHsps but DmHsp27. Sequence alignment of DmHsp22 with sHsps in
Drosophila and other organisms at the alpha crystalline domain (ACD) region
demonstrated the presence of three highly conserved arginine residues in this domain. Strong conservation of these residues suggest their possible involvement in structure and function of DmHsp22. Substitution of highly conserved arginine residues in mammalian sHsps is associated with some pathogenesis and triggers protein conformational changes as well as intracellular protein aggregation. Mutation of arginine to glycine at three highly conserved residues of ACD in DmHsp22 (R105, R109, R110) results in one oligomeric population as well which in the case of R110G disrupts the structure and causes formation of smaller oligomers.
VI
Although DmHsp22 as well as mutants have been characterized as effective in vitro chaperones, the exact mechanism(s) of action in mitochondria and information about protective behavior requires defining of in vivo protein interacting network. We have used immunoaffinity conjugation (IAC) technique to recover 60 proteins that specifically interact with DmHsp22 in vivo during normal and heat treatment using cell extract of mammalian cells expressing DmHsp22. The IAC performed on mitochondrial fraction identified 39 proteins that specifically interact with DmHsp22.
Combination of IAC with mass spectroscopy analysis of mitochondria of HeLa cells transfected with DmHsp22 resulted in identification of DmHsp22-binding partners under normal and under heat shock conditions. Interaction between DmHsp22 and two other mitochondrial chaperones was validated by immunoblotting. Our approach showed that HeLa cells expressing DmHsp22 increase maximal mitochondrial oxygen consumption and ATP contents which provides a new mechanistic role for DmHsp22 in mitochondria. Furthermore, exogenous luciferase activity slightly increased in HeLa cells expressing DmHsp22 after the enzyme activity reduced as a result of exposure to heat.
In summary, this project has characterized the oligomeric structure of DmHsp22 and a number of mutants in the alpha crystalline domain while providing a potential mechanistic role in mitochondrial homeostasis. Determining mitochondrial network of DmHsp22 suggest its importance in this organelle not only as a molecular chaperone but also as a protein involved in several significant cellular functions.
Table of Contents
Résumé ... III
Abstract ... V
Table of Contents ... VII
List of tables ... XIV
List of figures ... XV
List of Abbreviations and Acronyms ... XVIII
Acknowledgements ... XXII
Foreword ... XXIV
1 Introduction ... 1
1.1 Heat shock proteins ... 1
1.1.1 Heat shock as cellular stressor ... 1
1.1.2 Appearance of heat shock proteins ... 1
1.1.3 Regulation of heat shock response ... 2
1.1.4 Heat shock proteins as molecular chaperones ... 3
1.2 Small heat shock proteins (sHsps) ... 5
1.2.1 Primary structure of small Hsps ... 6
1.2.2 Dynamic behavior of sHsps ... 10
1.2.3 Chaperone activity and substrate recognition by sHsps ... 14
1.2.4 Regulation of sHsps activity ... 19
VIII
1.3 The response to stress in Drosophila melanogaster ... 26
1.3.1 The sHsps in Drosophila melanogaster ... 26
1.3.2 Developmental expression and function of D. melanogaster sHsps ... 29
1.3.3 Cellular localization of D.melanogaster sHsps ... 31
1.3.4 Expression of D. melanogaster sHsps in Adult ... 32
1.3.5 Thermal induction of D.melanogaster sHsps ... 32
1.3.6 Drosophila melanogaster Hsp22 ... 33
1.3.7 Problems and Objectives ... 35
Chapter 2. Oligomeric structure and chaperone-like activity of Drosophila melanogaster mitochondrial small heat shock protein Hsp22 and arginine mutants in the alpha-crystallin domain ... 36
Résumé ... 37
Abstract ... 38
2 Oligomeric structure and chaperone-like activity of Drosophila melanogaster mitochondrial small heat shock protein Hsp22 and arginine mutants in the alpha-crystallin domain ... 39
2.1 Introduction ... 39
2.2 Materials and methods ... 41
2.2.1 Amino acid sequence alignment ... 41
2.2.2 Cloning and expression of recombinant proteins... 41
2.2.3 Purification of recombinant proteins ... 42
2.2.4 Fluorescence spectroscopy ... 42
2.2.6 Size Exclusion Chromatography (SEC) ... 43
2.2.7 SDS-PAGE and Native gels ... 44
2.2.8 Heat-induced aggregation assay ... 44
2.2.9 DTT-induced Insulin assay ... 44
2.3 Results ... 45
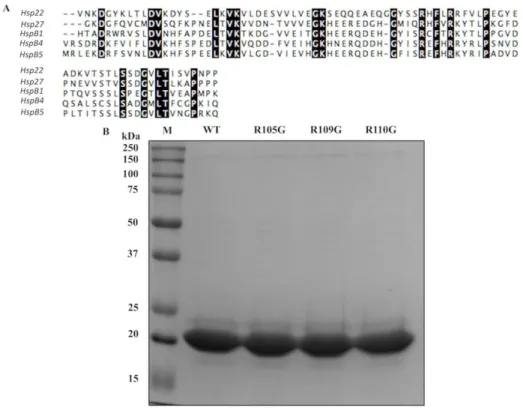
2.3.1 Sequence alignments and protein production ... 45
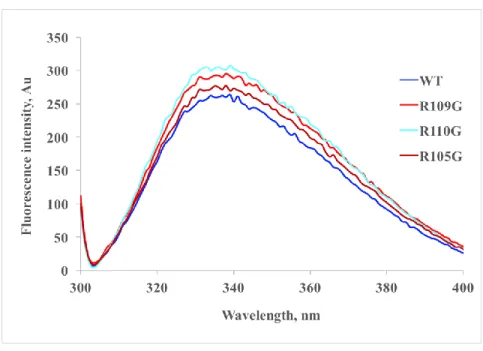
2.3.2 Fluorescence spectra of the DmHsp22WT and its ACD arginine mutants 47 2.3.3 Thermal transition of Hsp22 and its ACD arginine mutants ... 49
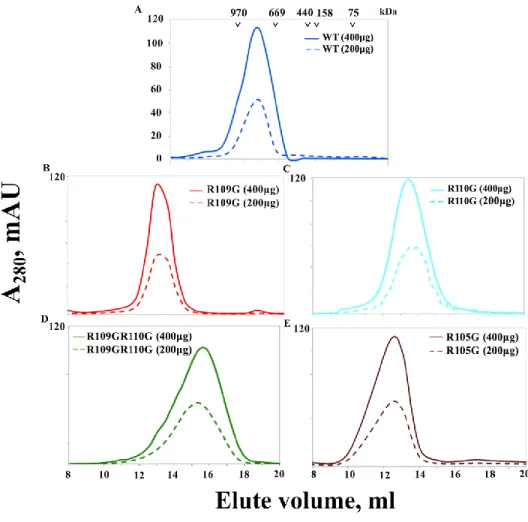
2.3.4 Mutation of arginine residues in the ACD influences the oligomeric structure of DmHsp22 ... 51
2.3.5 Chaperone-like activity ... 55
2.4 Discussion ... 58
2.5 Acknowledgements ... 60
2.6 References... 61
Chapter 3. Identification of proteins interacting with the mitochondrial small heat shock protein hsp22 of Drosophila melanogaster: implication in mitochondrial homeostasis ... 68
Résumé ... 69
Abstract ... 70
3 Identification of proteins interacting with the mitochondrial small heat shock
protein hsp22 of Drosophila melanogaster: implication in mitochondrial homeostasis
X
3.2 Materials and methods ... 73
3.2.1 Construction of expression vectors ... 73
3.2.2 Cell culture and transfection ... 74
3.2.3 Immunofluorescence microscopy ... 74
3.2.4 Cellular fractionation ... 74
3.2.5 Electrophoresis and western blot analysis ... 75
3.2.6 Immuno affinity capture experiments ... 76
3.2.7 Mass Spectrometry analysis ... 76
3.2.8 Mitochondrial oxygen consumption ... 77
3.2.9 ATP content measurements ... 78
3.2.10 Luciferase reactivation assay ... 78
3.3 Results ... 80
3.3.1 Mitochondrial localization of DmHsp22-Flag in transiently transfected HeLa cells 80 3.3.2 Identification and characterization of DmHsp22-associated proteins ... 82
3.3.3 DmHsp22 interacts with proteins involved in different molecular functions 89 3.3.4 ATP synthase subunits are the main DmHsp22-binding proteins ... 91
3.3.5 Heat shock influences stability of mitochondrial partners of DmHsp22 .... 93
3.3.6 DmHsp22 effects on mitochondrial oxygen consumption and ATP content of cells 96 3.3.7 DmHsp22 is an efficient chaperone in human cells... 98
3.5 Acknowledgements ... 103
3.6 References... 104
3.7 Supporting information ... 110
4 Analysis of mitochondrial partners of DmHsp22 at physiological conditions 113 4.1 Introduction ... 113
4.2 Material and methods ... 114
4.2.1 Cell culture and transfection ... 114
4.2.2 IAC on pure mitochondria fraction ... 114
4.2.3 Mass Spectrometry analysis ... 114
4.3 Results ... 118
4.3.1 Identification and characterization mitochondrial DmHsp22 interactors . 118 4.3.2 Distribution of DmHsp22-associated proteins at different mitochondrial sub-compartments ... 118
4.3.3 Bioinformatics analysis and functional Annotation of DmHsp22-associated proteins 120 4.4 Conclusion ... 120
5 General discussion and conclusion ... 124
5.1 DmHsp22 and arginine mutants of ACD are mitochondrial sHsps .... 124
5.2 Oligomeric structure of DmHsp22 at physiological condition and in response to temperature changes ... 126
XII
5.4 A general overview on the DmHsp22-associated partners in
mitochondria of cells at physiological and heat shock conditions ... 131
5.5 From mitochondrial network of DmHsp22 to its influence on mitochondrial respiration and effect as mitochondrial chaperone ... 132
5.6 Immunoaffinity on pure mitochondria confirms interaction of DmHsp22 and some of the mitochondrial partners ... 133
5.7 References... 136
Appendix A. Materials and Methods ... 175
A.1 Cloning, site directed mutagenesis, expression and purification of vectors 175 A.2 Proteomic approaches ... 178
IPTG induction of protein expression ... 178
Determining the protein solubility ... 179
Purification of recombinant proteins ... 180
Expression and Purification of SUMO hydrolysis enzyme ... 184
Size Exclusion Chromatography (SEC) ... 185
SDS-PAGE, native electrophoresis and western blot ... 187
Measuring protein concentration ... 188
Chaperone assays to prevent aggregation of different substrates ... 189
A.3 Fluorescence spectroscopy ... 191
A.4 Dynamic Light Scattering (DLS) ... 191
A.5 Cell culture and transfection ... 192
Indirect immunofluorescence... 194
A.6 Immuno-Affinity Conjugation (IAC) ... 195
Sample preparation for IAC ... 196
XIV
List of tables
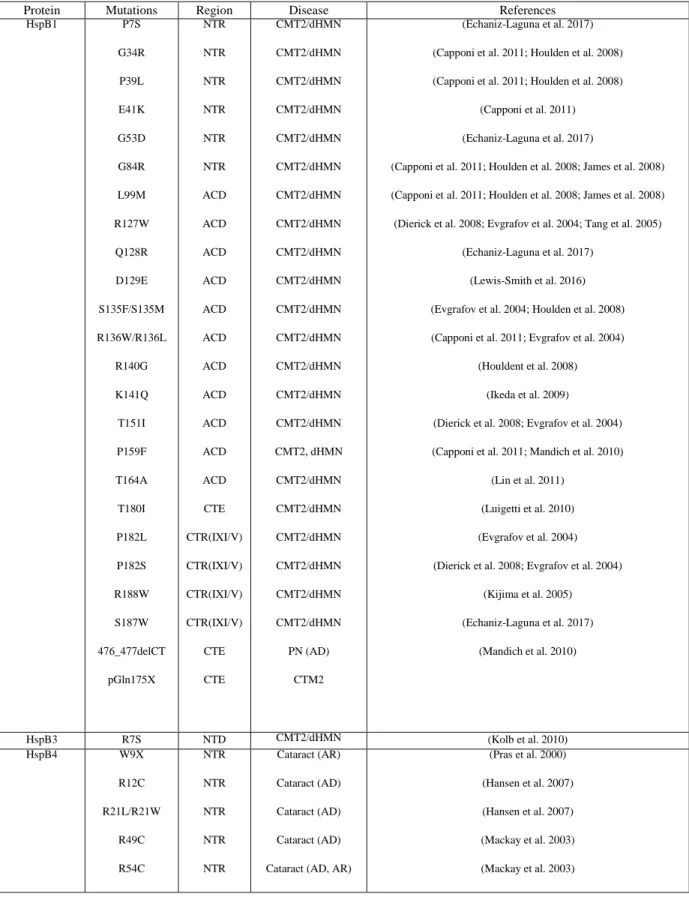
Table 1-1 Diseases-associated mutations in human sHsps ... 24
Table 1-2 Drosophila melanogaster sHsps characteristics ... 27
Table 2-1 Primers to construct Drosophila melanogaster Hsp22 arginine mutants ... 41
Table 3-1 Forward and reverse primers applied to the constructs of pcDNAHsp22 wild type, pcDNAHsp22WT-Flag and pcDNA-Flag ... 73
Table 3-2 Proteins interacting with DmHsp22 by orbitrap fusion LC-MS/MS mass
spectrometer... 85
Table 4-1 DmHsp22-interacting proteins detected through MS ... 115
Table 5-1 Thermal transition of DmHsp22WT and arginine mutants... 127
Table A-1 Primers designed to construct different mutants of DmHsp22 in the region ACD ... 177
List of figures
Figure 1.1 Primary structure of sHsps ... 10
Figure 1.2 Two ways of dimerization in sHsps ... 12
Figure 1.3 Comparison of the three available oligomeric structure of sHsps ... 14
Figure 1.4 Model for chaperone function of Shsps ... 18
Figure 1.5 Multiple sequence alignment of full length of DmHsp22 with sHsps from other organisms ... 28
Figure 2.1 Multiple sequence alignment of full length of DmHsp22 with sHsps from other organisms ... 46
Figure 2.2 Fluorescence spectra of the DmHsp22WT and its arginine mutants ... 48
Figure 2.3 DLS analysis of DmHsp22WT and arginine mutants of ACD shows their behavior by increasing the temperature ... 50
Figure 2.4 SEC profiles of the DmHsp22WT and arginine mutants ... 53
Figure 2.5 Novex™ 4-12 % Tris-Glycine native gel analysis ... 54
Figure 2.6 Chaperone-like activity of DmHsp22WT and its arginine mutants by using luciferase (Luc) (A and B) and Insulin (C and D) as protein substrates ... 56
XVI
Figure 2.7 Chaperone-like activity of DmHsp22WT and arginine mutants using citrate synthase (CS) (A and B) and malate dehydrogenase (MDH) (C and D) as protein
substrates ... 57
Figure 3.1 Characterization of DmHsp22WT and DmHsp22WT-Flag expressing cells ... 81
Figure 3.2 Identification of DmHsp22-associated proteins ... 84
Figure 3.3 Gene Ontology analysis of DmHsp22-associated proteins ... 91
Figure 3.4 Abundancy of ATP synthase (Complex V) subunits and subunit composition of DmHsp22-associated partners ... 93
Figure 3.5 Heat shock and recovery does not influence expression levels of DmHsp22. Confirmation of DmHsp22’s interaction with Hsp60 and Hsp70 ... 95
Figure 3.6 Mitochondrial oxygen consumption in intact cells and cellular ATP concentration ... 97
Figure 3.7 DmHsp22 is an important factor in thermotolerance under heat stress condition ... 99
Figure 3.8 Biological relevance of DmHsP22-associated proteins ... 110
Figure 3.9 Mitochondrial distribution and biological relevance of DmHsp22-associated proteins following heat shock exposure and recovery ... 111
Figure 4.1 Mitochondrial distribution of DmHsp22-associated proteins ... 118
Figure 4.3 GO molecular functions of DmHsp22-interacting proteins ... 121
Figure 4.4 GO Protein class of DmHsp22-associated proteins ... 122
Figure 5.1 Intracellular localization of DmHsp22 and arginine mutants ... 125
Figure 5.2 Schematic overview of mitochondrial proteins interacting DmHsp22 ... 133
Figure A.1 Cloning cDNA of DmHsp22 into the pETHSUK expression vector ... 176
Figure A.2 Induction of DmHsp22WT and arginine mutants in E.coli using 0.4 mM IPTG ... 179
Figure A.3 SDS-PAGE of protein solubility test ... 180
Figure A.4 Schematic overview of expression and purification of recombinant proteins using Ni-NTA column ... 182
Figure A.5 Cutting 6His-SUMO tag from N-terminal of DmHsp22 by SUMO hydrolase enzyme ... 183
Figure A.6 Preparative size exclusion chromatography ... 184
Figure A.7 Recombinant purified SUMO hydrolysis enzyme on SDS-PAGE ... 185
Figure A.8 Principle of in vitro chaperone-like activity against different substrates. OD is optical density ... 190
Figure A.9 Cloning cDNA of DmHsp22 and Flag tag into the pcDNA mammalian expression vector ... 193
XVIII
List of Abbreviations and Acronyms
°C: Degree Celsius
ACD: Alpha crystalline domain
ANT: Mitochondrial adenine nucleotide translocase ATP: Adenosine three phosphate
BSA: Bovine serum albumin CD: Circular dichroism cDNA: Complementary DNA
ClpP: ATP-dependent Clp protease proteolytic subunit CMT2: Charcot-Marie-Tooth neuropathy
CO2: Carbon dioxide
CS: Citrate synthase CTD: C-terminal domain CTE: C-terminal extension
DAPI: 4',6-Diamidino-2-Phenylindole, Dihydrochloride DHMN: Distal hereditary motor neuropathy
DLS: Dynamic light scattering Dm: Drosophila melanogaster
DMEM: Dulbecco’s Modified Eagle medium DNA: Deoxyribonucleic acid
DRM: Desmin related myopathy DTT: Dithiothreitol
ER: Endoplasmic reticulum ETC: Electron transport chain
F0: Factor sensitive to the oligomycine
F1: Factor 1
FBS: Fetal bovine serum
FOXO: Fork head transcription factor
FRET: Fluorescence resonance energy transfer Fwd: Forward
H2O: Hydrogen oxide
H2O2: Hydrogen peroxide
HAF-1: Transcription initiation factor-1 HSE: Heat shock element
Hsp: Heat shock protein HSR: Heat shock response
IGC: Interchromatin granule clusters IgG: Immunoglobulin G
IMM: Inner Mitochondrial membrane IMS: Inter membrane space
IPTG: Isopropyl ß-D-1 thiogalactopyranoside KDa: Kilo Dalton
LB: Luria Bertani Broth Luc: Luciferase
MDH: Malate dehydrogenase Mg/ml: Milligrams/Milliliters
XX mRNA: Message Ribonucleoprotein
MS: Mass spectroscopy MW: Molecular Weight NaCl: Sodium Chloride
Ni-NTA: Nickel Nitrilo Triacetic Acid NLS: Nucleus localization sequence
NMR: Nuclear magnetic resonance spectroscopy NTD: N-terminal domain
O2: Oxygen molecule
OCR: Oxygen consumption rate OD: Optical density
OMM: Outer mitochondrial membrane OPA1: Optic atrophy 1 protein
OXPHOS: Oxidative phosphorylation P3000: Pellet 3000
PQC: Protein quality control PCR: Polymerase chain reaction PMSF: Phenylmethylsulfonyl fluoride PSI: Pound per square inch
PTMs: Post translational modifications Rev: Reverse
Rh: Hydrodynamic radius
RIPA: Radio immunoprecipitation assay buffer RNA: Ribonucleic acid
ROS: Reactive oxygen species RPM: Rotation per minute RT: Room temperature S3000: Supernatant 3000 SB: Sample buffer
SDS-PAGE: Sodium dodecyl sulfate Polyacrylamide gel electrophoresis SDS: Sodium dodecyl sulfate
SEC: Size exclusion chromatography SHsp: Small Heat shock protein
SOC: Super optimal broth with catabolite repression SUMO: Small ubiquitin related modifier
TH: Total homogenate
UPR: Unfolding protein response UV: Ultraviolet
XXII
Acknowledgements
This thesis is one of my major accomplishments in life and would not have been possible without the love, support and patience of my family, friends and advisors. Hereby, I would like to thank my fantastic advisor, Professor Robert M. Tanguay who always encouraged me with a smile and gave me more confidence and strength during my PhD study at Laval University. I was very lucky to get a chance to have you as my advisor. Also, my genuine appreciations to Dr. Geneviève Morrow for her extreme patience and support especially during the first year of my studies. Thank you both for constructively ignoring my mistakes and highlighting my strengths to grow up in your team. I am quite in debt for all your unsparing and precious help and guidance.
Special thanks to my committee members, Professor. Josée N. Lavoie, Professor. Edward Khandjian and Professor Lawrence Hightower for accepting to review this project and for their attendance in my defense session. I would also like to thank my lab colleagues from the beginning until now for their helps and supports during my PhD including: Dr. Mohamed Taha Moutaoufik, Maxime Bourrelle-Langlois, Vincent Roy, Idalia Castro, Dr. Francesca Angileri and Dr. Ornella Pellerito. Special thanks to my friend and coworker Dr. Mohamed Taha Moutaoufik for kindly translating the abstracts of this thesis into French.
Moreover, I would also like to underline the kindness of Dr. Rong Shi for allowing me to use the Fast Protein Liquid Chromatography in his laboratory and Jérémie Hamel for providing the required training. I must also acknowledge Dr. Sylvie Bourassa from Proteomics platform of CHU de Quebec Research Center-CHUL for help with Mass Spectrometry experiments.
I would like to thank Dr. Nicolas Pichaud and Dr Étienne Hébert Chatelain who hosted me in their laboratory at the Department of Chemistry and Biochemistry and Department of Biology at University of Moncton. Thanks to all very kind lab members especially Yasmine Ould Amer for their supports. I would like to thank Dr. Stéphanie Finet who performed DLS assay in her laboratory at the Institute of Mineralogy, Physics of Materials and Cosmo-chemistry (IMPMC), Pierre-et-Marie-Curie University (Paris, France).
I am truly thankful to my dear brother Erfan Dabbaghizadeh for his presence in Quebec and helping me through dilemmas and also my best friend Banin Alizadeh Sahraei for being like a sister to me and for all the good moments we experienced together.
Finally, very special thank from bottom of my heart to my love and my dear friend and husband Ali Faridkhou for your unconditional support and understanding and for your constant source of assurance. We started this journey together and grew up gradually and learned a lot in this way. I love you so much my dearest. I dedicate this thesis to you.
XXIV
Foreword
As part of this doctoral project, I had the opportunity to work at the Institute of Integrative Biology and Systems (IBIS) of Laval University on the Study the structure and function of mitochondrial small heat shock protein 22 in Drosophila melanogaster. This PhD thesis comprises five chapters starting from introduction as chapter one and ending with general discussion and conclusion as chapter 5. Appendix has been inserted at the end of this thesis.
Chapter 2 is based on the following article: “Oligomeric structure and chaperone-like activity of Drosophila melanogaster mitochondrial small heat shock protein Hsp22 and arginine mutants in the alpha-crystallin domain, Afrooz Dabbaghizadeh, Stéphanie Finet, Genevieve Morrow, Mohamed Taha Moutaoufik and Robert M Tanguay., Cell Stress and Chaperone, 2017, 22 (4) 577-588”. All the experiments presented in this article except for DLS including: cloning of DmHsp22 into the pETHSUK vector, mutagenesis, expression, purification as well as biochemical, bioinformatic and biophysical analyzes were performed by me. I also prepared the article in its published form along with figures and tables. Stéphanie Finet and Mohamed Taha Moutaoufik performed DLS experiment. Mohamed Taha Moutaoufik also helped me for designing figure 1a. All authors read and provided comments in the preparation of this manuscript. Geneviève Morrow and Robert M Tanguay supervised the entire work.
Chapter 3 is based on the following article: “Identification of proteins interacting with the mitochondrial small heat shock protein hsp22 of Drosophila melanogaster: implication in mitochondrial homeostasis, Afrooz Dabbaghizadeh, Geneviève Morrow, Yasmine Ould Amer, Etienne Hebert Chatelain, Nicolas Pichaud and Robert M. Tanguay., PLoS ONE, 2018, 6; 13(3)”. All the experiments including cloning, mutagenesis, cell culture, fluorescent microscopy, IAC, respiration and MS analysis were performed by Afrooz Dabbaghizadeh. Mass spectroscopy was performed by the Proteomics platform of CHU de Quebec. I also wrote the article in its final form while Yasmine Ould Amer, Etienne Hebert Chatelain and Nicolas Pichaud helped me in structuring figure 6. All the coauthors provided their feedback and comments for this work. Geneviève Morrow and Robert M. Tanguay supervised the work.
Chapter 4: All the experiments performed in this chapter, IAC and MS analysis were performed by Afrooz Dabbaghizadeh under the supervision of Robert M. Tanguay. MS was performed by the Proteomics platform of CHU de Quebec.
Chapter 5 is a general discussion and conclusion which has been written by Afrooz Dabbaghizadeh. All the figures and table in this chapter where produced by me. Table 5.1 was built according to the DLS results which Stéphanie Finet and Mohamed Taha Moutaoufik performed for me.
Appendix A is a general summary of all the techniques I have used during the process of this PhD. All the figures and schematic overviews have been produced by Afrooz Dabbaghizadeh.
1 Introduction
1.1 Heat shock proteins
1.1.1 Heat shock as cellular stressor
Hyperthermia (heat stress) is a strong cellular stress (Ivanova et al. 2015). Although organisms have adapted to live in their specific optimal temperature ranging from the freezing point of water to 113°C, still heat is a major stressor which triggers a survival problem (Lindquist 1986; Stetter 2006). Fruit fly Drosophila was the first organism in which the effect of heat was investigated (Ritossa 1962; Hightower 1990). Elevated temperature of just a few degrees causes denaturation and aggregation of proteins and leading to the significant damages to cellular structures and functions (Brown and Lupas 1998; D'Amico et al. 2006; Takai et al. 1998). Heat shock disrupts cytoskeleton reorganization of actin filaments to aggregation of filament-forming proteins and collapse of cytoskeleton network (Toivola et al. 2010; Welch and Suhan 1985; Welch and Suhan 1986). Stress condition causes decrease in the number and integrity of mitochondria and lysosomes and leads to the fragmentation of endoplasmic reticulum (ER) and Golgi system (Welch and Suhan 1985). Dropping mitochondrial number and disruption in oxidative phosphorylation dramatically decrease ATP levels (Lambowitz et al. 1983; Patriarca and Maresca 1990). RNA splicing is strongly affected by heat stress (Vogel et al. 1995). Nucleoli swells and forms large granular depositions which become visible aggregates (Welch and Suhan 1985). Cytosolic stress granules due to the heat dramatically influence mRNA function and decrease translation (Nover et al. 1989). Heat shock disrupts iron homeostasis and influences cytosolic pH by affecting cellular membrane (Coote et al. 1991; Piper et al. 2003). Together, these effects stop growth and proliferation and cause cell-cycle arrest (Yost and Lindquist 1986). To inhibit fatal consequences, cells start the expression of heat shock proteins (Hsps) which are highly conserved proteins involved in maintaining protein homeostasis (De Maio et al. 2012).
1.1.2 Appearance of heat shock proteins
including oxidative stress, protein aggregation, heavy metal, ethanol, virus infections and many other toxic materials (Ahn and Thiele 2003; Bournias-Vardiabasis and Buzin 1986; Courgeon et al. 1984; Heikkila et al. 1982; Michel and Starka 1986).
It was 55 years ago (in 1962), that Ferrucio Ritossa noticed the appearance of unique puffing pattern in the salivary gland of larvae of Drosophila busckii as a result of accidental elevation in incubator temperature (Jamrich et al. 1977; Ritossa 1966). Puffs are the main sites for synthetic activity especially RNA production. Distribution of puffs of salivary glands of 2L 14 and 2L 15 chromosomes of Drosophila busckii obviously were changed following elevated temperature from 25 to 30 °C or more for 30 min. Expression of these puffs is induced within a few minutes and in response to the many stressors (Berendes 1968; Leenders and Berendes 1972; Ritossa 1966). These changes followed by the synthesis of heat shock proteins or chaperones, which are among the most conserved proteins known, was called heat shock response (Tissieres et al. 1974). Tissières and Mitchell reported the rapid appearance of six strong protein bands following heat shock and suggested their association with heat-induced puffs (Tissieres et al. 1974).
In 70s, focusing on the transcribed corresponding gene and related proteins were observed in yeasts (Peterson et al. 1979). Short time later, these family were revealed to be related to the resistance against cellular stress in the cell (Landry et al. 1982).
Observing the similar phenomena in large variety of organisms as a general phenomenon of cellular adaptation to the high temperature (thermotolerance) and presence of conserved heat shock genes through evolution, heat shock response was considered a universal mechanism (Kelley and Schlesinger 1978; Lemaux et al. 1978; Lindquist 1986; McAlister and Finkelstein 1980). From that date up, investigation in this field was expanded in different directions.
1.1.3 Regulation of heat shock response
Heat shock transcription factors (HSFs) were discovered 30 years ago (Parker and Topol 1984). They are largely conserved from fungi to humans and regulate gene expression of heat shock proteins at the transcriptional level (Akerfelt et al. 2010; Anckar
3
and Sistonen 2011). They activate expression of chaperones by recognizing promoter binding sites referred to as heat shock elements (HSEs) which is composed of multiple inverted repeats in response to conditions that caused misfolding (Pelham 1982; Sakurai and Enoki 2010; Vihervaara et al. 2013). HSEs are targets of heat shock factors within promoter region of HS genes (Mendillo et al. 2012; Xiao et al. 1991) and consequently induce synthesis of HSPs. HSF1 and HSF2 are the most studied HSFs in human due to their established role in heat shock gene expression and their link to diseases such as neurodegenerative diseases (Akerfelt et al. 2010; Gomez-Pastor et al. 2017). In bacteria, Hsp70/Hsp40, Dnak and its cofactor DnaJ bind to the transcription factor sigma32 and make it inactivated, trigger its degradation and finally reduce its cellular level (Rodriguez et al. 2008). Upon stress condition, chaperones are more integrated in the unfolded proteins and are less available to bind to the transcription factor sigma32. Thus, sigma32 becomes active and moves to the nucleus and simulates RNA polymerase and transcription (Rodriguez et al. 2008; Voellmy and Boellmann 2007). In eukaryotes, monomeric form of HSF1 in complex with Hsp70 and Hsp90 is inactivated (Non-DNA-binding state) (Boyault et al. 2007; Zou et al. 1998). Repressive complex is disrupted in stress condition and causes disassociation of Hsp90 and activation of HSF1 (Mendillo et al. 2012). This causes trimerization, phosphorylation and translocation of HSF1 from cytosol to the nucleus (trimeric DNA-binding state) and its binding to HSEs (Akerfelt et al. 2010; Prahlad et al. 2008). The maximum amount of transcription is achieved only when HSF1 trimers remain bound to the HSEs. Reducing stress signals or damaged proteins rapidly reduce the heat-shock response (Abravaya et al. 1991; Yao et al. 2006). Deregulation of HSFs activity lead to the many pathological conditions (Gomez-Pastor et al. 2017; Kim et al. 2016). Some cancers are accompanied with elevated level of HSFs (Mendillo et al. 2012; Scherz-Shouval et al. 2014).
1.1.4 Heat shock proteins as molecular chaperones
Unfolded proteins tend to form large aggregates that severely impact normal cellular functions. The main function of Heat shock proteins (Hsps)/chaperones is to act as a buffer to limit misfolding and correct aggregated proteins to reduce the environmental and genetic effects on the proteome (Jacob et al. 2017). Indeed, they are involved in maintaining
intracellular protein homeostasis (Alderson et al. 2016; Chiti and Dobson 2006; Dimauro et al. 2016; Dobson 2003; Morimoto 2008; Tyedmers et al. 2010).
Molecular chaperones are necessary not only for de novo protein folding but also for refolding of nonnative polypeptide chains (Kerner et al. 2005; Mayer 2010). Under stress conditions the number of unfolded and aggregated proteins is elevated (Tyedmers et al. 2010). Exposure of hydrophobic amino acids is characteristic of nonnative proteins that make them recognizable by chaperones. Release of the substrates from chaperones in an ATP-dependent manner finally leads to reform native protein structure with correct folding (Narberhaus 2002; Richter et al. 2010).
Heat shock proteins are subdivided into five main conserved sub families according to their molecular masses and their sequence homologies; Hsp100 kDa (Hsp 100s), Hsp90 (85-90 kDa), Hsp70; DnaK (68-73 kDa), Hsp60; GroEL and the family of small heat shock proteins (12-42 kDa) (Park et al. 2007).
Hsp90 is one of the most abundant cytosolic chaperones under the physiological condition which makes up 1-2 % of the total cytosolic protein content in eukaryotes and is upregulated about two folds following stress (Buchner 1999; Krukenberg et al. 2011; Welch and Feramisco 1982; Whitesell and Lindquist 2005). Huge number of proteins work together in Hsp90 chaperone system in ATP-dependent manner including Sti1 and prolyl isomerase Cpr6 (Hessling et al. 2009; Richter et al. 2003). Hsp90 along with its co-chaperones has significant role in folding of at least 200 specific proteins from various signaling pathways (Csermely et al. 2005; Csermely et al. 1998).
Hsp100 is in the family of AAA ATPase group and divided into two subfamilies according to the number of AAA domains (Martin et al. 2005). Hsp100 family are essential components of the protein quality control (PQC) and play crucial role in development (Mogk et al. 2015). They act in major metabolic pathways to reactivate or degrade misfolded enzymes following environmental stresses (Pulido et al. 2016). Hsp100 (ClpB/Hsp104) acts together with Hsp70 to degrade toxic protein aggregates through pulling unfolded proteins into the central pore of hexameric ring and trigger their refolding (Doyle et al. 2007; Mogk et al. 2015; Schaupp et al. 2007; Weber-Ban et al. 1999).
5
Hsp70 is one of the most conserved chaperones present in all major cellular compartments and participates in divers cellular processes ranging from protein folding to protein translocation across the membrane (Clerico et al. 2015; Mayer and Bukau 2005). It is involved in de novo protein folding of nascent polypeptides in the ribosome and prevents their premature folding in physiological condition. It promotes translocation of newly synthesized proteins from cytosol into the cell organelles (Mayer and Bukau 2005). Structurally, it consists of ATPase and protein binding domains (Zhu et al. 1996). It binds with high affinity to the unstructured segment of polypeptides in the ADP post-hydrolysis state. Hsp40/DnaJ binds to the nonnative proteins to deliver them to Hsp70 (Kampinga and Craig 2010; Mayer et al. 1999). Indeed, J-protein and nucleotide exchange factor are two co-chaperones of Hsp70 (Craig and Marszalek 2017). Indeed, J-domain simulates ATP-hydrolysis by interacting with ATPase domain of Hsp70 (De Los Rios et al. 2006; Goloubinoff and De Los Rios 2007; Kampinga and Craig 2010).
1.2 Small heat shock proteins (sHsps)
Small heat shock proteins (sHsps) are a heterogeneous family of molecular chaperones and are among the first proteins expressed during stress response (Richter et al. 2010; Tissieres et al. 1974). They accumulate under stress conditions as well as ageing and are present in all three kingdoms of organisms except for some pathogenic bacteria (Bakthisaran et al. 2015; Basha et al. 2013; Caspers et al. 1995; de Jong et al. 1993; Franck et al. 2004; Haslbeck et al. 2005; Maaroufi and Tanguay 2013; Mymrikov and Haslbeck 2015; Narberhaus 2002; Plesofsky-Vig et al. 1992; Walther et al. 2015; Waters 1995; Waters and Vierling 1999b). The most studied members of this family are αA-crystallin (HspB4) and αß-crystallin (HspB5) in human (Haslbeck et al. 2016; Horwitz 2003), which work together with γ-crystallin to protect eye lens proteins from formation of irreversible aggregates (Bakthisaran et al. 2015; Mymrikov and Haslbeck 2015). The number of encoding genes varies in different organisms and is higher in higher eukaryotes (Haley et al. 2000; Haslbeck et al. 2005; Kriehuber et al. 2010). For example, the number of genes vary from one to twelve in bacteria, ten or more in human and murine genomes, fourteen in
Caenorhabditis elegans, twelve in Drosophila melanogaster and up to nineteen in higher
et al. 2010; Waters 2013). They are unique according to their level and time of expression and in their cellular localization which represent their adaptations based on their cellular and environment requirements (Morrow and Tanguay 2015b; Tanguay and Hightower 2015). They are ATP-independent holdases with huge ability to bind to the unfolded substrates and prevent them from irreversible aggregation. The clients proteins would then be released and become refolded with the help of other ATP-dependent chaperones at the normal condition or become degraded through the PQC machine (Basha et al. 2013; Basha et al. 2012; Kriehuber et al. 2010; Lee et al. 1997; Mogk et al. 2003). Up to now plant ,
Drosophila melanogaster and Toxoplasma gondii are unique compared to other eukaryotes
due to having of organelle-localized sHsps (de Miguel et al. 2005; Michaud et al. 2002; Morrow et al. 2000; Siddique et al. 2008; Waters 2013; Waters and Rioflorido 2007; Waters and Vierling 1999a). Small Hsps have some common characteristics such as low subunit molecular mass ranging between 12 to 43 kDa (Basha et al. 2013; Basha et al. 2012; Bepperling et al. 2012; de Jong et al. 1993; Delbecq and Klevit 2013). Structurally, they can exist as monomers and dimers but also form large dynamic oligomers typically composed of 24-40 subunits in which reversible dissociation of their subunits is important for their chaperone function at high temperature (Jaya et al. 2009; McDonald et al. 2012; Stengel et al. 2010; van Montfort et al. 2001b). Their expression is increased in response to the stress conditions and they have chaperone like activity (Basha et al. 2012; Delbecq and Klevit 2013; Hanazono et al. 2013; Haslbeck et al. 2005; Hilton et al. 2013; Kim et al. 1998). They form large oligomeric structure depending on the transient interactions between different regions of their structures (Basha et al. 2012; Haslbeck and Vierling 2015; Hilton et al. 2013). Small Hsps are key components of normal cells and any failure in their expression and/or their mutations has been associated with diseases such as myopathies, cataracts and neurodegenerative diseases (Sun and MacRae 2005). Their functions are diverse and include thermotolerance, cell differentiation, establishment of membrane, apoptosis and modulation of cytoskeletal dynamics (Arrigo and Gibert 2014; Haslbeck et al. 2016; Nakamoto and Vigh 2007).
7
Small Hsps are characterized by presence of a highly conserved middle region, called αlpha-crystallin domain (ACD) as their hallmark (Finn et al. 2014; Horwitz 1992; Kappe et al. 2010; Kriehuber et al. 2010; van Montfort et al. 2001b). ACD is flanked with less conserved and diverse N-terminal domain (NTD) in one side and short C-terminal domain (CTD) in the other side (de Jong et al. 1998; de Jong et al. 1993; Haslbeck et al. 2004; Kriehuber et al. 2010). Investigations have demonstrated that all three domains play an essential role in structure and function of sHsps (Mainz et al. 2015; McDonald et al. 2012; Van Montfort et al. 2001a). Analysis of more than 8,700 sHsp sequences has demonstrated that their structure is composed of on average 161 amino acids (Kriehuber et al. 2010). The length of ACD is well preserved through the evolution with an average 90-100 residues, which is the most bulk part of sHsps structure (approximately 58 %). NTD and CTD with an average 56 and 10 residues respectively form approximatively 35 and 6 % of sHsp structure (Hilton et al. 2013; Kriehuber et al. 2010). NTD and CTD represent the large variations in their length. Archeas and bacteria possess smaller length, while eukaryotes represent the longer sequence (Kriehuber et al. 2010).
The ACD is characterized by presence of charged amino acids (Kriehuber et al. 2010; Poulain et al. 2010). Expressed recombinant ACDs form dimers, but are not able to form higher-order oligomers (Bagneris et al. 2009; Feil et al. 2001; Kim et al. 1998). There are highly conserved notable positions at this domain in which mutations result in a variety of pathological conditions (Benndorf and Welsh 2004). Example of this is “Arginine disease” (R120G in HspB5) (Poulain et al. 2010).
CTD is composed of two structural parts known as tail and extension (CTE) usually separated by highly conserved sequence of IXI (Carver 1999; Delbecq and Klevit 2013; Taylor and Benjamin 2005; van Montfort et al. 2001b). IXI motif is critical for oligomer assembly of some sHsps (Delbecq et al. 2012; Delbecq and Klevit 2013; McHaourab et al. 2012). Loss of IXI motif resulted in disassembly of oligomeric structures in some of the bacteria, mammalian or plant sHsps (Kakkar et al. 2014; McHaourab et al. 2012; Quinlan et al. 2013; Shi et al. 2006; Studer et al. 2002; Zhou et al. 2016). Palindromic nature of IXI motif allows it to bind to the two different edges of ACD known as ß4/ ß8 groove in crystal structure of many identified homo-oligomers such as HspB5, MjHsp16.5 and wheat
Hsp16.9 (Delbecq and Klevit 2013; Kim et al. 1998; Laganowsky et al. 2010; van Montfort et al. 2001b). Highly conserved IXI motive acts as “anchor” to make oligomers and induce polydispersity of ACD (Delbecq and Klevit 2013). Mutation or deletion in IXI motif is deleterious not only for oligomerization but also for their chaperone like activity (Saji et al. 2008). In Hsp18.5 from Arabidopsis thaliana this motif is not required to form oligomeric structure and AtHsp18.5 exists as stable dimer (Basha et al. 2013).
CTD is polar, flexible and accessible for the solvent more due to extreme C-terminus for the residues located downstream of the IXI motif for example residues E164-S173 in HspB4 and residues E164-K175 in HspB5 which induce more flexibility as seen in NMR data (Carver et al. 1992; Carver and Lindner 1998; Jehle et al. 2011; Merck et al. 1993; Peschek et al. 2013; Treweek et al. 2010). Hydrophobic behavior of CTD induces solubility of oligomers especially via CTE. In this regard, substitution of polar residues of CTD by hydrophobic amino acids result in a decrease in flexibility, thermostability and diminishes its functional activity (Smulders et al. 1996). Recently, the importance of CTE in oligomerization, subunit exchange and chaperone function of Hsp16.3 in Mycobacterium
tuberculosis has been reported (Panda et al. 2017).
NTD possesses huge diversity in length and amino acid composition and with almost no conservation (Haslbeck et al. 2016). Variation in this segment is responsible for different oligomer size in sHsps (Haley et al. 2000). Investigations showed significant role of NTD in chaperone activity (Basha et al. 2013). Aromatic amino acids such as tryptophan and phenylalanine (Kriehuber et al. 2010) are located in this domain. Truncated N- terminal leads to the formation of oligomers which are obviously smaller in size compared to the full length protein (Chiou et al. 2010; Hilton et al. 2013; Ito et al. 1997; Kamei et al. 2000; Kato et al. 1998; Kundu et al. 2007; Lund et al. 1996; MacCoss et al. 2002; Merck et al. 1993; Miesbauer et al. 1994; Peschek et al. 2013; Renkawek et al. 1992). Nuclear localization sequence (NLS) of DmHsp27 (Michaud et al. 2008), mitochondrial localization sequence (MLS) in DmHsp22 and Toxoplasma gondii are located at this segment (de Miguel et al. 2005; Morrow et al. 2000). NTD is involved in formation of highly ordered oligomers in addition to responsibility for binding to the non-native client proteins in many sHsps (Fu et al. 2005; Giese and Vierling 2004; Jiao et al. 2005). NTD is rich in hydrophobic residues
9
which are buried in the oligomers at the normal conditions and exposed in response to elevated temperature while dissociate the oligomeric structure. This could trigger non-native substrate binding (Narberhaus 2002). Deletion of the full region of N-terminal in DmHsp27 results in total loss of chaperone-like activity and deletion of the FGFG sequence at the position 29-32 of DmHsp27 as well as the single mutations at these residues enhanced oligomers and chaperone activity which suggest the importance of this sequence in structure and function of Drosophila melanogaster (Moutaoufik et al. 2017).
In mammalian, NTR is considered to be main site for conformational changes and regulation of ACD due to the presence of phosphorylation’s sites and post-translation modifications such as oxidation, deamination and glycation (Chiou et al. 2010; Garrido et al. 2012; Ito et al. 1997; Kamei et al. 2000; Kriehuber et al. 2010; Lund et al. 1996; MacCoss et al. 2002; Miesbauer et al. 1994; Smith et al. 1992). Phosphorylation regulates the rate of association/dissociation of subunits as well as chaperone like activity. HspB1, HspB5, HspB6 and HspB8 in human are among the one in which phosphorylation was reported (Beall et al. 1999; Cantin et al. 2008; Dreiza et al. 2010; Ecroyd et al. 2007; Kato et al. 1998; Kostenko and Moens 2009; Rogalla et al. 1999; Villen et al. 2007). For example, phosphorylation of HspB5 on ser-59 and ser-45 leads to its translocation from cytoplasm to the nuclear speckles (Adhikari et al. 2004; den Engelsman et al. 2013). Furthermore, phosphorylation of Hsp27 and HspB5 in response to heat stress trigger formation of smaller oligomers that are less active in terms of chaperone-like activity (Mellier et al. 2013). In addition these phosphorylated structures are more prone to substrate binding due to the higher flexibility and accessibility of this part (Aquilina et al. 2004; Ito et al. 1997; Kato et al. 1998). This diversity in NTD possibly makes this region a key site for client’s identification (Lund et al. 1996; MacCoss et al. 2002; Miesbauer et al. 1994; Voorter et al. 1986). Recent study has been shown, NTD not only is necessary for in
vitro chaperone-like activity but also is essential and correlated with in vivo
thermotolerance (Cha et al. 2016). Studies on chloroplast-localized Hsp21 suggest the involvement of NTD and CTD in the oligomer stabilization and clients interactions (Ahrman et al. 2007a; Ahrman et al. 2007b; Lambert et al. 2011).
Figure 1.1 Primary structure of sHsps
ACD (dark blue); the most conserved region in the primary structure of sHsps is flanked with the non-conserved NTD (orange) and CTD (red). Conserved IXI motive (dark green), tail and extension (light green) have been located at CTD. Phosphorylation sites (gray) are located at the NTD of some of the sHsps.
1.2.2 Dynamic behavior of sHsps
Dynamic behavior of sHsps has been reported by Van Montfort in 2002 for the first time and later supported for many other sHsps using Florescence resonance energy transfer (FRET) and mass spectroscopy (MS) (Baldwin et al. 2011a; Baldwin et al. 2011c; Painter et al. 2008; Stengel et al. 2010). Small Hsps are dynamic in their secondary, tertiary and quaternary structures. They continually dissociate and re-associate with both their sub-oligomeric structures and client proteins (Baldwin et al. 2011a; Painter et al. 2008; Stengel et al. 2010; Van Montfort et al. 2001a). Dynamic feature of sHsps prevents achievement of a high-resolution oligomeric structure except recently reported information related to the ACD and dimer of some sHsps such as HspB5 (Delbecq and Klevit 2013). Subunit exchange is a rapid process and its rate is a significant factor determining the substrate binding (Delbecq et al. 2012). Small Hsps are able to exchange their subunits even under physiological conditions due to the dissociation/re-association of their oligomers (Bova et al. 1997; Bova et al. 2002; van Montfort et al. 2001b). Indeed, oligomerization and dissociation is pre-requisite for exposure of hydrophobic substrate binding sites, affect their binding affinity to the clients and consequently influence their chaperone functions (Delbecq and Klevit 2013; Delbecq et al. 2015; Haslbeck et al. 1999; Vanhoudt et al.
11
2000).
Rapid subunit exchange occurs for both polydisperse metazoan proteins as well as well-defined monodisperse Hsps such as wheat Hsp16.9 (Basha et al. 2012; Schneider et al. 2001; Uetrecht et al. 2010). Small Hsps increase their structural rearrangement in response to the elevated temperature (Basha et al. 2012; Sobott et al. 2002; Van Montfort et al. 2001a) and this varies in different sHsps (Basha et al. 2012; Van Montfort et al. 2001a). In wheat Hsp16.9 rearrangement appears to lead to dissociation of oligomer into the substrate-bound dimers, which are active heat shock binding forms. Oligomer dissociation causes NTD and CTE exposure leading to stabilized oligomeric structure (Basha et al. 2010; Stromer et al. 2004; Van Montfort et al. 2001a). In this case hydrogen exchange take places in less than 5 seconds which proves flexibility, dynamic and disordered character of NTD and support its role in substrate binding (Basha et al. 2006; Cheng et al. 2008; Patel et al. 2014).
Isolated ACD forms stable dimers but not oligomers (Bagneris et al. 2009; Baranova et al. 2011; Clark et al. 2011; Feil et al. 2001; Laganowsky et al. 2010). Crystal structure of Hsp16.5 from Methanocaldcoccus janaschii (24-mer) and Hsp16.9 from Triticum aestivum
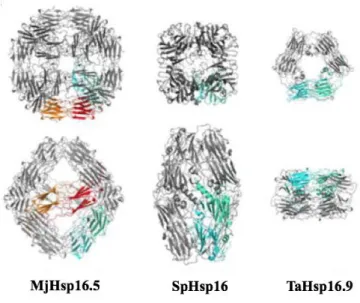
(12-mer) showed that ACD is a compact ß-sheet sandwich structure composed of eight IgG (immunoglobulin) -like ß-strand (ß2- ß9) which sets up in two anti-parallel layers of three (ß3- ß8- ß9) and four (ß4- ß5- ß6+7) ß-strands connected by a short inter-domain loop (Fig 1. 2) (Delbecq and Klevit 2013; Haslbeck et al. 2016; Kim et al. 1998; Van Montfort et al. 2001a; van Montfort et al. 2001b). Well-identified crystal structure of MJ Hsp16.5 and wheat Hsp16.9 have been obtained by X-ray (Kim et al. 1998; van Montfort et al. 2001b).
The mode of dimerization is different between metazoan and non-metazoan and presence of PG motif (conserved ProGly) in non-metazoan and KH (conserved LysHis) motif in metazoan supports the differences (Clark et al. 2011; Haslbeck and Vierling 2015). Dimeric structure in non-metazoan including plant, archaea and bacteria occur via inter-subunit ionic interaction between one conserved arginine residue located in the ß6-strand from one monomer to the aspartate or glutamate residue of ß6-strand of the partner dimer subunit (ß6-swapped dimer) (Bepperling et al. 2012; Clark et al. 2011; Hanazono et al.
2013; Hilario et al. 2011; Kim et al. 1998; Takeda et al. 2011; van Montfort et al. 2001b). Indeed, in non-metazoan, reciprocal swapping of ß6 strand from one monomer into the ß6 strand of the neighboring monomer leads to the dimer formation (Bepperling et al. 2012; Hanazono et al. 2013; Kim et al. 1998; Van Montfort et al. 2001a). Arabidopsis thaliana
Hsp18.5 lacks ß6, which is involved in dimerization of other plant and bacterial sHsps (Basha et al. 2013). It forms stable dimer rather than oligomer and its large oligomeric structure is more due to the substrates-sHsp interactions and is not prerequisite for its substrate binding. In contrast, lack of ß6 strand in metazoan like vertebrate leads to the extended ß7 which fused with ß6 strand and form “ß6+7” to interact with its equivalent ß strand from the neighboring monomer in an anti-parallel orientation (ß7-interface dimer) (Fig 1.2) (Bagneris et al. 2009; Baranova et al. 2011; Berengian et al. 1999; Delbecq and Klevit 2013; Jehle et al. 2010; Jehle et al. 2009; Koteiche and McHaourab 1999; Laganowsky et al. 2010).
X-ray crystallography registered three identified AP (antiparallel) interface (AP1, AP2 and AP3) for vertebrate based on the dimeric interaction between ß6+7 strands (Bagneris et al. 2009; Clark et al. 2011; Laganowsky et al. 2010). This nomination is mostly dependent on the decreasing overlapping sequence between ß6+7 strands in AP interface (Laganowsky et al. 2010). NMR together with hydrogen/deuterium exchange on full length of human HspB5 (Jehle et al. 2010) showed the very dynamic behavior of surface interface in HspB5 (Hasan et al. 2004; Jehle et al. 2010).
13
β6-swapped dimer of the ADC of M. jannaschii Hsp16.5 (X-ray crystallography, PDB: 1SHS) (Kim et al. 1998). ACD of each subunit is identified in green and gray (A). β7-interface of the ACD of human HspB5 (solid state NMR, PDB: 2KLR) (Jehle et al. 2011). ACD of each subunit is identified in green and gray (B). (Haslbeck and Vierling 2015).
Most sHsps tend to form large, usually polydisperse polymers compromising of 12-32 or even more subunits (Delbecq and Klevit 2013; Haslbeck and Vierling 2015). Structural information on sHsps are usually due to the truncated forms as it is hard to have high-resolution structure due to their polydispersity (Hilton et al. 2013; Van Montfort et al. 2001a; van Montfort et al. 2001b). Despite tendency of sHsps to form oligomers, some of the sHsps only present in the smaller structure (Basha et al. 2012) among which are HspB6 (Hsp20) and HspB8 (Hsp22) with dimeric or monomeric structure (Stamler et al. 2005). Tsp36 a sHsp from Taenia saginata forms dimeric and tetrameric structures(Kappe et al. 2004), Arabidopsis thaliana Hsp18.5 (Basha et al. 2013) and Deinococcus radiodurans Hsp17.7 (Bepperling et al. 2012) form dimers.
Dimers assemble together to form oligomers through interaction between different parts of sHsps (Braun et al. 2011; Delbecq and Klevit 2013). Formation of dimer is the first level of hierarchy in the sHsps structure (Delbecq and Klevit 2013). I-X-I/V motif from one dimer interacts with hydrophobic groove formed by ß4 and ß8 strands of neighboring dimer to form tetramer or hexamer, which is the second level of hierarchy (Braun et al. 2011; Fleckenstein et al. 2015). NTD is involved in formation of higher oligomeric structure of sHsps leading to a third level of hierarchy. Removing NTD leads to the formation of smaller oligomers (Haslbeck and Vierling 2015; Moutaoufik et al. 2017). Small Hsps are polydisperse and form population of oligomers containing different number of subunits, which are dynamic and constantly exchange their subunits (Braun et al. 2011; Tanguay and Hightower 2015). The degree of heterogeneity (presence of different oligomeric population) in different family of sHsps is not similar and reflect their chaperone like activity (Basha et al. 2012).
Figure 1.3 Comparison of the three available oligomeric structure of sHsps
M. jannaschii Hsp16.5 representing a 24mer (Kim et al. 1998). Schizosaccharomyces
pombe Hsp16 representing a 16mer (Hanazono et al. 2013). Tritium aestivum (wheat)
Hsp16.9 representing a 12mer (Van Montfort et al. 2001a). Dimeric building block presented in green-cyan to shows interconnections of dimers. The orange-red dimeric building block representing an octahedron (Haslbeck and Vierling 2015).
1.2.3 Chaperone activity and substrate recognition by sHsps
Small Hsps are molecular chaperones with the ability to recognize, bind and stabilize a wide range of denatured proteins and prevent them from irreversible aggregation in an ATP-independent form (Basha et al. 2013; Haslbeck et al. 1999; Horwitz 1992; Ingolia and Craig 1982; Jakob et al. 1993; McHaourab et al. 2002; Morrow et al. 2006; Ratajczak et al. 2009). This characteristic was first discovered for α-crystallin in the early 1990 (Horwitz et al. 1992; Jakob et al. 1993). Their ability to bind a plethora of non-native proteins have been shown in many studies against different substrates (Haslbeck et al. 1999; Horwitz et al. 1992; Studer and Narberhaus 2000). They could form stable complexes with the unfolded substrates (Haslbeck et al. 1999; Stengel et al. 2010) in which substrates are in the potential state to be refolded immediately after removing the stress condition (Ehrnsperger et al. 1997; Kampinga et al. 1994; Lee et al. 1997). Indeed, sHsps
15
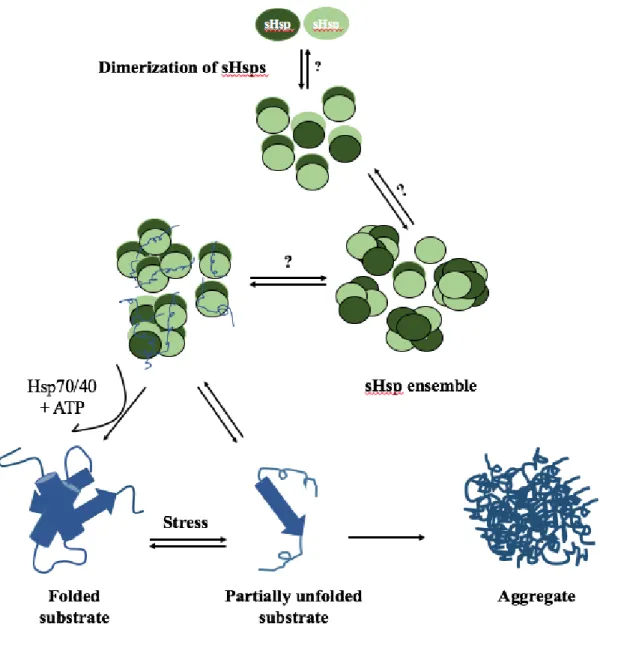
they become aggregated. However, they cannot rescue already aggregated substrates. Nevertheless, some of the unfolded intermediate structures separate from sHsps at the beginning of unfolding process and spontaneously become refolded (Bepperling et al. 2012; Haslbeck and Vierling 2015; McHaourab et al. 2002). Indeed, identity of substrates, degree of substrate’s unfolding and specific characteristic of sHsps determine the stability of the interaction between sHsps and substrates while some of the interactions are irreversible (Basha et al. 2012; Garrido et al. 2012; Haslbeck et al. 2005; Horwitz et al. 1992; McHaourab et al. 2009; Stengel et al. 2010). Electron microscopy (Basha et al. 2004; Haslbeck et al. 1999; Stromer et al. 2003) and mass spectrometry (Stengel et al. 2010) results revealed that sHsp-substrate complexes are larger and more soluble than the sHsp oligomers alone. Some studies suggest that sHsp-substrate complexes are dissociated forms of sHsps, usually dimers, and re-associated into another form oligomer (Haslbeck and Vierling 2015). Small Hsps work as a buffer system to protect unfolded proteins from irreversible aggregation during stress (Eyles and Gierasch 2010) (Figure 1.4).
In vitro investigations have revealed that unfolded proteins trapped in complexes with
sHsps release and refold in cooperation with ATP-dependent chaperone machinery including DnaK-DnaJ-GrpE in bacteria and Hsp70/Hsp40 system in mammalian and plants (Figure 1.4) (Ehrnsperger et al. 1997; Finka et al. 2011; Kampinga et al. 1994; Lee and Vierling 2000; Liberek et al. 2008; Ratajczak et al. 2009; Stege et al. 1994). In HspB5 release of substrates is dependent to the Hsp70/Hsp40 as well as phosphorylation (Peschek et al. 2013). Ratio of sHsp to substrate is important as ATP-dependent chaperone system is only efficient in the presence of excess concentration of sHsps to the substrates. In E. coli, presence of a protein disaggregase, ClpB, is an essential factor for refolding process (Bepperling et al. 2012; Cashikar et al. 2005; Haslbeck et al. 2005; Mogk et al. 2003; Ratajczak et al. 2009).
Although, in vivo potential substrates of sHsps in stress condition are poorly understood, proteomic approaches identified preferably interactions of some cytoplasmic proteins with sHsps. Among them are translation-related proteins including ribosomal proteins, amino-acyl tRNA synthases and some metabolic enzymes (Bepperling et al. 2012; Fu 2014; Fu et al. 2013). Proteomic approaches identified 127 proteins out of 17,225 as the
potential interactors of HspB4 (Fan et al. 2014). They are mostly related to the metabolic processes, protein transportation, localization and cell cycle (Fan et al. 2014).
Small Hsps are more effective to protect smaller substrates compared to larger ones
which supports the importance of mass ratio in this process (Basha et al. 2012). While substrate binding sites of sHsps remain to be identified, investigations suggest that multiple recognition sites work together to bind different substrates. Molecular dynamics experiments on NTD of dimer Hsp16.9 from wheat (1GME) and its homologous Hsp18.1 from pea show availability of hydrophobic surfaces on the NTD of sHsp to interact with hydrophobic regions of substrates supporting involvement of NTD for substrate recognition and chaperone activity (Patel et al. 2014). Other investigations suggest possible involvement of ACD and CTE in substrate recognitions (Cheng et al. 2008; Ghosh et al. 2007; Jaya et al. 2009). Experiments showing ACD alone possesses chaperone activity support this idea (Jehle et al. 2009; Laganowsky et al. 2010). Mutation of a very conserved G residue in the HspH in Bradyrhizobium japonicum reduced chaperone activity without affecting other properties showing involvement of ACD in this process (Hochberg et al. 2014; Lentze et al. 2004; Lentze and Narberhaus 2004). Mutations in amino acids located at the CTE of HspB5 influence chaperone activity as well as structure of sHsps (Treweek et al. 2007; Treweek et al. 2010). Hydrophobic grooves between ß4 and ß8 strands at ACD are potential binding sites due to their exposure under stress condition (Laganowsky et al. 2010; Treweek et al. 2007; van Montfort et al. 2001b). However, cross-linking experiments detected less accessible (<300 Aº) hydrophobic surfaces on ß4 -ß8 for substrate binding at ACD compared to the NTD (Ahrman et al. 2007b; Jaya et al. 2009; Patel et al. 2014). Accumulation of substrate binding sites at variable and non-conserved sites outside the ACD, may be an explanation for variation in substrate specificity of different sHsps (Kriehuber et al. 2010). Few investigations regarding to sHsp-binding sites on different substrates indicated that only small part of substrate’s sequence bound to the sHsp which supports the idea that only early exposed motif on the early unfolded intermediate structure of substrates binds to the sHsps (Ahrman et al. 2007a; Ahrman et al. 2007b; Cheng et al. 2008). Experiments to examine the structure of bound malate dehydrogenase to two
17
different sHsps support interaction with the early unfolded intermediate structure (Cheng et al. 2008).
The effect of oligomeric structure of sHsps on the chaperone activity is not always the same. Usually oligomeric sHsps are strong chaperones although HspB6, HspB8 and AtHsp18.5 with dimeric form possess robust chaperone-like activity (Basha et al. 2013; Bukach et al. 2004; Stamler et al. 2005). Tsp36 a sHsp from Taenia saginata is composed of dimeric and tetrameric structures with strong chaperone activity against insulin and CS (Stamler et al. 2005). Human HspB1, HspB4 and HspB5 are the strongest chaperones and HspB7 is not an efficient chaperone (Vos et al. 2010); this might be due to its unusual NTD (Huntley and Golding 2006). HspB5, HspB6 and HspB8 are able to decrease toxicity and formation of Aß fibril (Wilhelmus et al. 2006a; Wilhelmus et al. 2006b). HspB1 prevents formation of harmful and toxic tau (Shimura et al. 2004) and HspB8 is able to prevent in
vivo aggregation of poly Q containing proteins (Carra et al. 2005). Hsp26 and Hsp42 in
yeast dissociate to smaller active structure in response to the heat, which efficiently bind to the aggregation prone proteins and form large globular assembly (Haslbeck et al. 2005; Haslbeck et al. 1999).
Figure 1.4 Model for chaperone function of Shsps
Upon stress condition, proteins become partially unfolded and destabilized. Small Hsps can recognize and bind to denatured substrates in the ATP-independent manner and maintain them in the folding-competent state. Large sHsp complexes become active and form smaller structure probably dimer that can stabilize denatured substrates. These substrates can be captured by ATP-dependent chaperones and be refolded or either become aggregate and form insoluble complexes Model adapted from (Haslbeck and Vierling 2015).