Répartition et viabilité d'une population vulnérable de
bisons des prairies
Thèse
Ricardo Simon
Doctorat en biologie
Philosophiæ doctor (Ph. D.)
Résumé
L’identification des facteurs qui déterminent les patrons de répartition et d’abondance des animaux est un sujet de recherche actuel et majeur en écologie. L’objectif général de ma thèse était de mieux comprendre comment un facteur en particulier, la prédation – prise au sens large pour inclure la chasse par les humains – influence les liens entre l’utilisation de l’espace et la dynamique des populations. Ma thèse explore les liens entre le risque de prédation, les déplacements et la sélection de l’habitat, le taux de mortalité et la viabilité d’une population vulnérable d’un grand herbivore. Le modèle d’étude a été la population de bisons des prairies (Bison bison bison) du parc national de Prince Albert, en Saskatchewan, soumise notamment à la prédation par le loup gris (Canis lupus) et à la chasse par les humains. Dans le chapitre 1, je mets en lumière les stratégies d’utilisation de l’espace des bisons face aux loups, et vice-versa, à l’aide d’analyses de déplacements et de sélection de l’habitat. De la deuxième moitié de l’été au début de l’hiver, les bisons réduisaient le temps passé sur des parcelles riches en nourriture suivant l’augmentation du niveau de fréquentation à long terme de ces parcelles par les loups. Les bisons quittaient également ces parcelles rapidement lorsqu’un loup était à proximité. En hiver, cependant, les bisons ne semblaient réagir qu’en s’éloignant lorsqu’un loup était à proximité. L’absence de réaction par les bisons au risque à long terme de rencontrer un loup en hiver pourrait s’expliquer sur une base énergétique : la nourriture est alors moins digestible et la neige rend leurs déplacements plus coûteux. Bien que le risque de prédation influence l’utilisation de l’espace des bisons, je démontre dans le chapitre 2 que la prédation par les loups, ainsi que des épisodes stochastiques de maladie (anthrax), ne représentent pas actuellement des dangers pour la viabilité de la population. À l’inverse, sous les conditions actuelles, la chasse par les autochtones – légale mais non réglementée – est la raison principale pour laquelle le risque d’extinction de la population atteint 66% sur les 50 prochaines années. Les bisons sont vulnérables à la chasse lorsqu’ils sortent du parc pour se nourrir sur des champs agricoles, attirés par une riche nourriture. Mes analyses précisent que chaque 1% de temps additionnel passé sur des champs avec permission de chasse de
2011 à 2016 augmentait le risque de mortalité par la chasse de 9%. Je révèle en outre que le temps passé sur ces champs devrait chuter de 70% pour que l’effectif actuel de la population se maintienne dans le temps suivant un scénario où la population est encore soumise à la prédation par les loups et à des épisodes d’anthrax. L’utilisation de seulement cinq champs agricoles comptait déjà pour plus de 70% de l’utilisation totale des champs avec permission de chasse par les bisons. Mettre en place des plans de gestion qui ciblent davantage ces cinq champs serait une stratégie efficace à court terme pour freiner le déclin de la population. Bien qu’une telle stratégie puisse mener les bisons à augmenter leur utilisation d’autres champs, l’impact démographique de la chasse devrait diminuer en conséquence, au moins à court terme, car cette activité est interdite dans la plupart des autres champs utilisés régulièrement par les bisons. Enfin, dans le chapitre 3, je compare, à l’aide d’un modèle basé sur l’individu, l’efficacité de différentes interventions de gestion manipulant la profitabilité (c.-à-d., le ratio entre quantité d’énergie digestible et temps de consommation) et la répartition de la nourriture pour réduire le temps que les bisons passent à l’extérieur du parc. Mes simulations suggèrent qu’assécher des prés à l’intérieur du parc pour y augmenter la disponibilité en nourriture naturelle serait peu efficace. Cependant, mes simulations suggèrent également que la mise en culture, à l’extérieur du parc, d’espèces végétales moins profitables pour les bisons que la nourriture disponible dans le parc serait une meilleure intervention. Cette thèse révèle d’abord le caractère dynamique et complexe des stratégies anti-prédatrices d’utilisation de l’espace d’un grand herbivore dans un système multi-proies. Ce travail met ensuite en lumière l’utilité pratique des différentes approches reliant l’utilisation de l’espace et la viabilité d’une population afin d’orienter la mise en place d’interventions de gestion plus efficaces. Le résultat final est une étude de cas approfondie visant à améliorer notre capacité à sauvegarder à court terme des populations vulnérables à des menaces reparties de façon hétérogène dans l’espace.
Abstract
Determining the factors that shape patterns of animal distribution and abundance is a major topic in contemporary ecological research. The overarching objective of my thesis was to better understand how one such factor, predation – in its broader meaning to include harvesting by humans – influences the links between space use and population dynamics. My work explores the links between predation risk, movement and habitat selection, mortality rates and the viability of a threatened population of large herbivore. The study system was the plains bison (Bison bison bison) population of Prince Albert National Park, Saskatchewan, subject most notably to predation by grey wolves (Canis lupus) and harvest by humans. In chapter 1, I use movement and habitat selection analyses to reveal the strategies of space use deployed by bison in response to wolves and vice-versa. From mid-summer to the onset of winter, bison reduced the time spent in patches rich in food as the long-term risk of encountering wolves there increased. Bison also left these patches more quickly when wolves were relatively close by. In winter, however, bison were observed to only react by moving away from nearby wolves. The absence of a bison response to the long-term risk of encountering wolves in winter could be explained by energetic constraints, as food is less digestible and movement more costly due to snow cover during this season. Although I reveal that perceived predation risk influences bison use of space, I show in chapter 2 that neither wolf predation nor disease (in the form of anthrax outbreaks) represent a threat to the viability of the population. Rather, the main reason behind a probability of population extinction of 66% over the next 50 years under current conditions is the legal, yet unregulated, harvest by native hunters. Bison are vulnerable to harvest when they leave the park to forage on rich food available in agricultural fields. My analyses refine our understanding of this pattern by showing that every additional 1% of time spent in fields with hunting permission from 2011 to 2016 increased the risk of harvest mortality by 9%. I also reveal that the time bison spend in such fields must drop by 70% for population abundance to remain stable at its current level in a scenario of continued wolf predation and anthrax outbreaks. More than 70% of bison use of fields with hunting permission were
limited to just five fields. Management interventions targeting these riskier fields would be an effective short-term strategy to halt the population’s decline. Even though such an approach might lead bison to increase their use of other fields, the demographic impact of harvesting should consequently diminish, at least over the short term, given that harvesting is not permitted in most other fields used by bison. Finally, in chapter 3, I use an individual-based model to compare the relative effectiveness of different management interventions manipulating food profitability (i.e. the ratio between digestible energy and handling time) and distribution to reduce the time bison spend outside the park raiding crops and, thereby, the number of individuals harvested. My simulations suggest that draining meadows inside the park to increase the availability of natural forage there would not be very effective. However, my simulations also suggest that cultivating crops outside the park of lower profitability relative to natural forage inside the park would be a better intervention. My thesis reveals the dynamic and complex nature of the anti-predator movement and habitat selection strategies deployed by a large herbivore in a multi-prey system. My work also highlights the practical interest of linking spatial distribution to population viability to lead to more effective management interventions. The overall result is a thorough case study aimed at improving our ability, over the short term, to conserve populations vulnerable to threats which are distributed heterogeneously in space.
Table des matières
Résumé ... ii
Abstract ... iv
Table des matières ... vi
Liste des figures ... xi
Liste des tableaux ... xvii
Remerciements ... xxii
Avant-propos ... xxiv
Introduction générale ... 1
1. L’écologie spatiale pour relier répartition spatiale et viabilité : cadre théorique et exemples empiriques ... 1
2. Le rôle de la prédation dans l’abondance et la répartition des proies ... 5
3. Le jeu spatio-temporel prédateurs-proies ... 10
4. L’hétérogénéité spatiale de l’habitat et la prédation ... 14
5. Homo sapiens en tant que prédateur ... 15
6. L’écologie spatiale dans la gestion et la conservation de la faune ... 16
7. Objectif et organisation de la thèse ... 19
8. Modèle d’étude : les bisons des prairies du parc national de Prince Albert ... 21
Chapitre 1. Complex tactics in a dynamic large herbivore-carnivore spatiotemporal game ...25
1.1 Résumé ... 26
1.2 Abstract ... 27
1.3. Introduction ... 28
1.4. Materials and Methods ... 31
1.4.2. Ecological seasons ... 32
1.4.3. Statistical analysis 1: seasonal resource matching to test the Leap-Frog hypothesis ... 33
1.4.4. Statistical analysis 2: player responses to opponent to test the Risky Places, Risky Times and Shell Game hypotheses ... 34
1.4.5. Residency time in meadows and direction of movement with respect to opponent ... 36
1.4.6. Long-term use of space by the opponent ... 36
1.4.7. Index of opponent proximity ... 37
1.5. Results ... 38
1.6. Discussion ... 40
1.7. Supplementary Material ... 50
1.7.1. Appendix 1 – Distribution of bison and wolf GPS collar data across years and seasons ... 50
1.7.2. Appendix 2 – Variance inflation factors ... 51
1.7.3. Appendix 3 – Direction of bison movement with respect to wolf position in spring-summer ... 53
1.7.4. Appendix 4 – Direction of bison movement with respect to wolf position in winter in Prince Albert national park from 2007 to 2016 ... 54
Chapitre 2. Linking habitat use to mortality and population viability to disarm an ecological trap ...55
2.1 Résumé ... 56
2.2 Abstract ... 57
2.3. Introduction ... 58
2.4.3. Photography data: tracking adult females through time ... 63
2.4.4. Capture-mark-recapture (CMR): estimating abundance and survival probability of adult females ... 64
2.4.5. Stage ratios: estimating the abundance of female calves and juveniles ... 65
2.4.6. Harvest data: minimum estimate of the proportion of mortality due to harvest ... 65
2.4.7. Stochastic matrix population model: projecting current conditions into the future ... 66
2.4.8. Cox proportional hazards model: linking adult female survival to the intensity of use of agricultural fields with hunting permission ... 68
2.4.9. Spatially explicit population viability analysis: linking the intensity of use of fields with hunting permission to population persistence ... 69
2.5. Results ... 69
2.6. Discussion ... 71
2.7. Supplementary Material ... 80
2.7.1. Appendix 1 – Description of the method used to identify adult females through time, including quantification and correction of measurer bias, false-rejection and true-false-rejection rates ... 80
2.7.2. Appendix 2 – Calf- and juvenile-adult female empirical ratios and harvest data ... 83
2.7.3. Appendix 3 – Stochastic projections of the abundance of female bison in Prince Albert national park and adjoining lands, Canada, for next 50 years under parameters yielding a stable population ... 84
2.7.4. Appendix 4 – Survival estimate for bison in Prince Albert National Park and adjoining lands, Canada, obtained from GPS-collared adult females (2005-2016) ... 85 2.7.5. Appendix 5 – Stochastic projections of the abundance of female bison in Prince Albert national park and adjoining lands, Canada, for next 50 years
under parameters resulting from the percentage of time spent in fields with
hunting permission ... 86
Chapitre 3. Crop raiders caught in an ecological trap: individual-based models of optimal foragers identify effective solutions ...87
3.1. Résumé ... 88
3.2. Abstract ... 89
3.3. Introduction ... 90
3.4. Materials and Methods ... 93
3.4.1. Scenarios of intervention in virtual landscapes ... 93
3.4.2. Habitat, Global Positioning System (GPS) and harvest data ... 94
3.4.3. Model description ... 95
3.4.4. Model parametrising and calibration ... 97
3.4.5. Model validation ... 100
3.5. Results ... 101
3.5.1. Model parametrising and calibration ... 101
3.5.2. Model validation ... 102
3.5.3. Scenario testing ... 102
3.6. Discussion ... 103
3.7. Supplementary Material ... 113
3.7.1. Appendix 1 – Parallel and high-performance computing in individual-based simulations on bison movement and harvest rates in Prince Albert national Park and adjoining lands ... 113
3.7.2. Appendix 2 – Methods for the step selection function (SSF) used to parametrise the movement of bison agents in an individual-based model ... 115
3.7.4. Appendix 4 – Percentage of locations of simulated bison in meadows
before and after the simulated drainage of all meadows (Scenario 1b) .. 117
Conclusion générale ...118
1. Contributions à l’écologie et à la gestion de la faune ... 119
2. Limites et perspectives ... 124
3. Synthèse pour informer la gestion de la population d’étude ... 131
Annexe A – Synthesis to inform the management of the study population ...139
Liste des figures
Introduction générale
Figure 1. Schéma conceptuel de l’écologie spatiale. La valeur adaptative d’un
individu, relative aux autres individus de la population, est le résultat des conditions, des ressources et des risques qu’il retrouve lors de son utilisation de l’espace. L’intégrale de la valeur adaptative de tous les individus détermine les taux vitaux et donc le taux de croissance de la population. La viabilité de la population est alors le taux de croissance projeté dans le futur. La densité de la population a un effet de rétroaction sur le taux de croissance et, par conséquent, sur la viabilité. La densité peut également impacter l’hétérogénéité de l’habitat (p. ex, surexploitation des ressources), ainsi que l’utilisation de l’espace (p. ex, évitement ou attirance vers les congénères) et la valeur adaptative des individus (p. ex, effets d’Allee). Adapté de Matthiopoulos et al. (2015). ... 3
Figure 2. Aire de répartition de la population de bisons des prairies du Parc
National de Prince Albert (PNPA), Saskatchewan, Canada. PCM : polygone convexe minimal. ... 24
Chapitre 1
Figure 1.1. Relative odds (eβ, with 95% confidence interval) in the
summer-autumn of selecting a meadow (left) and remaining in a meadow ≥ 1 hour (right) for bison and wolves in Prince Albert National Park, Canada, as a function of the long-term spatial utilization distribution (UD) of the opponent, from 2007 to 2016. Horizontal lines (eβ =1) represent absence of selection or avoidance. Mean
wolf UD for bison locations: 0.086 (range 0.004 to 44.39). Mean bison UD for
wolf locations: 0.60 (range 0.038 to 36.98). UD values were normalised and scaled from 0 to 100. ... 48
Figure 1.2. Overview of the bison-wolf spatiotemporal game in Prince Albert
reflects the fact that bison distribution is mostly governed by food distribution. Risky times refer to prey responses to immediate risk of predation. Risky places refer to prey responses to the spatial, long-term risk of predation. The Shell Game occurs when prey are constantly on the move to avoid predators capable of remembering prey locations. Key events occurring within each season are illustrated in the inner circle. Icons proportionally illustrate group size and composition for each species. ... 49
Annexe au chapitre 1
Figure S1.1. Direction of movement with respect to the location of the nearest
collared wolf for GPS-collared female bison in Prince Albert National Park, Canada, from 2007 to 2016, when the predator was either far (> 150m) or close (≤ 150m) in winter. The value of 150 m was chosen to illustrate model effects, but the analysis was conducted on a continuous scale. A value of –1 indicates bison movement in the opposite direction of wolf position, whereas a value of +1 indicates the opposite. Observed steps: N=34,742 and N=76 for left and right panels above respectively. Random steps: N=347,420 and N=760 for left and right panels respectively. ... 54
Chapitre 2
Figure 2.1. Overview of the model used to evaluate the sustainability of the
harvest driving an ecological trap for bison in lands adjoining Prince Albert national park, Canada. Subscript t indicates that values vary with time. Variable
h determines the presence (=1) or absence (=0) of harvest. Other refers to wolf
predation and additional causes of death (e.g. accidents, old age). The value of adult female survival (𝑃3𝑡) for each simulation was drawn randomly between 𝑃3𝑚𝑖𝑛𝑡 and 𝑃3𝑚𝑎𝑥𝑡, the calculation of which is shown in the horizontal brackets. 𝑃3𝑒𝑙𝑜𝑤 and 𝑃3𝑒ℎ𝑖𝑔ℎ are constants representing the lowest (0.71) and highest (0.87) values of adult female survival estimated empirically. Parameter
of compensation, and a is the proportion of mortality due to anthrax. Other vital rates were also drawn from empirical intervals, with the exception of P2 for which only one estimate was available. P2 = G2. Initial stage numbers were drawn randomly (see main text). 𝑵𝑡 is the stage structure vector and I is the 3 × 3 identity matrix. ... 77
Figure 2.2. Stochastic projections of the abundance of female bison in Prince
Albert national park and adjoining lands, Canada, for the next 50 years under three scenarios. Causes of mortality are harvest by humans, anthrax outbreaks (disease) and other (i.e. wolf predation, accidents and old age). Presence of harvest mortality indicates that the population is caught in an ecological trap (i.e. maladaptive habitat selection, see main text). Graphs show the 1000 simulations of each scenario (in red or blue) along with the mean (straight black line) and the associated 95% confidence interval (dashed black lines). ... 78
Figure 2.3. Agricultural fields with and without hunting permission used by
bison in lands adjoining the southwest corner of Prince Albert national Park, Canada, from July to November 2005-2016. The histogram shows the percentage of locations on fields with hunting permission obtained from 48 GPS-collared adult females monitored during the same period. Fields 1 to 5 alone accounted for 72% of locations on fields with hunting permission and were the site of 70% of all bison harvest events recorded on fields from 2011 to 2016. ... 79
Annexes au chapitre 2
Figure S2.1. Proportion of harvested individuals per sex (A) and stage (B) per
year from 2011 to 2016 (mean = 23 individuals harvested/year, range: 20 to 26 individuals), and proportion of individuals per stage (C) in the total population per year from 2011-2013 and 2015-2017 for plains bison in Prince Albert national park and adjoining lands, Canada. Each point is a year, while the red
Figure S2.2. Stochastic projections of the abundance of female bison in Prince
Albert national park and adjoining lands, Canada, for the next 50 years under several sources of mortality and parameter values yielding a stable population (i.e. mean population growth rate 𝜆 = 1). Graphs A and B exclude harvest mortality but include compensation through wolf predation and other minor causes of death. Graphs C and D include harvest mortality, wolf predation and the other minor mortality sources. Proportion of mortality due to harvest: p. Proportion of compensatory mortality: c. All 1000 simulations (grey) are shown along with the mean (straight black line) and the associated 95% confidence interval (dashed black lines). ... 84
Figure S2.3. Estimated survival rate for adult female bison in Prince Albert
National Park and adjoining lands, Canada, obtained from GPS-collared females from 2005 to 2016 (N = 48, 24 of which died during the interval, 12 from harvesting and 12 from combined wolf predation and other causes such as accidents and anthrax). ... 85
Figure S2.4. Stochastic projections of the abundance of female bison in Prince
Albert national park and adjoining lands, Canada, for the next 50 years given the percentage of time in the year bison spend on fields with hunting permission (Tperm). The value of 10% corresponds to the mean observed for 30 GPS-collared
females from July to November 2011-2016 (graphs A and B). Graphs C and D show the value of Tperm needed for mean population growth rate = 1. All 1000
simulations (grey) are shown along with the mean (straight black line) and the associated 95% confidence interval (dashed black lines). ... 86
Chapitre 3
Figure 3.1. Structure and process scheduling of the individual-based model
used to simulate bison movement and harvest rates in Prince Albert National Park and adjoining lands, Canada. Agents are either bison or hunter. Scores for inter-polygon movement are the sum of selection coefficients for landscape attributes... 109
Figure 3. 2. A section of bison range in Prince Albert National Park (PANP) and
parametrising, validating and testing management scenarios with an individual-based model (IBM) simulating bison movement and harvest rates. A: The landscape in 2016, which served as template for intervention scenarios. Fields 1 and 2 were cultivated with high profitability crops (alfalfa/timothy) for bison in 2013 (not shown) but low profitability crops in 2016 (wheat and canola). B: Raster layer used to simulate hunter movement. C: Water distribution in meadows inside PANP as observed in the 1990s. D: No high profitability crop in fields with hunting permission. Profitability is digestible energy intake divided by handling time (kJ/min). ... 111
Figure 3.3. Percentage of bison locations outside Prince Albert National Park
(PANP), Canada, and number of harvested females obtained from individual-based simulations comparing the relative effectiveness of changes to food distribution and profitability in reducing both. N = 40 simulations for each scenario. Gray points show values for each simulation, whereas black squares show the mean. * denotes p < 0.05, i.e. no overlap between non-parametric bootstrap 95% confidence intervals of current conditions (2016) and the scenario of interest. Percentage of locations outside the protected area is an index of the intensity of crop raiding by bison. Increase in natural forage was obtained by simulating drainage of water-logged meadows. Profitability is digestible energy intake divided by handling time (kJ/min). ... 112
Annexes au chapitre 3
Figure S3.1. Schematic representation of the buffer zone of redundant
landscape between two adjacent sections of a virtual landscape partitioned for the parallel processing of agent-based simulations of bison and hunter movement in Prince Albert national park and adjoining lands, Canada. The size of the buffer – covering the maximum distance that could be travelled by the hunter, the agent capable of travelling the furthest – allowed all agents to move
movement in an individual-based model (simulated). As all simulations resulted in a similar step length distribution, only one simulation is shown here. ... 116
Figure S3.3. Percentage of locations of simulated bison in meadows in Prince
Albert National Park, Canada, before (current conditions) and after drainage of all meadows used by bison inside the park. Total meadow area inside the park was multiplied by 2.2 following the simulated drainage of meadows. N = 40 simulations for each scenario. Gray points show values for each simulation, whereas black squares show the mean. ... 117
Liste des tableaux
Chapitre 1
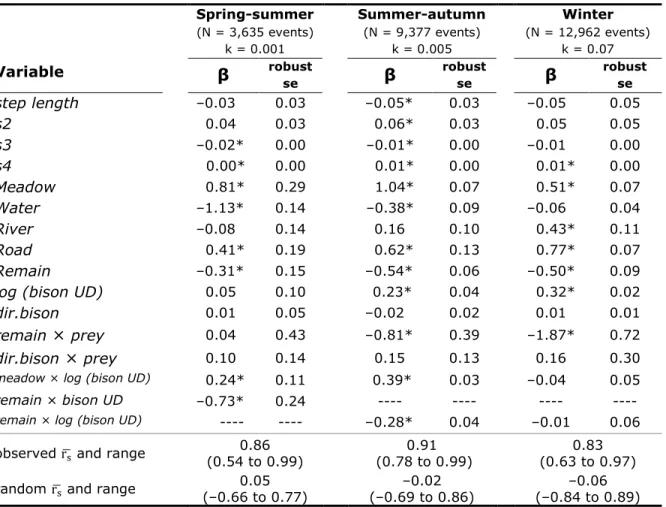
Table 1.1. Parameter estimates (β) of linear mixed-effects models assessing
the influence of meadow profitability on the intensity of meadow use by GPS-collared wolf and female bison in Prince Albert National Park, Canada, during three seasons from 2007 to 2016. ... 45
Table 1.2. Parameter estimates (β) and associated robust standard errors (se)
for bison habitat selection and movement models in response to wolf proximity and long-term use of space in Prince Albert National Park, Canada, for three seasons from 2007 to 2016. ... 46
Table 1.3. Parameter estimates (β) and associated robust standard errors (se)
for wolf habitat selection and movement models in response to bison proximity and long-term use of space in Prince Albert National Park, Canada, for three seasons from 2007 to 2016. ... 47
Annexes au chapitre 1
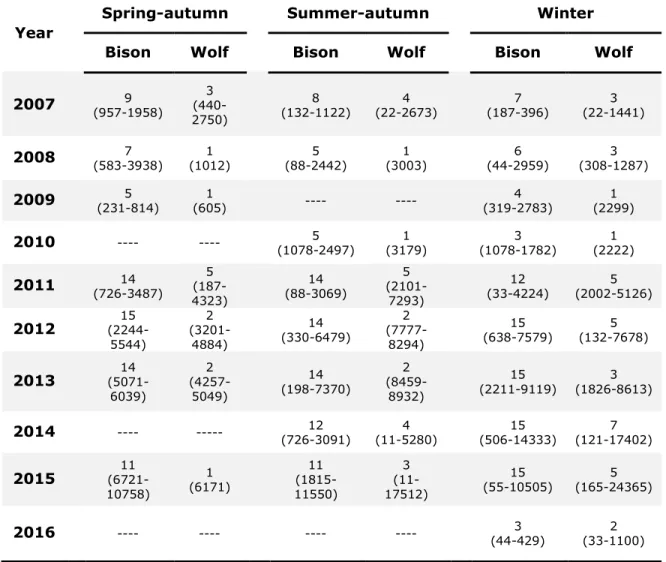
Table S1.1. Number of individuals and number of GPS locations (range, i.e.
minimum and maximum) across seasons and years for bison and wolves in Prince Albert national park, Canada, used to assess their predator-prey spatiotemporal game from 2007 to 2016. ... 50
Table S1.2. Variance inflation factors for covariables used in step selection
function models investigating habitat selection and responses of female bison to wolf distribution in Prince Albert national park, Canada, from 2007 to 2016. . 51
Table S1.3. Variance inflation factors for covariables used in step selection
function models investigating habitat selection and responses of wolves to bison distribution in Prince Albert national park, Canada, from 2007 to 2016. ... 52
Table S1.4. Parameter estimates for bison movement with respect to wolf
Chapitre 2
Table 2.1. Estimates of abundance and associated 95% confidence intervals
obtained from photographic capture-mark-recapture for female bison caught in an ecological trap in lands adjoining Prince Albert National Park, Canada, from 2011 to 2013 and 2015 to 2017. ... 75
Table 2.2. Estimates of adult female survival with 95% confidence intervals and
minimum estimates of the proportion of total mortality due to human harvest per year for adult female bison caught in an ecological trap in lands adjoining Prince Albert national park, Canada, from 2011 to 2017. Survival estimates were obtained through photographic capture-mark-recapture. ... 75
Annexes au chapitre 2
Table S2.1. Coefficients and 95% confidence intervals (CI) obtained from linear
models with no intercept relating measurement ratios of bison horns and face obtained from photographs made by measurer 1 and measurer 2, and measurer 1 and measurer 3. Coefficients were used to eliminate differences between measurers. ... 81
Table S2.2. Mean and standard deviation of calf-, juvenile-, 1-year-old and
2-year-old-to-adult female ratios obtained empirically for the bison population of Prince Albert national park, Canada, from 2015 to 2017. ... 83
Chapitre 3
Table 3.1. Selection coefficients (β) for landscape attributes and step length for
bison agents in an individual-based model (IBM) simulating bison movement and harvest rates in Prince Albert national park and adjoining lands, Canada. β for variables shown with robust standard errors (se) and p-values were obtained from an empirical step selection function model using data from 2013 and 2016 (N = 27,829 steps observed for 20 GPS-collared female bison). β for unused and polygon area were imposed to reproduce observed patterns. ... 107
Table 3.2. Patterns of observed and simulated bison movement and harvest in
Prince Albert National Park and adjoining lands, Canada. Patterns were used to both parametrise and validate the individual-based model used. ... 108
Conclusion générale
Tableau 1. Processus biologiques pertinents à la population de bisons des
prairies du parc national de Prince Albert pouvant être inclus dans un modèle basé sur l’individu. Les références sont à titre indicatif et ne constituent pas une liste exhaustive. ... 131
« my fate
like the fox I run with the hunted and if I’m not the happiest man on earth I’m surely the luckiest man alive. »
Charles Bukowski, The Pleasures of the Damned, Poems 1951-1993
« A man will always lack something of being a really good woodsman, in the finer sense, until he is so steeped in the atmosphere of the Wild and has become so possessed, by long association with it, of a feeling of close kinship and responsibility to it, that he may even unconsciously avoid tramping on too many flowers on his passage through the forest. »
Remerciements
J’aimerais remercier d’abord mon directeur de thèse, Daniel Fortin, pour ses idées et ses questionnements qui ont contribué à améliorer mon travail, de m’avoir appris le besoin de rigueur et de précision dans les propos, d’avoir peaufiné mon écriture scientifique, et d’avoir assuré l’essentiel des conditions matérielles et financières pour le déroulement de ma thèse.
Je tiens également à remercier Gilles Gauthier et Steeve Côté, membres de mon comité d’encadrement, pour des discussions utiles sur l’avancement de mes travaux. Merci aussi à Julie Turgeon pour son soutien tout au long de ma thèse, et à Simon Chamaillé-Jammes du CNRS pour avoir accepté d’être l’examinateur externe de mon travail.
Je voudrais remercier le Conseil de Recherches en Sciences Naturelles et en Génie (CRSNG) du Canada, le Centre d’Études de la Forêt (CEF), le Département de Biologie de l’Université Laval, les Fonds Richard-Bernard et l’État français pour divers soutiens financiers.
At Parks Canada, I would like to thank my co-author Seth Cherry for his contribution and useful discussions, and Joanne Watson for her invaluable help, particularly during fieldwork. I am also grateful to Norman Stolle, Glenn Rupert, Dustin and Digit Guedo, Todd Shury, Danielle Thompson, Jana Curran, Justin Shave, Theresa Stene and Peter deMontigny for their contributions. Prince Albert National Park will forever be a very special place for me.
Un énorme merci à mes assistant(e)s de terrain Yolande Boyer, Étienne Belles-Isles, Emmanuel Jean et Marie-Pier Poulin pour leur aide majeure et pour les aventures que nous avons vécues ensembles !
J’aimerais remercier mes actuels et anciens collègues de laboratoire Marie-Caroline Prima, Guillemette Labadie, Alexandre Terrigeol, Virginie Vanlandeghem, Audrey-Jade Bérubé, Léa Harvey, Florian Barnier, Olivia Tardy, Tom Mason, Marie Sigaud et Jerod Merkle pour des bouts de code R, des révisions de manuscrits, des discussions diverses et surtout pour le partage
d’expériences. Je remercie aussi les étudiant(e)s aux laboratoires de Gilles Gauthier, Steeve Côté et Jean-Pierre Tremblay, et les membres du Comité de Protection aux Animaux de l’Université Laval. Je tiens aussi à souligner l’aide essentielle de Laurent Duchesne et surtout de Maxime Boissonneault de Calcul Québec pour l’utilisation du modèle basé sur l’individu présenté au chapitre 3. Merci aussi à Marie-Claude Martin, pour son aide sur divers volets de ma thèse, et au personnel du secrétariat du Département de Biologie – Louise Lapointe, Marie-Soleil Desjardins et surtout Josée Verret – pour leur aide administrative.
I thank Kelly Paul and members of the Sturgeon River Bison Stewards, as well as landowners who allowed access to their fields, particularly Phil Aarrestad, Robert Gaudry and Errol Vaadeland and family. I am grateful to Lanson Gaudry for the great time we had together, particularly when out there looking for buffalo! I thank several native hunters for interesting conversations, particularly Tim Peekekoot from Athakakoop Cree Nation for sharing his beliefs with me in an unforgettable setting. I am also grateful to Gord Vaadeland at the Canadian Parks and Wilderness Society for the opportunity of getting my research out into the real world of conservation policy and advocacy.
Je suis profondément redevable à Dan Simon, Pierre Dupont, Amandine Pierre, Jana Curran, Diana Gennari Azevedo et Marie-Caroline Prima pour m’avoir écouté et encouragé lors de mes moments de doute et de déprime. Merci…! Je tiens à remercier également Antoine Gendreau-Turmel et Alain Mercier de JimiJam pour notre aventure musicale, un autre élément crucial qui m’a permis de tenir. Et MK, j’en ai hâte aussi !
Agradeço enfim à minha família, Nouailhetas e Simon, pelo apoio incondicional e pelo amor que sempre manifestaram por mim. Amo todos vocês. Eu gostaria em particular de agradecer à minha irmã Monique, a verdadeira cientista da família, pelo apoio nos moments difíceis. Eu tenho muito orgulho de
Avant-propos
Ce doctorat se présente sous la forme d’une thèse avec trois articles scientifiques. La thèse comporte une introduction et une conclusion générales qui lient l’ensemble des articles, bien que ces derniers puissent être lus séparément.
En tant que premier auteur des trois articles, j’ai élaboré les objectifs de recherche en collaboration avec mes co-auteurs, j’ai effectué toutes les analyses et j’ai rédigé la première version complète des articles. Daniel Fortin, mon directeur de thèse, a contribué largement aux étapes de planification de la recherche et aux révisions des manuscrits. Une partie des données que j’ai utilisées a été recueillie par Daniel Fortin, par d’autres étudiants sous sa supervision et par le personnel de Parcs Canada.
Mon chapitre 1 est sous presse dans Oikos, avec Daniel Fortin et Seth Cherry (Parcs Canada) comme co-auteurs. Mon chapitre 2 est publié dans
Biological Conservation, alors que mon chapitre 3 est en cours d’évaluation dans Ecological Applications. Daniel Fortin est mon seul co-auteur pour ces deux
derniers chapitres. Les chapitres 1 et 2 de ce document sont essentiellement les versions publiées, mais avec quelques ajouts ou modifications mineurs en réponse à des commentaires du jury d’évaluation de la thèse.
J’ai également contribué à un article en dehors du travail présenté dans cette thèse. J’ai réalisé des analyses spatiales et j’ai participé à la révision de l’article de Sigaud et coll. qui sera resoumis au Journal of Animal Ecology après révision. J’y suis le deuxième auteur.
Introduction générale
1.
L’écologie spatiale pour relier répartition spatiale et viabilité :
cadre théorique et exemples empiriques
L’objectif ultime de l’écologie est de comprendre et éventuellement de prévoir la répartition et l’abondance des organismes dans différents contextes (Begon et al. 1996, Krebs 2009). Ces deux phénomènes sont liés car les conditions (p. ex., la température et les précipitations), les ressources (p. ex., la nourriture et les abris) et les risques de mortalité que les individus retrouvent dans l’espace déterminent leur probabilité de survivre et de se reproduire, et donc l’abondance de leur population (Begon et al. 1996, Krebs 2009). Un chantier majeur de recherche est alors de comprendre les mécanismes sous-jacents qui gouvernent l’utilisation de l’espace des animaux mobiles, ce qui revient à déterminer où, quand, comment et pourquoi les animaux se déplacent (Nathan et al. 2008). Ces mécanismes incluent tant les besoins (se nourrir, s’abriter, se reproduire), le degré de mobilité, la connaissance de l’environnement et les capacités de navigation des animaux que la répartition spatio-temporelle des conditions, des ressources et des risques dans l’espace et dans le temps (Nathan et al. 2008). Sur cette base mécaniste, il est possible de bâtir un pont entre les patrons d’utilisation de l’espace et les patrons d’abondance des animaux (Morales et al. 2010, Matthiopoulos et al. 2015). L’utilité de cette approche est non seulement de permettre une meilleure compréhension des patrons écologiques, mais également d’aboutir à des principes et à des pratiques plus efficaces de gestion (Levin 1992, Chave 2013), comme il sera illustré dans cette thèse.
DeCesare et al. 2014, Matthiopoulos et al. 2015, Torney et al. 2018). Le point de départ conceptuel (Fig. 1) est que la valeur adaptative d’un individu dépend de ses réactions à la répartition hétérogène des conditions, des ressources et des risques qu’il retrouve dans l’espace et dans le temps (Morris 2003, Matthiopoulos et al. 2015). Les individus devraient alors faire des choix de sélection d’unités spatiales (p. ex., une parcelle de nourriture) en fonction des coûts et des bénéfices pour la survie, la croissance et la reproduction qui y sont associés (Morris 2003, DeCesare et al. 2014). Par exemple, se nourrir sur une parcelle permettrait à un individu d’acquérir de l’énergie pour sa survie et sa reproduction (bénéfices), mais pourrait également l’exposer à un risque accru de mortalité par la prédation (coûts). Des études empiriques ont en effet réussi à mettre en évidence des liens entre l’utilisation de l’espace des individus et des indices de valeur adaptative. L’utilisation d’indices s’impose car il est difficile, voire impossible, de mesurer directement la valeur adaptative d’un individu, car il s’agit d’un concept relatif à tous les individus d’une population (Begon et al. 1996, Gaillard et al. 2010). Il a été démontré, par exemple, que des femelles adultes de chevreuil (Capreolus capreolus) et de cerf rouge (Cervus elaphus) ayant accès ou sélectionnant davantage des parcelles de nourriture de haute qualité ont eu un succès reproducteur à vie (c.-à-d., le nombre de descendants produits par un individu au cours de son existence) supérieur à des congénères utilisant des parcelles de moindre qualité (McLoughlin et al. 2006, McLoughlin et al. 2007).
Figure 1. Schéma conceptuel de l’écologie spatiale. La valeur adaptative d’un individu,
relative aux autres individus de la population, est le résultat des conditions, des ressources et des risques qu’il retrouve lors de son utilisation de l’espace. L’intégrale de la valeur adaptative de tous les individus détermine les taux vitaux et donc le taux de croissance de la population. La viabilité de la population est alors le taux de croissance projeté dans le futur. La densité de la population a un effet de rétroaction sur le taux de croissance et, par conséquent, sur la viabilité. La densité peut également impacter l’hétérogénéité de l’habitat (p. ex, surexploitation des ressources), ainsi que l’utilisation de l’espace (p. ex, évitement ou attirance vers les congénères) et la valeur adaptative des individus (p. ex, effets d’Allee). Adapté de Matthiopoulos et al. (2015).
Outre le succès reproducteur à vie, la probabilité de survie (ou son complément, la probabilité de mortalité) figure parmi les indices les plus utilisés pour relier l’utilisation de l’espace à la valeur adaptative. Ceci est particulièrement vrai dans le cas des grands herbivores, où la survie des femelles adultes détermine en grande partie le taux de croissance à long terme des populations (Gaillard et al. 2010). Losier et al. (2015), par exemple, ont révélé que lorsque la proportion des coupes forestières était relativement élevée dans
de forêt en régénération sont fortement sélectionnées par des orignaux (Alces
alces) en raison d’une abondance de nourriture. La présence d’orignaux attire
les loups, ce qui augmente alors le risque de prédation pour les caribous. Cet exemple sert par ailleurs à illustrer que les individus d’une population ne réagissent pas tous de la même manière à l’hétérogénéité de l’habitat. Ainsi, certaines tactiques d’utilisation de l’espace peuvent être disproportionnellement plus responsables que d’autres pour un patron quelconque observé au niveau de la population.
La prochaine étape dans le schéma de l’écologie spatiale (Fig. 1) représente un changement d’échelle d’organisation biologique : l’intégrale de la valeur adaptative des individus résulte dans les taux vitaux (survie et reproduction) de la population (Matthiopoulos et al. 2015). Ceux-ci déterminent à leur tour le taux de croissance et donc la dynamique de la population dans le temps (Matthiopoulos et al. 2015). Par simple extension, il est alors possible de relier l’utilisation de l’espace des individus à la viabilité de la population (Fig. 1). Cette viabilité correspond à la probabilité de persistance de la population dans le temps, telle qu’évaluée à partir de projections du taux de croissance de la population dans le futur (Beissinger and McCullough 2002, Morris and Doak 2002). Fryxell et al. (2005), par exemple, ont démontré que la viabilité des populations de gazelles de Thomson (Gazella thomsoni thomsoni) requiert que les animaux aient accès à une surface de plus de 1 600 km² de prairies pour persister face à une répartition éphémère dans l’espace et dans le temps de leur nourriture. DeCesare et al. (2014), quant à eux, ont relié des patrons spatiaux de survie des femelles adultes de caribou à la répartition spatiale de leurs ressources et du risque de prédation par le loup pour créer une carte de la répartition spatiale du taux de croissance de la population.
Des modèles de dynamique des populations qui prennent en compte les liens entre l’hétérogénéité de l’habitat et le taux de croissance de la population, comme illustré ci-dessus, sont dits « spatialement explicites ». Des simulations ont démontré que le taux de croissance prévu par ces modèles s’ajuste mieux
au vrai taux de croissance de la population que l’équivalent prévu par des modèles non-spatiaux (Matthiopoulos et al. 2015). De plus, puisque les modèles spatiaux prennent en compte les mécanismes qui régissent la répartition spatiale des individus ou de la population, ils sont également plus performants pour anticiper le taux de croissance d’une population soumise à un nouveau paysage ou à des conditions environnementales nouvelles (Morales et al. 2010, Matthiopoulos et al. 2015, Doherty and Driscoll 2018). Leur utilité dans un contexte où les activités humaines continuent de modifier les habitats de la planète et créent des nouveaux écosystèmes (Sih 2013, Corlett 2015) est donc évidente. Pourtant, malgré un cadre théorique robuste et un fort potentiel pratique, l’utilisation de ces modèles en écologie fondamentale et appliquée est encore limitée. La mise en lumière des liens entre les mécanismes d’utilisation de l’espace des individus et la persistance des populations est, en effet, un chantier encore largement inachevé (Matthiopoulos et al. 2015, Torney et al. 2018). Le défi est essentiellement de démêler l’importance relative des divers facteurs – tels que la physiologie, les interactions sociales, la répartition spatio-temporelle des ressources, la mémoire, le risque de prédation, etc. – sur les choix de déplacements des animaux (Torney et al. 2018).
2. Le rôle de la prédation dans l’abondance et la répartition des
proies
La prédation est, dans sa définition la plus large, la consommation d’un organisme, la proie, par un autre, le prédateur, qui attaque lorsque la proie est encore en vie (Begon et al. 1996). Cette définition inclut ainsi le parasitisme et la consommation des végétaux par les herbivores. Dans le cadre de cette thèse, cependant, il sera question notamment des « vrais prédateurs », c’est-à-dire, des organismes qui tuent leurs proies presque instantanément après une attaque réussie, et qui au cours de leur vie consomment plusieurs proies (Begon
L’une des plus anciennes questions de recherche en écologie est de comprendre l’impact de prédateurs sur l’abondance de proies, et vice-versa. Dans un système simple avec une seule espèce prédatrice et une seule espèce proie – et faisant notamment abstraction de l’espace – la prévision théorique fondamentale est une fluctuation cyclique et couplée dans le temps de l’abondance des prédateurs et des proies (Begon et al. 1996, Bonsall and Hassell 2007). Une augmentation du nombre de proies devrait engendrer une augmentation du nombre de prédateurs, suivie d’une diminution du nombre de proies en raison d’un niveau de prédation accru, suivie enfin d’une diminution du nombre de prédateurs en raison d’une disponibilité de proies moindre. Une réduction du niveau de prédation permettrait alors à la population des proies d’augmenter à nouveau, et le cycle repart. Les travaux classiques de Lotka et Volterra ont réussi à modéliser cette dynamique intuitive à l’aide d’équations différentielles qui relient le nombre de prédateurs au nombre de proies et vice-versa (cf. Bonsall and Hassell 2007).
L’exemple empirique incontournable des fluctuations cycliques dans les populations de prédateurs et de proies est celui du lièvre d’Amérique (Lepus
americanus) et du lynx du Canada (Lynx canadensis) dans la forêt boréale de
l’Amérique du Nord. À partir des archives de la Compagnie de la Baie d’Hudson sur le nombre de fourrures récoltées annuellement, Elton (1924) a mis en évidence des cycles d’abondance relativement réguliers et couplés d’environ une dizaine d’années entre les deux espèces. Le lynx du Canada est un spécialiste dont le régime alimentaire est fortement basé sur le lièvre d’Amérique, de sorte que c’est l’abondance de lièvres dans une année donnée qui détermine l’abondance de lynx quelques années plus tard (Stenseth et al. 1997). La question fondamentale est alors d’identifier la cause du cycle dans les populations de lièvre : est-il dû effectivement à la prédation – par le lynx mais également par une pléthore d’autres prédateurs aviaires et terrestres – ou plutôt dû aux variations de disponibilité de nourriture (Sinclair et al. 2000) ? Cette question s’insère dans un débat plus large sur le rôle des mécanismes dits de bas vers le haut (« bottom-up », c.-à-d., basés sur la limitation des ressources)
ou de haut vers le bas (« top-down », c.-à-d., basés sur la prédation) dans la régulation de l’abondance des populations animales et dans la structuration des communautés écologiques (Polis 1999, Oksanen and Oksanen 2000, Sinclair et al. 2000).
Une série remarquable d’observations empiriques, des manipulations expérimentales et des simulations débutant dans les années 1970s et continuant de nos jours ont apporté quelques éléments de réponse pour expliquer les cycles du lièvre d’Amérique (voir un aperçu historique et l'état de l'art dans Krebs et al. 2018). C’est finalement la prédation, et non la disponibilité des ressources, qui semble en être la cause principale. L’impact des prédateurs sur le taux de croissance des lièvres se fait de deux façons: directement par la consommation d’individus, mais aussi indirectement par une suppression du succès reproducteur des femelles en raison du stress engendré par des poursuites récurrentes (Krebs et al. 2018).
Cet effet indirect des prédateurs, non lié à la consommation, sur la démographie des proies par la peur (Brown et al. 1999, Ripple and Beschta 2004, Stankowich and Blumstein 2005, Preisser and Bolnick 2008, Clinchy et al. 2013) pourrait bien figurer parmi les découvertes les plus extraordinaires des 20 dernières années en écologie. L’un des exemples les plus connus, mais controversé, vient de l’interaction entre des loups réintroduits et des wapitis dans l’écosystème du grand Yellowstone aux USA. Creel et al. (2007) ont étudié plusieurs populations hivernales du cervidé pour démontrer des corrélations entre la seule présence des loups et la démographie des wapitis : plus le nombre de loups dans l’aire de répartition de la population était élevé, moins il y avait de wapitis femelles gestantes, et moins il y avait de faons l’année suivante. Ces corrélations suggèrent que les comportements anti-prédateurs des wapitis, bien que réduisant le risque de mortalité, engendrent aussi des coûts (Creel et al.
répondu à cette remise en cause de leurs résultats en faisant la démonstration que la méthodologie utilisée par Middleton et al. (2013) pourrait sous-estimer la fréquence et l’intensité des réactions anti-prédatrices des proies par un facteur de plus de dix. Quoi qu’il en soit, l’exemple illustre un concept fondamental dans l’interaction prédateurs-proies : la nécessité, pour les proies, de trouver un compromis coûts-bénéfices entre se nourrir pour survivre et se reproduire, et éviter de se faire tuer (Lima and Dill 1990, Lima 1998). Les comportements anti-prédateurs sont alors un élément capital reliant la répartition d’une population de proies à son abondance (Fig. 1).
Un autre point conceptuel essentiel à comprendre dans l’interaction prédateurs-proies est que l’intensité des réactions anti-prédatrices des proies n’est pas forcément positivement corrélée au taux direct de prédation (c.-à-d., au nombre de proies qui sont tuées) (Creel and Christianson 2008). Des investissements relativement élevés en temps et en énergie dans des comportements anti-prédateurs peuvent résulter dans un faible taux direct de prédation s’ils s’avèrent efficaces, par exemple (Creel and Christianson 2008). En outre, il a été démontré que la seule perception du risque de prédation par les proies, même lorsque le risque réel de mortalité était faible, peut déclencher des comportements anti-prédateurs et impacter en cascade les ressources de la proie suffisamment pour entraîner des changements aux communautés écologiques. Par exemple, la simple diffusion d’enregistrements sonores d’aboiements de chien (Canis lupus familiaris) a réduit l’effort d’approvisionnement des ratons laveurs (Procyon lotor) au point de faire presque doubler l’abondance de leur proie préférée, des crabes rouges (Cancer
productus) (Suraci et al. 2016). Cette augmentation de l’abondance des crabes
a induit une réduction de l’abondance d’un compétiteur et de celle de leurs ressources alimentaires. La réintroduction du loup au parc national de Yellowstone aurait aussi engendré une cascade trophique, en favorisant la régénération des peuplements de peupliers faux-trembles (Populus tremuloides) grâce à des changements au comportement d’approvisionnement des wapitis (Ripple et al. 2001, Ripple and Beschta 2004, Fortin et al. 2005, Painter et al.
2018) – bien que cet exemple soit, encore une fois, contesté (Kauffman et al. 2010, Winnie 2012, Beschta et al. 2014, Winnie 2014, Cusack et al. 2019).
Dans tous les cas, l’utilisation de l’espace par les proies est influencée par leurs comportements anti-prédateurs à plusieurs échelles spatiales et temporelles (Lima and Zollner 1996, Lima 1998, 2002). La possibilité d’échapper aux prédateurs a été mise en avant comme un facteur, outre celui d’accéder à une meilleure nourriture, impliqué dans les longues migrations saisonnières entreprises par les caribous dans le Grand Nord de l’Amérique (Rettie and Messier 2000) et par les gnous bleus (Connochaetes taurinus) dans les grandes prairies de l’Afrique australe (Fryxell et al. 1988), par exemple. À une échelle plus fine, il a été démontré que des wapitis réussissent à minimiser le risque de prédation dans leurs domaines vitaux en y utilisant davantage des zones à forte empreinte humaine, évitées par leurs prédateurs (Hebblewhite and Merrill 2009). L’évitement des zones à risque a également été observé chez les wapitis à l’échelle de la journée (Creel and Winnie 2005, Creel et al. 2008) et aux heures (Fortin et al. 2005, Creel et al. 2013, Middleton et al. 2013, Kohl et al. 2018). Diverses espèces herbivores en Afrique réagissent également au risque de prédation à long et à court terme par des lions (Panthera leo), et ceci tant à l’échelle d’une saison qu’à celle de la journée ou du cycle circadien jour-nuit (Valeix et al. 2009a, Valeix et al. 2009b, Palmer et al. 2017).
L’approche ayant possiblement contribuée le plus pour mettre en lumière l’impact de la perception du risque de prédation sur l’utilisation de l’espace de proies est celle basée sur l’utilisation de parcelles. L’observation fondamentale ici est que les ressources alimentaires sont réparties de façon hétérogène dans l’espace et dans le temps ; une parcelle peut alors être définie comme une aire relativement homogène où la nourriture de l’espèce d’intérêt est concentrée à un moment donné (Charnov 1976, Begon et al. 1996). Une question centrale est
l’approvisionnement optimal (« optimal foraging »), qui prévoit que la sélection naturelle ait façonné des stratégies comportementales maximisant l’efficacité de l’approvisionnement de l’animal (Stephens and Krebs 1986, Cezilly and Benhamou 1996). Une prévision théorique est que les animaux devraient quitter une parcelle de nourriture qui s’épuise lorsque leur taux net d’acquisition d’énergie y équivaut au taux moyen de l’habitat (théorème de la valeur marginale, Charnov 1976). Malgré le fait que cette prédiction soit soutenue par quelques études empiriques et expérimentales (cf. Cezilly and Benhamou 1996), d’autres observations empiriques démontrent aussi que les animaux peuvent quitter des parcelles bien avant ce seuil théorique (Mitchell and Lima 2002). Le risque de prédation figure parmi les facteurs qui pourraient expliquer ce départ théoriquement prématuré des parcelles de nourriture (Brown 1988, 1999, Mitchell and Lima 2002).
De nombreuses études, basées tant sur l’utilisation des parcelles que sur la répartition spatio-temporelle du risque de prédation, démontrent le rôle primordial des prédateurs dans l’utilisation de l’espace des proies (cf. Brown and Kotler 2004). Ces approches ont cependant été critiquées pour traiter souvent les prédateurs comme une source plutôt abstraite de risque, fournissant alors une compréhension incomplète des comportements anti-prédateurs des proies (Brown et al. 1999, Lima 2002). La stratégie d’utilisation de l’espace des proies devrait dépendre de celle des prédateurs, et vice-versa. L’approche dite du jeu spatio-temporel permet d’aborder la question du point de vue des deux joueurs.
3. Le jeu spatio-temporel prédateurs-proies
L’interaction prédateurs-proies peut être considérée comme un jeu dans lequel chaque joueur a des objectifs divergents : pour le prédateur, il s’agit de rencontrer et de tuer une proie ; pour la proie, il est question de se nourrir, d’éviter de rencontrer des prédateurs et de ne pas se faire tuer lors d’une rencontre (Brown et al. 1999). La théorie des jeux en écologie prédit alors l’émergence d’une stratégie conjointe dite évolutivement stable (« evolutionary
stable strategy ») entre les prédateurs et les proies, c’est-à-dire, une stratégie conjointe qui maximise la valeur adaptative de chaque joueur en fonction de la stratégie de l’adversaire (Maynard Smith 1982). Du point de vue spatial et lorsque les deux joueurs sont capables de se déplacer, les proies devraient éviter les zones où se concentrent les prédateurs, tandis que les prédateurs devraient sélectionner les zones où se retrouvent les proies (Sih 1984, 2005). Le modèle théorique de Sih (2005) prévoit une stratégie évolutivement stable peu intuitive pour les deux joueurs dans un environnement marqué par une répartition hétérogène des ressources pour les proies : les prédateurs devraient se retrouver davantage sur les parcelles les plus productives en ressources pour les proies, alors que les proies devraient se répartir de façon plus uniforme dans l’espace. Cette hypothèse, selon laquelle la répartition des prédateurs devrait coïncider avec celle des ressources des proies de façon plus marquée que la répartition des proies coïncide avec celle de leurs propres ressources, est connue sur le nom d’effet saute-mouton (« leap-frog effect », Sih 1984, 2005).
Une hypothèse alternative dans le jeu prédateurs-proies est le jeu de passe-passe (« shell game ») proposé par Mitchell et Lima (2002). Ces auteurs ont démontré, à l’aide de simulations, que les proies devraient se déplacer constamment entre les parcelles de nourriture afin de rendre leur localisation imprévisible à des prédateurs qui reviennent aux endroits où ils ont rencontré des proies précédemment (Mitchell and Lima 2002). Bien que le jeu de passe-passe soit parfois considéré ou mis en avant pour expliquer des patrons d’utilisation de l’espace des prédateurs et des proies (p. ex., Laundré 2010, Harvey and Fortin 2013, Mason and Fortin 2017), des exemples empiriques en faisant la démonstration sont rares. Le rôle de la perception du risque de prédation en tant qu’élément déclencheur de l’abandon des parcelles de nourriture avant leur épuisement par les proies reste particulièrement à démontrer. D’autres hypothèses, spécifiques au comportement anti-prédateur
perçoivent comme étant plus à risque à long terme (Creel et al. 2008). La définition de ce qui constitue le risque à long terme varie selon les études, pouvant aller d’une simple présence ou absence des prédateurs dans l’espace à des probabilités d’occurrence ou de succès de chasse au cours des saisons (Moll et al. 2017). Dans le cas de l’hypothèse des moments risqués, les proies ne réagiraient que lorsqu’un prédateur est proche et que le risque est immédiat (Creel et al. 2008, Moll et al. 2017).
Ces hypothèses peuvent être considérées comme différentes tactiques de jeu qui ne sont pas forcément mutuellement exclusives. La stratégie globale d’un joueur peut alors inclure plusieurs tactiques, et ceci selon la configuration spatio-temporelle de l’habitat (le « plateau de jeu », pour rester dans l’analogie) et les tactiques de l’adversaire. Un exemple relativement bien étudié pour illustrer ce point est le jeu spatio-temporel entre le loup, le caribou et l’orignal dans la forêt boréale aménagée de l’Amérique du Nord. Du printemps à l’automne, les caribous évitent ou traversent rapidement les zones de coupe en régénération car le risque d’y rencontrer un loup est relativement plus élevé que dans d’autres types d’habitat (Courbin et al. 2013, Latombe et al. 2014a, Basille et al. 2015). En effet, de l’été à l’automne, les loups se déplacent davantage dans ces zones car elles sont fortement traversées aussi par leur proie principale, les orignaux (Courbin et al. 2013). Lorsqu’un loup est proche ou qu’il a récemment été dans les parages durant cette période de l’année, les caribous évitent les zones riches en nourriture, habituellement sélectionnées, pour se réfugier dans des peuplements denses de conifères (Latombe et al. 2014a, Basille et al. 2015). En hiver, les caribous sélectionnent davantage des parcelles riches en nourriture si elles sont entourées de zones de refuge (Latombe et al. 2014a, Mason and Fortin 2017) ou si un loup a récemment été dans les parages. Dans ce cas, puisque la probabilité qu’un loup revienne sur la zone traversée est relativement faible dans les cinq prochains jours environ, les caribous profiteraient pour utiliser davantage des parcelles riches en nourriture (Latombe et al. 2014a). Les orignaux, pour leur part, ne semblent pas changer leur utilisation de l’espace en réaction au risque de prédation en hiver (Latombe et al. 2014a). Enfin les loups,
quant à eux, sélectionnent plus fortement les parcelles riches en nourriture pour leurs proies durant cette saison, et ceci même si ces parcelles ne correspondent pas aux zones les plus utilisées par leurs proies (Courbin et al. 2014). Cet exemple met aussi en lumière le rôle crucial des variations saisonnières dans la variabilité des tactiques de jeu de prédateurs et de proies (Basille et al. 2013).
Cependant, encore peu d’études ont adopté l’approche de considérer le jeu spatio-temporel prédateurs-proies du point de vue des deux joueurs (Lima 2002, Moll et al. 2017). La possibilité que les différentes hypothèses ne soient pas des tactiques mutuellement exclusives, mais plutôt des parties d’une stratégie plus globale, a aussi rarement été considérée. La prise en compte d’une dynamique temporelle à fine échelle dans le jeu est par ailleurs une évolution assez récente (p. ex., Palmer et al. 2017, Kohl et al. 2018), et pas assez documentée. Notre compréhension actuelle du jeu prédateurs-proies est incomplète et potentiellement trop simpliste. En outre, l’impact de la perception du risque de prédation sur l’utilisation de l’espace des proies de grande taille et relativement dangereuses pour les prédateurs (Mukherjee and Heithaus 2013) demeure méconnu. Ceci est d’autant plus vrai dans les systèmes multi-proies où les prédateurs ont des choix alternatifs de proies, moins dangereuses à attaquer. Bien que des observations empiriques suggèrent que ce sont des mécanismes de bas vers le haut qui gouvernent la répartition des proies de grande taille (p. ex., Sinclair et al. 2003), cela ne vaut pas dire que la perception du risque de prédation est négligeable dans leurs choix d’utilisation de l’espace. Cet oubli nous empêche de bien saisir le rôle des prédateurs dans la structuration et le fonctionnement des écosystèmes, notamment lorsque les proies en question sont des grands herbivores, dont l’utilisation de l’espace a un effet majeur sur l’écosystème (p. ex., sur le recyclage de nutriments ou sur la composition et la structure de la végétation, Danell et al. 2006).
4. L’hétérogénéité spatiale de l’habitat et la prédation
L’hétérogénéité spatiale de l’habitat est un facteur majeur qui permettrait aux prédateurs et aux proies de coexister (Begon et al. 1996, Bonsall and Hassell 2007, Gorini et al. 2012). Dans une série classique d’expériences, Huffaker (1958) et Huffaker et al. (1963) ont investigué le rôle de ce facteur dans l’interaction entre l’acarien prédateur Typhlodromus occidentalis et l’acarien herbivore Eotetranychus sexmaculatus. L’habitat y était essentiellement composé d’oranges, consommées par la proie, intercalées par des balles de caoutchouc. En l’absence du prédateur, l’abondance initiale des proies augmentait rapidement pour ensuite osciller dans le temps, sans événement d’extinction. L’introduction des prédateurs conduisait d’abord à une augmentation rapide de leur abondance au détriment de celle des proies, suivie de l’extinction des proies et des prédateurs eux-mêmes. Mais dans un habitat hétérogène, plus parcellaire et complexe, et dont la configuration favorisait davantage les déplacements des proies que des prédateurs, les deux espèces ont persisté dans le temps. Les proies étaient toujours capables de coloniser une parcelle et d’y proliférer avant l’arrivée des prédateurs.
Des modèles classiques de l’interaction prédateurs-proies, comme celui de Lotka-Volterra, supposent que le taux de rencontre entre les prédateurs et les proies soit simplement dépendant de leurs densités respectives (Bonsall and Hassell 2007). Or, cela n’est pas nécessairement le cas, car les deux acteurs ne font pas une même utilisation de l’espace, tel que déjà illustré par des exemples ci-dessus. L’hétérogénéité de l’habitat impacte par ailleurs non seulement le taux de rencontre, mais aussi l’efficacité de recherche, le taux de succès et même le taux de consommation du prédateur (Gorini et al. 2012). Ainsi, l’hétérogénéité spatiale crée un écart entre l’abondance totale d’une population de proies et la proportion effectivement disponible aux prédateurs (cf. Gorini et al. 2012). Cet écart représente ainsi un mécanisme reliant l’hétérogénéité spatiale de l’habitat à la répartition et à l’abondance d’une population (Fig. 1).
Un point essentiel est que l’hétérogénéité spatiale peut mener à des patrons contre-intuitifs d’abondance et de répartition des prédateurs et des proies en interaction. Murrell (2005), par exemple, a utilisé des simulations pour démontrer qu’une augmentation du taux de reproduction des proies peut, à terme, réduire leur abondance au lieu de l’augmenter. Dans le modèle de Murrell, les proies se déplacent pour éviter les prédateurs et réussissent à mieux le faire lorsque leur densité est relativement faible. L’augmentation de l’abondance des proies, due au taux de reproduction plus élevé, rend les groupes de proies plus nombreux et plus larges, et donc plus facilement repérables par les prédateurs. La conséquence est une augmentation du taux de prédation et une diminution des effectifs des proies à un niveau inférieur au nombre initial précédant l’augmentation du taux de reproduction (Murrell 2005). Ainsi, il est possible que des patrons inattendus d’abondance et de répartition émergent lorsque l’hétérogénéité spatiale est prise en compte dans l’interaction prédateurs-proies.
5. Homo sapiens en tant que prédateur
L’être humain chasse et consomme des animaux, c’est une évidence, mais l’inclusion d’Homo sapiens parmi les prédateurs naturels est discutable. En effet, aucun autre prédateur ne consomme un éventail aussi large de proies. Et puisque son taux d’exploitation du stade adulte des populations animales est jusqu’à 14 fois supérieur à celui de ces concurrents non-humains, l’homme peut être considéré comme un « super-prédateur » dont l’écologie est fondamentalement différente de celle des autres prédateurs de la planète (Darimont et al. 2015). La chasse et la consommation de viande de brousse est, par ailleurs, une menace majeure pour la biodiversité, notamment pour les mammifères et les oiseaux (Ripple et al. 2016, Benitez-Lopez et al. 2017, Ripple et al. 2017, van Velden et al. 2018). Cette exploitation non-durable est