La régulation de l’hepcidine
à travers les récepteurs Toll-like dans les macrophages
Par Antonio Layoun
Département de microbiologie et immunologie Faculté de médecine
Thèse présentée à la Faculté des études supérieures en vue de l’obtention du grade de maitrise
en microbiologie et immunologie
Décembre 2009
Université de Montréal Faculté des études supérieures
Cette thèse intitulée :
La régulation de l’hepcidine à travers les récepteurs Toll-like dans les macrophages
Présentée par : Antonio Layoun
a été évaluée par un jury composé des personnes suivantes :
Dr Marika Sarfati, président-rapporteur Dr Manuela Santos, directeur de recherche Dr Janos G. Filep, membre du jury
Résumé
L'interaction entre le système immunitaire et le métabolisme du fer est bien illustrée par l'anémie des maladies chroniques (ACD), qui est fréquemment rencontrée dans les infections chroniques, l'inflammation et le cancer. La majorité des modifications dans les paramètres du fer observées dans l’ACD tient compte des modifications de l’homéostasie du fer, avec la délocalisation du métal de la circulation et les sites de l'érythropoïèse au compartiment de stockage dans les macrophages. Les mécanismes de la réponse hyposidérémique impliquent des cytokines, notamment TNF-alpha et IL-6, qui régulent les niveaux de plusieurs gènes du métabolisme du fer, y compris les transporteurs de fer et de l'hepcidine, un régulateur négatif de l’absorption du fer, ce qui entraîne l'inhibition de l'exportation du fer à travers la ferroportine 1 (FPN1) au niveau de l'intestin et les macrophages. Des études antérieures ont montré que l'IL-6 induit l’expression d’hepcidine dans les hépatocytes, mais il y a très peu de données concernant la façon par laquelle l'hepcidine et la FPN1 sont régulées dans les macrophages. Récemment, nous avons constaté que l'induction de l'hepcidine dans le foie par le lipopolysaccharide (LPS) dépend de la voie de signalisation médiée par le récepteur Toll-like 4 (TLR4). Le but de ce travail est d’identifier les ligands des TLRs capables d'induire l'hepcidine dans les macrophages et de déterminer l’exigence des TLRs dans l’induction de l’hepcidine et le développement d’hyposidérémie. En plus, nous voulons étudier l’effet de l’inflammation causée par les ligands des TLRs sur le taux de fer sérique, la production des cytokines et l'expression de l’hepcidine et de la ferroportine. D’autre part nous voulons étudier l’effet du taux du fer sur la production d’IL-6 macrophagique en réponse à la stimulation par le TLR4. D'abord, pour identifier les ligands des TLRs capables d'induire l'hepcidine dans les macrophages, nous avons traité les macrophages RAW 264.7 et les macrophages péritonéaux de souris (MPMs) avec différents ligands TLRs et on a mesuré l’expression de l'hepcidine par qRT-PCR. Nous avons observé que Pam3CSK4 (Pam), un ligand de TLR2/1; LPS, un ligand de TLR-4 et FSL1 un ligand de TLR2/6 induisent l’expression de l'hepcidine dans les cellules RAW 264.7 et les MPMs, contrairement au polyinosinic: polycytidylic acid (Poly I: C), un ligand de TLR3. De plus, LPS était capable de réprimer l’expression de la ferroportine dans les cellules RAW 264.7. Afin de mieux définir la nécessité des TLRs pour assurer cette expression, nous avons utilisé les souris TLR-2 knock-out et on a établi que l'expression de l'hepcidine dans les macrophages par LPS, Pam ou FSL1 est dépendante du TLR2. En accord avec les expériences in vitro, les études effectuées in vivo ont montré que LPS réprime l’expression de la ferroportine, ainsi que PolyI:C n’est pas capable de stimuler l'expression d'hepcidine hépatique, par contre il était efficace pour déclencher une hyposidérémie. Ensuite, on voulait déterminer la voie de signalisation utilisée dans l’induction de l’hepcidine dans les macrophages. Comme il y deux voies majeures connues pour la signalisation des TLRs : une dépendante et l’autre indépendante de la protéine MyD88, on a étudié l’expression de l’hepcidine dans les MPMs isolés des souris MyD88-/- et nous avons constaté que l'absence de signalisation MyD88 abolit l'induction de l'hepcidine déclenchée par Pam, LPS et FSL1. D’autre part, la stimulation avec du LPS induisait in vivo la production 6 et de TNF-alpha, et la stimulation d’IL-6 était renforcée in vitro par la présence du fer. Ces observations indiquent que l’expression de HAMP (Hepcidin Antimicrobial Peptide) dans les macrophages peut être régulée par différents TLRs, ce qui suggère que la production d'hepcidine macrophagique fait partie d'une réponse immunitaire activées par les TLRs.
Abstract
The interaction between the immune system and iron metabolism is well exemplified in the anemia of chronic disease (ACD), which is frequently encountered in chronic infections, inflammation and cancer. The major changes in iron parameters observed in ACD ultimately reflect modifications in iron trafficking, with relocation of the metal from both the circulation and sites of erythropoiesis to the storage compartment in macrophages. Mechanisms in the hypoferremic response involve cytokines, including TNF-alpha and IL-6. These pro-inflammatory cytokines regulate the levels of several iron metabolism genes, including iron transporters and hepcidin, a negative regulator of iron absorption, resulting in the inhibition of iron export by ferroportine 1 (FPN1) from the intestine and macrophages. Previous studies showed that IL-6 upregulates hepcidin in hepatocytes, but there are very few data regarding how hepcidin and FPN1 expression is regulated in macrophages. More recently, we found that hepcidin induction in the liver by lipopolysaccharide (LPS) is dependent on the signaling pathway mediated by toll-like receptor 4 (TLR4). The aim of this work is to identify TLR ligands able to induce hepcidin in macrophages and to determine the requirement for TLRs in hepcidin expression and the development of hypoferremia. In addition, we want to study the effect of inflammation induced by TLR ligands on serum iron levels, cytokine production, hepcidin and ferroportin expression. On the other hand we want to study the effect of iron levels on IL-6 production by macrophages in response to TLR4 stimulation. First, to identify TLR ligands capable of inducing hepcidin in macrophages, we treated Raw 264.7 macrophages and thioglycollate-stimulated mouse peritoneal macrophages (MPMs) with various TLR ligands and measured hepcidin and ferroportin expression by real-time RT-PCR. We observed that Pam3CSK4 (Pam), a TLR1/2 ligand; LPS, a TLR-4 ligand; and FSL1 a TLR6/2 ligand, but not polyinosinic: polycytidylic acid (poly I:C), a TLR3 ligand, upregulate hepcidin expression in both Raw 264.7 cells and MPMs. Furthermore, LPS was able to repress ferroportine expression in RAW 264.7 macrophages. To further define the requirement for the identified TLRs, we used TLR-2 knockout mice and established that upregulation of macrophage hepcidin expression by Pam or FSL1 is TLR2 dependent, respectively. In agreement with the in vitro experiments, when tested in vivo LPS repressed ferroportine expression and polyI:C failed to induce hepatic hepcidin expression but was effective in triggering hypoferremia. We next investigated whether MyD88, the predominant but not exclusive intracellular signal transduction pathway for TLR-4, is necessary for hepcidin induction in macrophages. Using MyD88 knockout mice, we found that the absence of MyD88 signaling abolishes hepcidin induction triggered by Pam, LPS and FSL1.On the other hand, stimulation with LPS induced in vivo the production of IL-6 and TNF-alpha, and IL-6 stimulation was enhanced in vitro by high amount of iron in macrophages.These observations indicate that HAMP (Hepcidin Antimicrobial Peptide) expression in macrophages can be regulated through multiple TLRs, suggesting that macrophage hepcidin production is part of an immune response activated by the TLRs.
Liste des abréviations
8-OHG : Acide nucléique 8-hydroxyguanine
ACD : Anémie des maladies inflammatoires chroniques ADN: Acide désoxyribonucléique
Apo-Tf : Apotransferrine AP1 : Activator protein 1 ARN : Acide ribonucléique
ATPase: Adénosine triphosphatase BDH-2/3 : Beta-défensine humaine 2/3 BMP-RE : BMP-responsive elements
BMPRs : Bone morphogenetic protein receptors BSA : Bovine serum albumin
CAT : Catalase
CCPA : Conseil canadien de protection des animaux CD : Cellules dendritiques
CD163 : Cluster of differentiation 163
CIPA : Comité institutionnel de protection des animaux CMH1 : Complexe majeur d’histocompatibilité de classe 1 CP : Céruloplasmine
CPA : Cellules présentatrices d’antigène CpG : Cytosine-guanine
CTFF : Capacité totale de fixation du fer CYBRD1 : Cytochrome b réductase 1 Db : Doubles brins
DD : Death domain DTT : Dithiotreitol DFO : Desferrioxamine
DMEM : Dubelcco's modified eagle medium ECP : Eosinophil cationic protein
FAC : Ferric Ammonium Citrate FACS : Cytométrie en flux FBS : Fetal bovine serum Fe-S : Clusters fer-soufre
FIH : Facteurs inductibles par l’hypoxie FPN1 : Ferroportine1
FS : Fer sérique
FTH : Ferritin heavy chain
GADD45 : Growth Arrest and DNA-Damage Inducible 45 Gene GDF15 : Growth differentiation factor 15
GPI : Glycosylphosphatidylinositol GPL : Lipophosphoglycanes GPx : Gluthatione peroxydase HEPHA : Hephaestine
HAMP : Hepcidin Antimicrobial Peptide Hb : Hémoglobine
HCP1 : Hème Carrier Protein 1 HH : Hémochromatose héréditaire HJV : Hémojuvéline
HJVm : Hémojuvéline membranaire HJVs : Hémojuvéline soluble HO-1 : Hème oxygénase 1 Hp : Haptoglobine
HSV : Herpès simplex virus i.p. : Intrapéritonéale
IFNs : Interférons
IL1 β : Interleukine 1 beta IL-6 : Interleukine 6
IMP : Integrin-Mobilferrin Pathway IRAK : IL-1 Receptor-Associated Kinase
IRE : Iron responsive elements IRF3 : Interferon regulatory factor 3 IRP1 : Iron regulatory proteins 1 IRP2 : Iron regulatory proteins 2 LAM : Lipoarabinomannanes LB : Lymphocytes B
LIP : Labile iron pool Lipocaline 2: LCN2 LOXs : Lipoxygénases LPS : Lipopolysaccharide LRR : Leucine rich repeats LT : Lymphocytes T
MAP kinases: Mitogen-activated protein kinases MCMV : Cytomégalovirus murin
MFI : Mean fluorescence intensity MMTV : Tumeur mammaire murines MPMs : Macrophages péritonéaux murins NaN3: Sodium Azide
NF-κB : Nuclear factor κB NK : Natural killer
NRAMP : Natural resistance-associated macrophage protein NTBI : Non transferrin bound iron
O2 - . : Superoxyde
p53 : Tumor protein 53
PAMPs : Pathogen associated molecular patterns PBS : Phosphate-buffered saline
Poly(I:C) : Polyionosine-polycytidylic acid PRRs : Pattern-recognition receptors PS : Peniciline et Streptomycine RE : Réticulum endoplasmique
RNRs : Ribonucléotides réductases RO- : Alcoxy radical
ROI : Reactive oxygen intermediates ROO- : Alkyl oxyradical
ROOH : Hydroxy peroxydes des lipides RT : Reverse transcription
SDS : Sodium dodecyl sulfate Sb : Simples brins
SOD : Superoxide dismutase
Steap3: Six-transmembrane epithelial antigen of the prostate 3 ST% : Saturation de la transferrine
sTfR1 : Récepteur de la transferrine 1 soluble Tf : Transferrine
TfR : Récepteur de la transferrine TfR2 : Transferrine 2
TGF-β : Transforming growth factor β TH : LT Helper
TIR : Toll/IL-1 receptor TLR4 : Récepteur Toll-like 4 TLRs : Toll-like receptors
TNF-α : Facteur nécrose tumorale α UNC93B1 : Unc-93 Homolog B1 USF2 : Upstream transcription factor 2 UTR : Untranslated regions
Table des matières
I. INTRODUCTION...1
1. Propriétés chimiques et fonctions biologiques du fer ...3
1.1 Les hémoprotéines...4
1.2 Les protéines fer-soufre...5
1.3 Autres protéines contenant du fer...6
1.4 Protéines liant le fer...8
2. Toxicité du fer...9
3. Homéostasie cellulaire du fer...11
3.1 Captation du fer...11
3.2 Capture cellulaire du fer par des mécanismes dépendants de la Transferrine...11
3.2.1 Récepteur de la transferrine (TfR)...11
3.2.2 Cycle de la transferrine...13
3.3 Capture cellulaire du fer par des mécanismes indépendants de la Tf ...15
3.4 Devenir du fer cellulaire...17
3.5 Exportation du fer par la ferroportine ...18
3.6 Régulation de l’homéostasie cellulaire du fer par le système IRE/IRP ...18
4. Homéostasie systémique du fer ...21
4.1 Distribution corporelle du fer...21
4.2 Erythrophagocytose et recyclage du fer héminique par les macrophages...23
4.2.1 Mécanismes de l’érythrophagocytose et du recyclage du fer...23
4.3 Absorption intestinale du fer ...25
4.3.1 Absorption du fer non héminique...26
4.4 Régulation systémique du fer...28
5. Hepcidine ...31
5.1 Découverte de l’hepcidine comme étant un peptide antimicrobien ...31
5.2 Structure de l’hepcidine ...32
5.3 Sites d’expression de l’hepcidine...33
5.5 Mécanisme d’action de l’hepcidine...36
5.5.1 Axe fonctionnel hepcidine-ferroportine ...36
5.5.2 Interaction moléculaire entre l’hepcidine et la ferroportine ...37
5.6 Régulation de l’expression hépatique d’hepcidine...39
5.6.1 Régulation d’hepcidine par le signal des réserves en fer...39
5.6.2 Régulation d’hepcidine par le signal inflammatoire...41
5.6.3 Régulation d’hepcidine par l'anémie, l'érythropoïèse et l'hypoxie ...42
5.7 Pathologies liées à l’hepcidine ...44
5.7.1 Hémochromatose héréditaire...44
5.7.2 Anémie des maladies inflammatoires chroniques (ACD) ...44
6. Interaction entre le système immunitaire et le fer...45
6.1 Homéostasie du fer et immunité adaptative ...45
6.2 Homéostasie du fer et immunité innée...46
7. Les récepteurs Toll-like ...49
7.1 Localisation, structure et ligands des TLRs ...49
7.1.1 Localisation des TLRs ...49
7.1.2 Structure des TLRs ...51
7.1.3 Ligands des TLRs...53
7.2 Modulation de la réponse immunitaire par le TLRs ...55
7.3 Voies de signalisation des TLRs ...58
8. Rôle des TLRs dans l’expression d’hepcidine et le développement des maladies inflammatoires ...61
II. OBJECTIFS ET BUTS SPÉCIFIQUES ...63
III. MATÉRIELS ET MÉTHODES ...67
IV. RÉSULTATS ...74
V. DISCUSSION ET CONCLUSION ...96
VI. PERSPECTIVES...103
Liste des figures
Figure 1. Structure des clusters fer soufre. ...5
Figure 2. Détoxification de l’oxygène et réaction de Fenton. ...10
Figure 3. Capture du fer via la voie dépendante de la transferrine...14
Figure 4. Régulation du métabolisme cellulaire du fer par le système IRE/IRP...20
Figure 5. Distribution corporelle du fer chez l’adulte. ...22
Figure 6. Erythrophagocytose et de recyclage du fer héminique ...24
Figure 7. Absorption intestinale du fer alimentaire par les entérocytes. ...27
Figure 8. Séquences de l’hepcidine chez les mammifères et les poissons.. ...34
Figure 9. Structure de l’hepcidine humain. ...34
Figure 10. Axe fonctionnel hepcidine-ferroportine dans le contrôle du fer. ...38
Figure 11. Régulation de l’expression hépatique d’hepcidine. ...43
Figure 12. Mécanismes de rétention du fer durant les infections...48
Figure 13. Les récepteurs Toll-like et leurs ligands. ...50
Figure 14. Structure des TLRs...52
Figure 15. Modulation de la réponse immunitaire par les récepteurs Toll-Like. ...57
Figure 16. Les voies de signalisation induites par les TLRs. ...60
Figure 17. Expression de l'hepcidine dans les macrophages RAW 264.7...76
Figure 18. Etude cinétique de l’expression de l'hepcidine dans les RAW 264.7. ...77
Figure 19. Etude cinétique de l’expression de la ferroportine dans les RAW 264.7...78
Figure 20. Marquage de surface du F4/80 et analyse par FACS...79
Figure 21. Expression de l'hepcidine dans les MPMs des souris sauvages...80
Figure 22. Expression de l'hepcidine dans les MPMs TLR2-/-...81
Figure 23. Expression de l'hepcidine dans les MPMs MyD88-/-. ...82
Figure 24. Expression de l'hepcidine hépatique in vivo. ...84
Figure 25. Taux du fer sérique...85
Figure 26. Saturation de la transferrine. ...86
Figure 27.Taux sérique de TNF-alpha...87
Figure 28. Taux sérique d’interleukine 6...88
Figure 29. Etude cinétique de l’expression de l'hepcidine hépatique...89
Figure 30. Etude cinétique de l’expression de la ferroportine hépatique par qRT-PCR. 90 Figure 31. Etude cinétique de l’expression protéique de FPN1 hépatique par WB. ...91
Figure 32. Quantification de l’expression protéique de FPN1.. ...91
Figure 33. Taux du fer sérique...92
Figure 34. Saturation de la transferrine. ...93
À ma famille qui m’a soutenu tout au long de mes études
Remerciements
Nombreux sont ceux qui m’ont aidé, encouragé, réconforté, soulagé, tout au long de ma maitrise et je ne saurais leur exprimer mes remerciements autant que je le souhaiterais; leurs discussions, leurs conseils, leur présence, me furent éminemment précieux. En écrivant leur nom sous la forme du cèdre de mon pays le Liban, je suis gré de la disponibilité, de la patience, et de la bienveillance de :
Dre Manuela Santos
Dre Marika Sarfati Dr Janos G. Filep Dr Edward Bagu Dr Rafik Terra Alexandre Reuben Jean-Baptiste Loubert
Annie Calvé Hua Huang Xuehai Wang
Zenghui Wu
INTRODUCTION
Le fer est un oligoélément nécessaire pour le fonctionnement normal de toutes les cellules de l’organisme. Chez l'Homme, bien qu’il soit présent en très faible quantité, le fer joue un rôle essentiel dans de nombreuses fonctions biologiques. C’est un composant essentiel des protéines et d’enzymes impliquées dans des processus métaboliques tels que le transport de l'oxygène, la synthèse d'ADN et la production d’énergie. Le fer est également essentiel pour la régulation de la croissance et la différenciation cellulaires. Cependant, le taux du fer dans l’organisme doit être bien régulé, sinon la carence en fer entraîne divers états pathologiques comme l’anémie et la diminution de l'immunité. D’autre part, la surcharge en fer potentialise la disposition des microbes et aggrave les infections, ainsi que la formation des radicaux libres ayant des effets toxiques sur les cellules et leurs composants, favorisant ainsi des pathologies cardiovasculaires, inflammatoires et des cancéreuses.
1.
Propriétés chimiques et fonctions biologiques du fer
Le fer est un composant de plusieurs métalloprotéines et joue un rôle crucial dans les activités biochimiques vitales, telles que la détection et le transport d'oxygène, le transfert d'électrons et la catalyse [1].
Dans tous les organismes, le fer est impliqué dans un large éventail de fonctions métaboliques, mais sa biodisponibilité est généralement limitée, car dans des conditions d’aérobie, le fer ferreux (Fe2+) est rapidement oxydé en fer ferrique (Fe3+) qui est pratiquement insoluble à pH physiologique [2].
Le fer a un potentiel redox en tant que métal de transition où il subit des réactions d’oxydation-réduction entre ses états ferreux et ferrique. Cette capacité de pouvoir accepter ou donner alternativement un électron fait du fer le cofacteur de choix de nombreuses réactions enzymatiques qui sont essentielles pour le métabolisme cellulaire et le fonctionnement de nombreuses enzymes cellulaires [1-2].
L’importance biologique du fer est largement attribuable à ses propriétés chimiques, par exemple sa capacité de former une variété de coordination complexe avec des ligands organiques, et cela dans un mode flexible et un environnement dynamique [2].
En général, trois types de réactions sont relatifs pour la plupart des fonctions biologiques du fer, chez tous les organismes: L’oxydoréduction, l'hydrolyse et la formation de complexes polynucléaires [1].
Les fonctions biologiques du fer sont la conséquence de sa liaison à de nombreuses protéines qui peuvent être classées en 3 catégories :
• Les hémoprotéines • Les protéines fer-soufre
• Autres protéines contenant du fer et protéines liant le fer 1.1 Les hémoprotéines
Une fraction importante du fer est associée à des protéines sous la forme d’un complexe appelé hème, un groupement fonctionnel des hémoprotéines qui sont présentes dans tous les tissus [3]. L’hème sert de prothèse pour des protéines impliquées dans le transport de l'oxygène, la respiration mitochondriale, le métabolisme des médicaments, la biosynthèse des stéroïdes, les défenses cellulaires antioxydantes et les processus de transduction du signal. Cette molécule est impliquée dans une variété d'événements biologiques en modulant la fonction ou bien l'état des hémoprotéines qui assurent différentes fonctions [4].
Parmi les hémoprotéines on distingue :
a) L’hémoglobine et la myoglobine : Impliquées respectivement dans le transport d’oxygène par les érythrocytes et les muscles [2].
b) Les cytochromes : Impliquées dans le transfert des électrons et dans le métabolisme énergétique mitochondrial. D’autres cytochromes, comme ceux du réticulum endoplasmique, sont impliquées notamment dans le métabolisme des acides gras [5].
c) Les oxydases, catalases et peroxydases : Impliquées dans le processus de détoxification des espèces réactives de l’oxygène (ROS) [6].
1.2 Les protéines fer-soufre
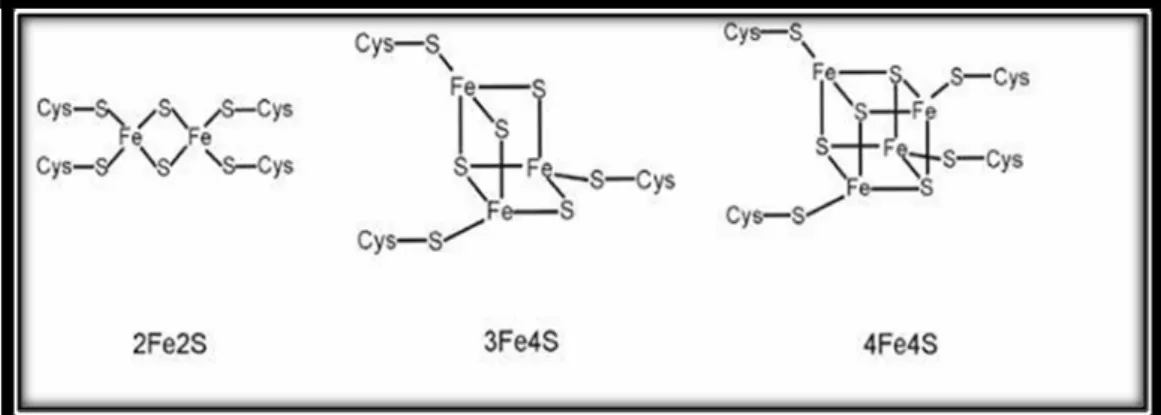
Dans les protéines fer-soufre, les atomes du fer sont assemblés avec les atomes du soufre dans différents états d’oxydation, tels que le 2Fe-2S, le 3Fe-4S ou le 4Fe-4S formant une structure appelée cluster [7], (Figure 1). Ces clusters fer-soufre (Fe-S) sont présents dans plus de 200 différents types d'enzymes ou de protéines et constituent l'une des plus anciennes structures des molécules organiques. Ainsi, le processus de biosynthèse des clusters Fe-S est essentiel pour presque toutes les formes de vie et il est remarquablement conservé chez les organismes procaryotes et eucaryotes [8]. Ces clusters sont présents dans toutes les cellules vivantes où ils participent à divers processus physiologiques, y compris la respiration, la photosynthèse, la réparation d’ADN, le métabolisme et la réglementation de l'expression génique [9]. Les clusters Fe-S sont reconnus depuis longtemps comme cofacteurs essentiels et polyvalents des protéines, par exemple la rubrédoxine et la ferrédoxine qui interviennent dans la catalyse et le transfert des électrons dans une gamme de réactions métaboliques [10].
Figure 1. Structure des clusters fer soufre.
Les atomes du fer sont assemblés avec les atomes du soufre dans différents états d’oxydation, tels que 2Fe-2S, 3Fe-4S et 4Fe-4S formant une structure appelée cluster. Adapté de [10].
1.3 Autres protéines contenant du fer
D’autres protéines peuvent se lier directement à l’atome du fer, telles que les lipoxygénases, la lactoferrine et les ribonucléotides réductases :
a) Les lipoxygénases :
Les lipoxygénases (LOXs) sont une famille du fer non hémique contenant des dioxygénases qui catalysent l'oxydation des acides gras polyinsaturés [11]. La formation d'hydroperoxydes d'acides gras peut se produire soit par oxydation chimique ou bien par l'action d'enzymes, telles que les lipoxygénases [12]. Ainsi, ces acides gras peuvent être ensuite métabolisés en aldéhydes volatiles et en acide jasmonique chez les végétaux [13], des lactones chez les champignons [14], et en lipoxines et leucotriènes chez les mammifères [15]. Dans les plantes et les mammifères, la synthèse des LOXs se fait d’une façon ubiquitaire et récemment, elles ont été détectées dans les champignons et un certain nombre de bactéries [16]. Ces molécules jouent un rôle important dans la cicatrisation des plaies et les processus de défense chez les végétaux, tandis que chez les mammifères ils sont impliqués dans l'inflammation, l'asthme et les cardiopathies. Chez les champignons, ils jouent un rôle dans la régulation de la production des mycotoxines et du cycle de vie sexuée et asexuée [14]. À noter que jusqu’à présent, rien n’est connu sur la fonction biologique de ces enzymes chez les procaryotes.
b) La lactoferrine :
La lactoferrine (Lf) est une glycoprotéine de la famille des transferrines. Elle est exprimée et sécrétée par les cellules glandulaires et se trouve dans les granules secondaires des neutrophiles à partir de laquelle elle est libérée dans les tissus infectés et dans le sang au cours du processus inflammatoire. La Lf est connue pour être une protéine multifonctionnelle ou multi-tâches. C'est une composante importante du système immunitaire inné des mammifères.
Ses effets vont de la protection anti-microbienne directe des activités contre un large panoplie de micro-organismes comme les bactéries, virus, champignons et des parasites, à des activités anticancéreuses et anti-inflammatoires. Alors que la chélation du fer est au centre de certaines des fonctions biologiques, d'autres activités impliquent des interactions de la Lf avec des composants moléculaires et cellulaires de l’hôte et des agents pathogènes. Sa puissante activité antimicrobienne, les propriétés immunomodulatrices, ainsi que ses fonctions anti-cancérigènes et son importance croissante dans la prestation du fer et la croissance osseuse, font de cette molécule et ses dérivés des outils importants pour la santé [17].
c) Les ribonucléotides réductases :
Les ribonucléotides réductases (RNRs), également connues sous le nom de ribonucléosides diphosphates réductases, sont les seuls enzymes responsables de la conversion des nucléotides en déoxynucléotides dans toutes les cellules en division, et cela pour la synthèse de l'ADN [18]. La réaction catalysée par les RNRs est strictement conservée dans tous les organismes vivants. En outre, RNR joue un rôle critique dans la régulation du taux total de la synthèse d'ADN, de sorte que la quantité de l'ADN cellulaire est maintenue à un taux constant pendant sa réparation et durant la division cellulaire [19].
1.4 Protéines liant le fer a) La transferrine :
La transferrine (Tf) ou sidérophiline est une globuline monomérique synthétisée par le foie, constituée d'une seule chaîne polypeptidique porteuse de 2 sites de captation du fer, à raison de 2 atomes du fer par molécule de transferrine. Le rôle fondamental de la transferrine est de contrôler les niveaux du fer dans l’organisme en liant, séquestrant et transportant les ions Fe3+ ferrique, ce qui contribuera à maintenir la biodisponibilité du fer en empêchant en même temps le dépôt d'agrégats insolubles d'hydroxyde ferrique [20]. Dans des conditions physiologiques normales, la capacité de Tf à lier le fer inorganique est très forte et sa saturation est à l’entour de 35%, ce qui suggère qu'il existe une capacité suffisante pour empêcher la libération du fer toxique dans la circulation qui endommage les cellules en catalysant la formation de radicaux libres [21]. En outre, la Tf est susceptible d'être impliquée dans le transport d'un large éventail d'ions de métaux autres que le fer. En plus, la Tf a une activité antimicrobienne qui repose apparemment sur le contact réel avec les bactéries plutôt que la privation du fer. Cependant, la Tf peut aider à la prolifération rapide des cellules tumorales, et cela par la translocation du fer aux sites tumoraux [20].
b) La ferritine :
La ferritine, ou hémosidérine, est une protéine chargée du stockage du fer au sein des cellules à l’état soluble et non-toxique. D’autre part, la ferritine peut libérer une grande quantité du fer quand le corps est en carence. C’est une protéine cytoplasmique composé de 25 sous-unités hétérodimériques de chaînes légères (L-ferritin) et lourdes (H-(L-ferritin) pouvant stocker jusqu'à 4500 ions Fe3+ [22]. Enfin, la ferritine est présente dans le foie, le cœur, la rate et la moelle osseuse et une quantité infime se trouve dans le sang sous forme de ferritine sérique [21].
2. Toxicité du fer
L'efficacité du Fer ferreux en tant que donneur d'électrons et celle du Fer ferrique comme accepteur d'électrons, avec un potentiel d'oxydoréduction compatible avec les contraintes de l'environnement cellulaire, est une caractéristique fondamentale pour de nombreuses réactions biochimiques, ce qui rend le fer un minéral et un nutriment essentiel [2]. Cependant, comme le corps n'a pas de voie active pour l'excrétion du fer, une charge continue supérieure à 1-2 mg/jour se traduira par une surcharge en fer et provoquera des dommages aux organes comme le foie, le cœur, le pancréas, la thyroïde et le système nerveux central [21].
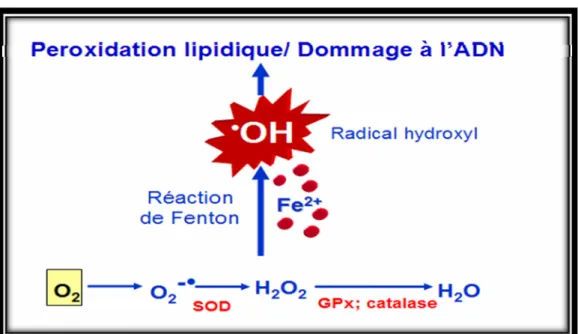
La principale cause de ces dommages d’organes est due à la production d'espèces réactives de l'oxygène (ROS) qui est largement basée sur les réactions chimiques de Fenton et Haber-Weiss où des quantités catalytiques du fer sont suffisantes pour produire des radicaux hydroxyles (OH-), de superoxyde (O2 - .) et des peroxydes
d'hydrogène (H2O2) [23]. Dans les conditions physiologiques normales, le fer
extracellulaire est exclusivement lié à la transferrine qui maintient le fer soluble et non toxique incapable de s'engager dans les réactions de Fenton et Haber-Weiss. Tandis qu’une petite portion de l’oxygène utilisé par la mitochondrie est réduite pour former des superoxydes qui seront convertis en eau (H2O) par la superoxide
dismutase (SOD), la catalase (CAT) et la gluthatione peroxydase (GPx) (Figure 2). Les radicaux libres sont des espèces hautement réactives et peuvent favoriser l'oxydation des protéines, la peroxydation des lipides membranaires et la modification des acides nucléiques. Parmi les ROS, le radical hydroxyle est la fraction la plus toxique et s'adresse aux glucides, aux protéines et aux acides nucléiques. Il est connu que la réaction des radicaux hydroxyles avec la base de l'acide nucléique 8-hydroxyguanine (8-OHG) est fortement corrélée à la tératogénicité et la cancérogénicité par des stress oxydatifs. Un autre ROS puissant montrant une réactivité similaire à celle du radical hydroxyle c’est l’hydroxy- peroxyde des lipides (ROOH).
Dans la surcharge en fer, les produits lipidiques tels que le peroxidative malondialdéhyde et les 4-hydroxy-2-nonenal sont augmentés, cela va entraîner la formation d’alkyl oxyradical (ROO-) et d’alcoxy radical (RO-). Ces radicaux lipidiques possèdent une demi-vie plus longue que les radicaux hydroxyles et ont également une plus forte capacité pour la toxicité chronique des cellules et les dommages de l'ADN [21].
Figure 2. Détoxification de l’oxygène et réaction de Fenton.
L’oxygène utilisé par la mitochondrie est réduit pour former des superoxydes qui seront convertis en eau (H2O) par la superoxide dismutase (SOD), la catalase (CAT)
et la gluthatione peroxydase (GPx). Dans des conditions physiologiques, le fer est lié à la transferrine qui maintient le fer soluble et non toxique incapable de s'engager dans les réactions de Fenton. Tandis que dans le cas d’un excès en fer ferreux (Fe2+), ce dernier va s’engager dans la réaction de Fenton et entrainer la
3. Homéostasie cellulaire du fer
L’homéostasie du fer doit être assurée à la fois au niveau cellulaire et par l’organisme dans son ensemble. En ce qui concerne l’homéostasie cellulaire, cette dernière doit être strictement contrôlée au niveau de la captation, du stockage et de l'exportation du fer.
3.1 Captation du fer
Comme le fer est un cofacteur essentiel pour un large éventail d’importants processus cellulaires, sa captation par divers types de cellules est nécessaire. On distingue trois mécanismes de séquestrations du fer par les cellules :
- Mécanismes dépendants de la Tf et utilisés par presque toutes les cellules. - Mécanismes spécifiques observés dans les macrophages et les entérocytes. - Mécanismes indépendants de la Tf.
3.2 Capture cellulaire du fer par des mécanismes dépendants de la Transferrine 3.2.1 Récepteur de la transferrine (TfR)
Le TfR1 joue un rôle majeur dans l'absorption du fer cellulaire en se liant et internalisant le fer lié à la Tf (holotransferrine). Il appartient au groupe des récepteurs des membranes plasmiques présentant un cycle endocytose-recyclage qui s'amorce avec la fixation de leur ligand. Le TfR1 est une glycoprotéine dimerique faite de deux chaînes identiques ayant une masse moléculaire de 84,91 Da [24, 25]. Ces deux chaînes sont en situation parallèle et solidarisées par deux ponts disulfures établis entre les deux résidus 89 et les deux résidus 98 [26].
Chacune des deux chaînes comporte 760 résidus d'acides amines distribués en trois domaines fonctionnels [24, 25]:
- Un domaine cytosolique aminoterminal de 61 acides aminés, directement impliqué dans le déclenchement de l'internalisation du récepteur chargé de son ligand [27].
- Un domaine transmembranaire de 28 acides aminés assurant l'ancrage du récepteur dans la membrane plasmique [28, 29].
- Un domaine extracellulaire carboxyterminal comportant 671 acides aminés, qui fixe la Tf avec une affinité beaucoup plus grande lorsque celle ci est chargée de ses deux atomes du fer [28[30].
La plupart des cellules des mammifères expriment à la surface de leurs membranes des récepteurs pour la transferrine. L'expression la plus large a lieu sur les cellules nécessitant un apport important et continu en fer comme les précurseurs médullaires de la lignée érythrocytaire. Il est noté que le nombre de récepteurs augmente au cours de la maturation cellulaire, ainsi que dans le foie et les cellules en division rapide normales et malignes [31].
Récemment, un homologue proche du récepteur de TfR1 a été cloné : c’est le récepteur de la transferrine 2 (TfR2). Ce dernier présente 45% d’homologie avec TfR1 dans le domaine extracellulaire, et il est capable de lier la Tf pour fournir le fer aux cellules, mais cela avec une affinité 25 fois plus faible, suggérant que la capture du fer n’est pas la principale fonction de ce récepteur. Toutefois, la distribution du TfR2 diffère de celle du TfR1, l'expression la plus large a lieu sur les hépatocytes en différenciation et beaucoup moins sur les érythroblastes [31]. D’autre part, TfR2 n'était pas en mesure de compenser la fonction du TfR1 dans les souris « knock-out » qui meurent dans l’utérus [32]. Ainsi que des mutations au niveau du gène de ce récepteur ont été identifiées chez des patients italiens qui ont développé une surcharge en fer hépatique, indiquant qu'en plus de sa capacité de transporter le fer dans les cellules, TfR2 est également important dans le maintien de l'homéostasie du fer [33].
3.2.2 Cycle de la transferrine
En condition normale, le fer est majoritairement transféré vers le compartiment médullaire où il est nécessaire à la synthèse de l’hémoglobine. La fraction du fer non orientée vers ce compartiment est partagée entre les divers autres sites d’utilisation et les sites de stockage représentés surtout par les hépatocytes particulièrement sensibles aux surcharges en fer [34].
Le fer est principalement transporté dans le plasma sous forme de Fe3+ lié à la Tf. Le complexe Tf-Fe3+, nommé holotransferrine, est ensuite capté par TfR1 présent
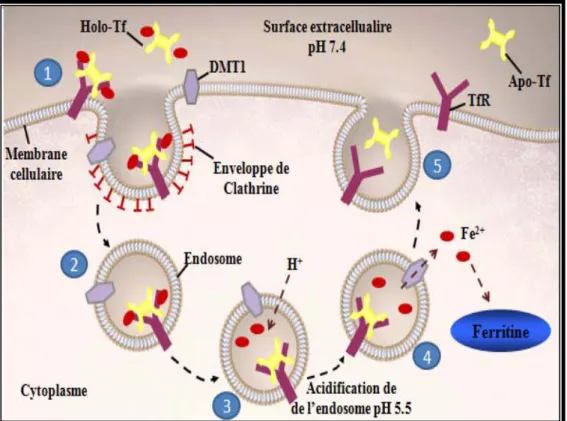
au niveau des différents organes. Le modèle d’internalisation du fer lié à la transferrine par le TfR1 recycle le complexe transferrine-récepteur et s’appelle le « transferrin-to-cell cycle » [35] (Figure 3).
Ce cycle débute par la liaison de la Tf ferrique au TfR1 à la surface cellulaire par une interaction physico-chimiques, ne nécessitant ni de température physiologiques ni d'énergie métabolique [23]. Le complexe Tf-Fe3+-TfR1, par un processus énergétiquement dépendants, est internalisé par endocytose suite à la formation de puits tapissés de clathrines qui s’invaginent pour former un endosome [36]. L’endosome formé subit une protonation ou bien une acidification progressive qui est ATPase-dépendante, ce qui entraîne la libération du Fe3+ de la Tf, et cela dans un pH acide d’environ 5,5. Malgré que le pH minimum dans les endosomes ne soit pas suffisant pour libérer le fer de ces deux sites de liaison dans la Tf, les cellules sont capables de le faire avec une capacité remarquable. En fait, des études ont montrés que le TfR1 joue un rôle non seulement par sa liaison à la Tf ferrique, mais aussi dans l'accélération de la dissociation du Fe3+ de la Tf dans de l’endosome [37,
38]. Ensuite, le Fe3+ est réduit en Fe2+ possiblement par la ferriréductase Steap3 « six-transmembrane epithelial antigen of the prostate 3 » et est transporté par le « divalent metal transporter 1 » (DMT1) à travers la membrane endosomale dans le cytoplasme [39, 40].
Au pH acide de l’endosome, l’apotransferrine (Tf sans fer) a une forte affinité pour le récepteur, pour cela elle demeure liée au TfR1 et le complexe est recyclé vers la surface cellulaire. À la surface cellulaire et dans un pH physiologique, l’affinité de l’apotransferrine pour le récepteur est perdue, ce qui entraîne sa séparation de TfR1 qui sera alors libre de lier d’autre Tf diferrique [41]. Si le TfR1 n’est pas lié à la transferrine diferrique, il n’est pas endocyté et passe dans la circulation sous forme de TfR1 soluble (sTfR1) grâce à des protéases membranaires [42].
Figure 3. Capture du fer via la voie dépendante de la transferrine.
Le complexe fer-transferrine (holotransferrine) est capté par le récepteur de la transferrine (TfR). Ensuite, il est inclus après invagination de la membrane dans une vésicule d’endocytose incluant le « divalent metal transporter 1 » (DMT1) qui assure la pénétration du fer dans le cytoplasme et ainsi la transferrine perds le fer pour devenir apotransferrine (Apo-Tf, Tf sans fer). Après passage du fer dans le cytosol, la vésicule d’endocytose s’ouvre en se fusionnant avec la membrane et le complexe Apo-Tf et TfR1 est recyclé vers la surface cellulaire libérant l’Apo-Tf transferrine dans la circulation générale. Adapté de [34].
3.3 Capture cellulaire du fer par des mécanismes indépendants de la Tf
Dans les conditions physiologiques, la Tf est la principale source du fer pour la plupart des cellules de l'organisme [43]. Cependant, dans divers états pathologiques de surcharge en fer, comme l’hémochromatose et la thalassémie, la quantité de cet ion dans la circulation dépasse la capacité de liaison à la transferrine, conduisant à l'apparition d’une forme du fer très réactif, c’est le fer non lié à la Tf (NTBI) « non-transferrin-bound iron » [44, 45].
Malgré les investigations, la détermination de la forme chimique de NTBI s'est avérée difficile à définir. Des études suggèrent qu’il est retrouvé sous la forme de complexe citrate-fer ou bien albumine-fer. Vraisemblablement, NTBI est présent principalement sous la forme Fe3+ [46]. Dans des circonstances normales, la Tf à une affinité exceptionnellement élevée pour le fer, ainsi la quantité de NTBI dans le plasma est extrêmement faible [47].
Auparavant, on croyait que la Tf était le seul moyen pour délivrer le fer aux cellules. Tandis que chez les humains ayant une maladie congénitale touchant la Tf, comme l’atransferrinémie (absence de la Tf) ou l’hypotransferrinémie (déficit en Tf), le fer est rapidement délivré aux tissus, et cela est accompagné par des états de surcharge en fer [48-50]. Mêmes observations ont été validées chez des souris dont le gène codant la Tf a été supprimé [51]. Ces observations appuient donc la présence des moyens de captation du fer cellulaire par des mécanismes indépendants de la Tf. Jusqu'à présent, il reste à clarifier comment NTBI fait son chemin à travers la membrane plasmique pour renter dans les cellules, mais plusieurs candidats ont été proposés :
a) Ferritine : En plus de la voie Tf-TfR, les hépatocytes et les cellules érythroïdes précurseurs peuvent également capturer le fer via la ferritine [52].
b) DMT1: En tant que transporteur membranaire majeur du fer, les souris dépourvues de DMT1 peuvent encore accumuler le fer au niveau du foie, suggérant que DMT1 n'est pas la seule voie d'entrée de NTBI [53].
c) Lipocaline 2 (LCN2): Connue sous le nom de lipocaline associée au gélatinase de neutrophile, c’est un petit peptide antimicrobien qui lie les sidérophores (de petites molécules synthétisées par les bactéries et les champignons) qui ont une haute affinité pour le fer. Lipocaline 2 transporte les complexes sidérophore-fer dans des cellules [54]. Malgré sa capacité de lier le fer libre dans la circulation, une étude récente faite dans notre laboratoire a montré que lipocaline 2 n’est pas essentielle dans le transport du fer dans le foie durant l’hémochromatose [55].
d) L-Type Ca2+ : Les canaux Ca2+de type L est une voie importante car elle permet l’entrée de NTBI au niveau du cœur. Ainsi, dans des conditions de surcharge en fer, cette entrée mène à une accumulation du fer au niveau de cet organe et engendre éventuellement une cardiomyopathie [56].
e) SFT: Stimulateur de transport du Fe. Cette protéine a été isolée des cellules érythroïdes des leucémies, en tant que transporteur du fer. Il semble jouer un rôle dans les mécanismes d’absorptions dépendantes et indépendantes de la transferrine, mais son rôle reste mal caractérisé [57, 58].
f) Zip14: Le transporteur de Zinc Zip14 pourrait être impliqué dans la capture du NTBI. Il a été montre qu’il facilite l’absorption de NTBI dans les hépatocytes [59].
3.4 Devenir du fer cellulaire
Après avoir pénétré dans la cellule, le fer doit être correctement réparti entre trois pools différents représentés par le pool de transit, le pool fonctionnel et le pool de stockage [60].
a) Le pool de transit :
Encore appelé pool du fer de bas poids moléculaire ou bien pool du fer labile (LIP). À partir de ce dernier le fer est adressé soit vers le pool fonctionnel, soit vers le pool de stockage. Il s’agit du fer présent dans le cytosol lié à des espèces chimiques de bas poids moléculaire [61]. Les niveaux de LIP reflètent le statut cellulaire et ainsi, des augmentations ou des baisses dans le LIP, activent des réponses liées à une déficience ou à une surcharge en fer [6].
b) Le pool fonctionnel :
Le pool fonctionnel est constitué par la quantité du fer nécessaire et suffisante pour assurer la survie et les fonctions respiratoires de la cellule, ainsi que les communications intercellulaires. Il s’agit du fer incorporé dans les protéines héminiques dont l’hémoglobine et les cytochromes et aussi du fer cofacteur de multiples protéines contrôlant le cycle cellulaire, comme : p53 « Tumor protein 53 », GADD45 « Growth Arrest and DNA-Damage Inducible 45 Gene », WAF1/p21 « cyclin-dependent kinase inhibitor » [62].
c) Le pool de stockage :
Le pool de stockage est dans sa grande majorité représenté par le fer lié à la ferritine. À tout moment, ce fer pourra être libéré pour gagner les sites d’utilisation et cela en fonction des besoins.
3.5 Exportation du fer par la ferroportine
La ferroportine (FPN1) est connue comme étant l’exporteur unique du fer cellulaire chez les vertébrés [63-65]. La ferroportine se trouve dans tous les tissus impliqués dans le métabolisme du fer, tels que les entérocytes duodénaux, les macrophages et les hépatocytes.
Des études chez des souris « knock-out » ont montré que la perte totale de l’expression du ferroportine est létale au stade embryonnaire, en raison de l'incapacité de transférer le fer de la mère à l'embryon [66]. En effet, chez des souris « knock-out » , où l’expression de la ferroportine est préservée durant l'interface materno-fœtale, les nouveaux nés présentaient une carence sévère en fer, causée par une déficience dans l’absorption intestinale du fer alimentaire, une diminution de la libération du fer hépatique, ainsi qu’une réduction de la libération du fer par les macrophages [66]. Ces études suggèrent que chez les mammifères, la ferroportine est le seul exporteur significatif du fer dans les tissus impliqués dans l'absorption, le recyclage et le stockage du fer.
Parmi les nombreux mécanismes qui concourent au maintien de l’homéostasie cellulaire du fer, un mécanisme post-transcriptionnel des gènes liés au métabolisme du fer semble particulièrement important : le système IRE/IRP.
3.6 Régulation de l’homéostasie cellulaire du fer par le système IRE/IRP
L'homéostasie cellulaire du fer est assurée par un contrôle de l’expression du récepteur de la transferrine et de la ferritine, qui régulent respectivement l'absorption et le stockage du fer. La synthèse du TfR1 et de la ferritine est réciproquement contrôlée au niveau post-transcriptionnel, et cela en réponse à la concentration du fer intracellulaire [67].
Les ARNm codant le TfR1 et la ferritine contiennent au niveau de leurs régions 5’ ou 3’ non traduites (UTR) « untranslated regions », des motifs structuraux appelés éléments de réponse au fer (IREs) « iron responsive elements » [68].
Les IREs sont des séquences nucléotidiques particulières avec des structures secondaires en épingle à cheveux [69]. Les ARNm de la ferritine possèdent une séquence IRE dans leur région 5’UTR, tandis que les ARNm codant le TfR1 possèdent cinq séquences IRE dans leur région 3’UTR [70, 71].
Les IREs sont la cible de deux protéines cytoplasmiques régulatrices du fer (IRP1 et IRP2) « iron regulatory proteins 1 and 2 », qui peuvent lier la séquence IRE avec une haute affinité. L’incidence de la liaison IRP-IRE est différente selon que la séquence IRE est localisée en 5’ ou bien en 3’ UTR. En effet, la liaison de l’IRP à l’IRE en région 5’UTR bloque la traduction de l’ARNm considéré, alors que sa liaison à l’IRE en région 3’UTR empêche la dégradation de cette ARNm et permet donc sa traduction [72].
Dans le cas de carence en fer cellulaire, IRP1 et IRP2 vont se lier sur l’IRE en 5’UTR du mRNA de la ferritine et entraîner sa dégradation, empêchant ainsi le stockage du fer en excès, d’autre part IRP1 et IRP2 vont se lier sur l’IRE en 3’UTR de l’ARNm de la TfR et augmenter sa traduction, augmentant ainsi l’entrée du fer dans la cellule [73, 74].
Par contre, lorsque la concentration cellulaire en fer est élevée, les IRPs ne peuvent pas s’y associer aux IRE des ARNm [75] (Figure 4). L’IRP1 va s’associer avec un cluster 4Fe-4S, ce qui l’empêche de se lier à l’IREs des ARNm du TfR1 et de la ferritine, tandis que l’IRP2 est dégradée par ubiquitination [76].
Aussi, les ARNm de DMT1 et de la Tf possèdent respectivement une et cinq séquences IRE dans leur région 3’UTR, ainsi que la Fpn qui a un IRE dans son extrémité 5’UTR [77].
Cependant, le rôle des IREs dans le contrôle de l’expression de la Tf, du FPN1 et de DMT1 par les concentrations intracellulaires en fer n’est pas encore bien élucidé. Enfin, le système IRP/IRE peut être influencé par des signaux indépendants du fer, tels que les niveaux du peroxyde d'hydrogène et de l'oxyde nitrique, ainsi que par l'hypoxie [74].
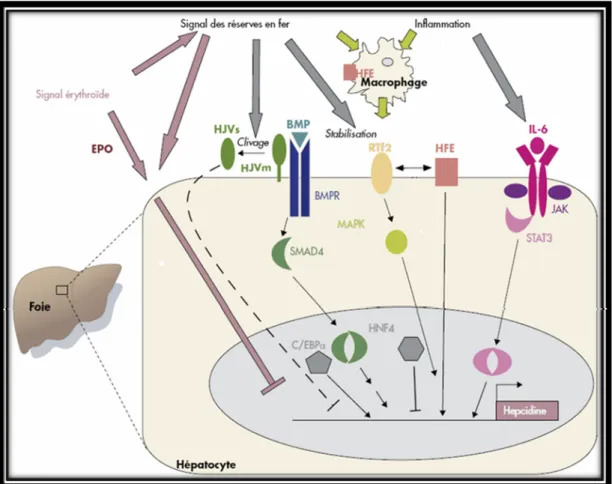
Figure 4. Régulation du métabolisme cellulaire du fer par le système IRE/IRP. Lorsque la cellule est déficiente en fer, IRP1 et IRP2 vont se lier sur l’IRE en 5’UTR du mRNA de la ferritine et entraîner sa dégradation, empêchant ainsi le stockage du fer en excès, d’autre part IRP1 et IRP2 vont se lier sur l’IRE en 3’UTR de l’ARNm de la TfR et augmenter sa traduction, augmentant ainsi l’entrée du fer dans la cellule. Par contre, lorsque la concentration cellulaire en fer est élevée, les IRPs ne peuvent pas s’y associer aux IRE des ARNm, ce qui augmente l’expression de la ferritine et diminue celle du TfR. Adapté de [78].
4. Homéostasie systémique du fer
Le fer est un véritable paradoxe étant d’une part indispensable à notre organisme et pouvant de l’autre lui être nocif. Comme les mammifères ne possèdent pas de voie physiologique efficace pour l'excrétion du fer, la régulation de cet ion métallique s’avère tout à fait nécessaire. L'homéostasie systémique de l’absorption et du recyclage du fer est réglementée notamment au niveau de son absorption d’une façon à empêcher aussi bien un trop peu qu’un trop du fer [34].
4.1 Distribution corporelle du fer
Afin de comprendre comment l'homéostasie systémique du fer est maintenue, il est prudent d'examiner le mouvement de cet ion entre les différents compartiments fonctionnels du corps. L’organisme d’un Homme adulte contient environ 4 g du fer total [79]. Dans des conditions normales, 1-2 mg du fer sont absorbés quotidiennement via les entérocytes des villosités du duodénum et de la partie proximale du jéjunum. Ce fer alimentaire absorbé est ensuite libéré dans la circulation où il va être lié à la Tf sérique. Environ 3 mg du fer-Tf sont présents dans la circulation sanguine. Bien que cela ne représente qu'une faible proportion du fer corporel total, ce compartiment est vital pour le maintien de l'homéostasie. En fait, ce fer est utilisé par la moelle osseuse à un rythme approximatif de 22 mg par jour pour la synthèse de l'hémoglobine des globules rouges durant l’érythropoïèse [79]. Dans le corps humain, 80% du fer est incorporé dans l’hémoglobine des globules rouges circulants [60]. Cependant, les globules rouges sénescentes sont éliminés de la circulation par les macrophages de système réticulo-endothélial où le fer sera libéré de l'hémoglobine et sera ou bien stocké dans la protéine de stockage la ferritine, ou bien libéré dans la circulation pour lier la Tf. Les macrophages recyclent à l’entour de 22 mg du fer par jour, remplaçant ainsi le montant repris par la moelle osseuse (Figure 5).
Pour des quantités absolument infimes, d’autres cellules de l’organisme absorbent le fer en utilisant la voie du TfR1. Ce fer va être utilisé pour la synthèse de différentes protéines intracellulaires contenant du fer, telles que l'hème, les cytochromes et les clusters fer-soufre. Approximativement, 10 à 15% du fer est présent dans telles protéines, avec 80% dans les myoglobines des cellules musculaires et le 20% restant est stocké dans la ferritine, principalement dans les macrophages et les hépatocytes du foie [79].
Figure 5. Distribution corporelle du fer chez l’adulte.
Le fer absorbé au niveau du l’intestin se lie à transferrine (TF) pour être distribué vers son plus grand consommateur la moelle osseuse et les globules rouges circulantes. D’autre part, les macrophages tissulaires phagocytent les érythrocytes sénescents et recyclent le fer contenu dans l’hème. Enfin, le fer est stocké dans les hépatocytes et retourne vers la circulation en cas de besoin.
4.2 Erythrophagocytose et recyclage du fer héminique par les macrophages Les macrophages sont des phagocytes professionnels chargés de l’élimination de nombreux microorganismes, tels que les cellules apoptotiques, les pathogènes intracellulaires et les globules rouges sénescents. Ces différentes activités sont essentielles à l’immunité innée et adaptative, au remodelage tissulaire et au métabolisme du fer [80]. En effet, dans les maladies inflammatoires chroniques, l'anémie est fréquemment due à la rétention du fer dans les macrophages activés par les cytokines [81].
Chez l'Homme, plus de 60% du fer de l'organisme est incorporé dans l'hémoglobine des globules rouges dont la durée de vie est limitée à 120 jours. Ensuite, les érythrocytes sénescents circulants seront phagocytés par les macrophages de la rate, de la moelle osseuse et du foie. Ce processus permet de recycler 25 à 30 mg du fer par jour, correspondant aux besoins en fer nécessaires pour produire journellement environ 200 milliards de nouveaux érythrocytes. Ainsi, l’érythrophagocytose est une composante fondamentale de l’homéostasie du fer, et le recyclage du fer héminique représente la principale source du fer pour l’érythropoïèse [45].
4.2.1 Mécanismes de l’érythrophagocytose et du recyclage du fer (Figure 6)
L’accumulation progressive de modifications biochimiques membranaires par les globules rouges circulants conduit à la reconnaissance spécifique des globules rouges sénescentes. Cette étape est suivie par l’internalisation du globule rouge par phagocytose. Ensuite, le phagosome continue sa maturation par une série d’événements de fusion membranaire avec différents compartiments vésiculaires, tels que les endosomes précoces, les endosomes tardifs et les lysosomes pour former un phagolysosome. Dans ce dernier, l’hème est dégradée par l’hème oxygénase 1 (HO-1), ainsi le Fe2+ est libéré et sort de l’endosome par DMT1 qui est
un membre de la famille des protéines associées à la résistance naturelle du macrophage (NRAMP) « natural resistance-associated macrophage protein » [82].
Une fois libéré dans le cytosol, le fer héminique sera soit capté par la ferritine pour former le pool de réserve, soit recyclé vers le plasma à travers la FPN1, qui exporte le fer vers l’espace extracellulaire. Ensuite, Fe2+ sera oxydé en Fe3+ par la céruloplasmine (CP) et se fixera sur la transferrine plasmatique pour être distribué aux précurseurs érythroïdes de la moelle osseuse [83]. De même les macrophages sont capables de recycler et de stocker le fer à travers le récepteur CD163 « Cluster of differentiation 163 » qui entraînera l’endocytose du complexe hémoglobine (HB) et haptoglobine (HP), ce qui représente aussi une voie importante pour l'absorption du fer dans les macrophages [84]. Enfin, les macrophages sont aussi capables de capter le fer à travers un mécanisme dépendant du complexe Tf-Fe3+-TfR1.
Figure 6. Erythrophagocytose et de recyclage du fer héminique. Les macrophages sont capables de recycler et de stocker le fer contenu dans l’hème des globules rouges sénescentes. L’hème est dégradée par l’hème oxygénase 1 (HO-1), ainsi le Fe2+ est libéré et sort de l’endosome par DMT1. Dans le cytosol, le fer sera soit capté par la ferritine pour former le pool de réserve, soit recyclé vers le plasma à travers la ferroportine1 (FPN1). Par suite, le Fe2+ sera oxydé en Fe3+ par la céruloplasmine (CP) et se fixera sur la transferrine plasmatique pour être distribué aux précurseurs érythroïdes de la moelle osseuse.
4.3 Absorption intestinale du fer
L’absorption du fer alimentaire s’effectue au niveau des entérocytes des villosités du duodénum et de la partie proximale du jéjunum. Ces derniers sont issus de la maturation des cellules souches des cryptes. Dans les conditions normales, l’apport alimentaire en fer apporté par l’absorption intestinale est de 1 à 2 mg par jour [43]. Malgré que cette quantité absorbée soit faible, l’absorption intestinale du fer constitue une étape capitale qui doit être bien régulée.
Les cellules souches des cryptes expriment à leur surface le TfR, et sont donc capables de capturer le fer par un mécanisme dépendant de la Tf [85, 86].
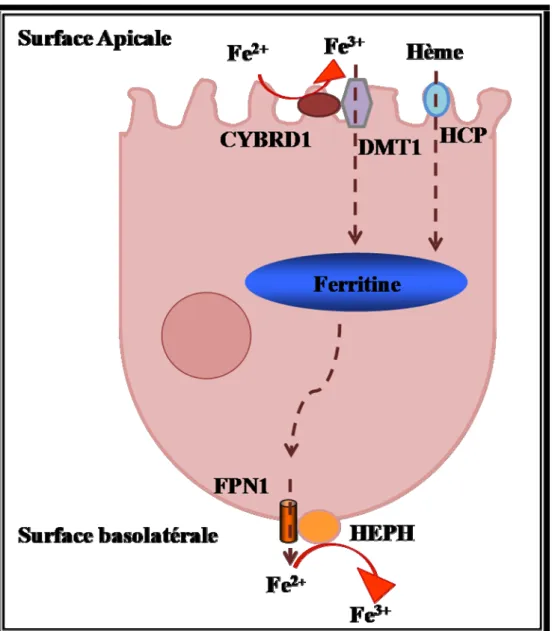
Le fer provenant de la diète se trouve sous deux formes : le fer héminique contenu dans la viande et le fer non héminique présent dans les végétaux et les produits laitiers [87]. Dans les entérocytes matures, le passage du fer héminique ou non héminique, de la lumière intestinale au plasma comporte trois étapes (Figure 7): 1) Franchissement de la membrane apicale, de la lumière digestive vers l’entérocyte.
2) Transport à l’intérieur de l’entérocytes, de la membrane apicale vers la membrane basolatérale.
4.3.1 Absorption du fer non héminique
Les deux formes Fe 2+ et Fe 3+ sont présentes dans la lumière intestinale. Pour être absorbé, le Fe3+ contenu dans l’alimentation sera réduit en Fe2+ par une ferriréductase membranaire la cytochrome b réductase 1 (CYBRD1) présente d’une façon abondante au niveau de la membrane apicale des entérocytes [88]. Ensuite, le Fe2+ est transporté vers le cytoplasme par DMT1 qui est présent à des taux élevés dans les entérocytes de la bordure en brosse duodénale [89]. Ainsi, le Fe2+ rejoint le pool du transit à partir duquel il sera soit stocké dans la ferritine, soit exporté au pôle basolatérale de l’entérocyte pour rejoindre la circulation sanguine par l’action coordonnée de FPN1 et l’hephaestine (HEPHA), un homologue de la céruloplasmine [63, 64].
4.3.2 Absorption du fer héminique
L’absorption du fer héminique s’effectue d’une façon presque similaire à celle du fer non héminique. En premier lieu, le fer héminique est transporté vers le cytoplasme par le transporteur apical HCP1 (Hème Carrier Protein 1) [90]. Ensuite, l’hème est dégradé par HO-1 et ainsi le Fe2+ est libéré et peut rejoindre le pool du fer non héminique internalisé par DMT1.
4.3.3 Absorption du fer par la voie du complexe intégrine–mobilferrine
Un autre mécanisme permettant également l’absorption du fer non héminique, fait intervenir la voie du complexe intégrine–mobilferrine « integrin-mobilferrin pathway » (IMP). À la surface cellulaire des entérocytes, l’intégrine et la mobilferrine se lient uniquement au Fe3+ et le transportent vers le cytoplasme. Une fois dans le cytoplasme, le complexe protéique intégrine-mobilferrine se combine à la flavine monooxygénase et à la β2-microglobuline pour former la paraferritine. Cette dernière est un complexe ayant une masse moléculaire de 520 kDa possédant une activité ferriréductase permettant la conversion du Fe3+ en Fe2+. Cependant, ce modèle d’absorption est seulement observé dans les entérocytes et son mécanisme exact n’est pas encore complètement élucidé [91].
Figure 7. Absorption intestinale du fer alimentaire par les entérocytes.
Le fer héminique sera réduit en Fe2+ par la cytochrome b réductase 1 (CYBRD1), ensuite le Fe2+ est transporté vers le cytoplasme par DMT1, tandis que le fer héminique est transporté vers le cytoplasme par le transporteur apical HCP1 (Hème Carrier Protein), pour rejoindre ensuite le pool du fer non héminique. Ainsi, le Fe2+ rejoint le pool du transit à partir duquel il sera soit stocké dans la ferritine, soit exporté au pôle basolatérale de l’entérocyte pour rejoindre la circulation sanguine par l’action coordonnée de ferroportine (FPN1) et hephaestine (HEPH).
4.4 Régulation systémique du fer
L’Homme ne possède pas une régulation de voie physiologique pour l'excrétion du fer. Par conséquent, l'homéostasie du fer dans l'organisme est régulée au niveau de son absorption. C’est pour cela que la régulation inadaptée de l’absorption du fer conduit à une carence ou bien à une surcharge en cet ion métallique [34]. En fait, plusieurs facteurs contribuent au maintien de l'homéostasie du fer, notamment la diète alimentaire, le contenu total en fer de l’organisme, l’activité érythropoïétique, l’hypoxie et les infections.
a) La diète alimentaire :
L’absorption intestinale du fer est influencée par la quantité du fer récemment apportée par l’alimentation. Ainsi, une quantité excessive du fer dans l’alimentation pendant plusieurs jours peut induire une moindre absorption du fer par les entérocytes. Ce phénomène est aussi connu sous le nom de « bloc muqueux » [92]. b) Le contenu en fer de l’organisme :
L’absorption intestinale du fer varie d’une façon inversement proportionnelle au stock du fer dans l’organisme. Il est bien établi que dans des conditions déficientes en fer, l'absorption est significativement stimulée, tandis que lorsque les réserves en fer sont reconstituées, l’absorption revient au niveau basal. Ce type de régulation semble intervenir dans la programmation des cellules souches des cryptes qui agissent comme senseur du niveau de saturation de la Tf sérique. En fait, la quantité du fer absorbée à partir de l’alimentation est déterminée par les concentrations de DMT1 et de FPN1 au niveau des entérocytes matures et ces concentrations sont probablement programmées par l’action du système IRE/IRP en fonction de la quantité du fer au niveau des cellules cryptiques [41].
c) L’activité érythropoïétique :
La majorité du fer de l'organisme est utilisée par la moelle osseuse pour la synthèse de l'hémoglobine des globules rouges durant l’érythropoïèse. Pour cela, l’activité érythropoïétique a une fonction dominante dans le contrôle de l'homéostasie du fer. En d'autres termes, ce signal a une plus grande capacité d’augmenter l'absorption du fer par rapport au contenu total en fer de l’organisme [34, 93]. En outre, dans les troubles touchant l'érythropoïèse tels que la thalassémie, l’anémie sidéroblastique et l’atransferrinémie, ce signal augmente l'absorption du fer indépendamment des réserves corporelles, ce qui entraîne une accumulation pathologiques du fer [2]. d) L’hypoxie :
L’hypoxie aiguë entraîne une augmentation de l’absorption du fer, ainsi que la production de l'érythropoïétine (EPO) qui est une hormone qui stimule l’érythropoïèse. Cela va produire un effet semblable au signal de l’activité érythropoïétique. Cependant, il est incertain si cette voie de régulation est vraiment distincte de celle induite par les régulateurs de l’activité érythropoïétique [60]. e) Les infections :
Les infections sont des régulateurs puissants du métabolisme du fer. Durant les infections, l’organisme stimulera la séquestration du fer pour empêcher le développement des bactéries pathogènes. Pour cela, la libération du fer par le foie et les macrophages ainsi que l’absorption intestinale sont diminuées [94]. Les régulateurs inflammatoires sont dominants par rapport aux régulateurs érythropoïétiques. Par exemple, dans l'anémie des maladies des infections chroniques, les cytokines inflammatoires réduisent l’érythropoïèse due à la séquestration du fer. Par contre, cela activera le régulateur érythropoïétique qui va stimuler l'absorption du fer.
Enfin, dans cette situation contradictoire, le régulateur inflammatoire domine le régulateur érythropoïétique entraînant une diminution de l’absorption intestinale et une anémie inflammatoire chronique [95].
L’hiérarchie entre les différents régulateurs de l’homéostasie de cet ion prédit l'existence d’un facteur permettant de coordonner correctement l'utilisation du fer de stockage, d'absorption et de recyclage [70]. Ainsi, la recherche des agents modulant l'homéostasie a fait une évolution considérable avec la découverte de l'hepcidine qui est considéré comme le régulateur central de la l'homéostasie du fer [96].
5. Hepcidine
L’hepcidine tient son nom de son site majeur de synthèse « hep » pour hépatique et « idine » de son activité bactéricide [97]. L’hepcidine est un peptide hormonal, appartenant à la famille des β-défensines, synthétisé dans le foie, distribué dans le plasma et excrété dans l'urine [98].Cette hormone peptidique est en effet l’hormone du fer autant recherchée. Elle joue un rôle hyposidérémiant en réglant l'absorption intestinale du fer, le recyclage du fer par les macrophages et la mobilisation du fer hépatique [99].Outre son rôle dans la régulation du métabolisme du fer, l’hepcidine joue un rôle important dans la défense innée de l’hôte contre les infections [100]. Cependant, des études récentes ont clairement montré que la déficience en hepcidine due à la dérégulation de sa synthèse semble être la cause ultime de la plupart des formes des maladies de surcharge en fer [101].
5.1 Découverte de l’hepcidine comme étant un peptide antimicrobien
L’hepcidine a été découverte dans l’urine humaine et était nommé hepcidine [100]. Indépendamment, un autre groupe de recherche a découvert le même peptide dans le plasma et l'a nommé LEAP-1 « liver-expressed antimicrobial peptide » [102]. Appartenant à la famille des β-défensines, l'hepcidine ressemble structurellement à la famille des défensines à cause des quatre ponts disulfures dans sa structure tertiaire. En plus, tel que la plupart des peptides antimicrobiens naturels, l’hepcidine présente deux propriétés fondamentales: la charge cationique et le caractère amphipathique (hydrophobe et hydrophile) qui facilitent sa capacité à perforer les membranes des pathogènes [103].
Comme certains peptides antimicrobiens, l'hepcidine présente une double fonction : En plus de son rôle hyposidérémiant dans le métabolisme du fer, l'hepcidine fait partie du système immunitaire inné et constitue ainsi la première ligne de défense contre les infections fongiques et bactériennes. Des études ont montré, que l’activité antimicrobienne et antifongique de l’hepcidine sont beaucoup plus efficaces dans l’urine que dans le sang. En fait, l'hepcidine est active contre les bactéries Gram+ comme Staphylococcus aureus, les bactéries Gram- par exemple Escherichia coli et les levures tel que Candida albicans [104].
Ainsi, l'hepcidine agit de deux manières contre les bactéries et les levures : 1) En attaquant et perforant directement la membrane des pathogènes [98].
2) En limitant la quantité du fer libre, ce qui inhibe la prolifération des micro-organismes [98].
5.2 Structure de l’hepcidine
Un seul gène code pour l’hepcidine chez l’Homme, tandis qu’il y a deux gènes codant l’hepcidine chez la souris, (hepc1 et hepc2), qui présentent respectivement une homologie de 76 et 58% avec le gène hepcidine humain [105, 106].
Le gène hepcidine humain (HAMP) est situé sur le bras long du chromosome 19. Les 2.5 Kb du gène Hamp contiennent deux introns et trois exons qui se transcrivent en ARNm de 0.4 Kb [101]. Cette ARNm est traduit en préprohepcidine de 84 acides aminés ayant dans sa partie N-terminale un site de clivage par la furine. Le clivage enzymatique permet d’obtenir un peptide de 25 acides aminés côté C-terminal, appelé hepcidine-25, qui constitue la seule forme mature et active [107-109].
L’étude d’hepcidines synthétiques mutées a permis d’établir que les 5 acides aminés N-terminaux sont essentiels pour l’activité de l’hepcidine [110]. À noter qu’il existe d’autres formes d’hepcidine de 20 et 22 acides aminés retrouvées essentiellement au niveau urinaire, provenant probablement de la dégradation de l’hepcidine 25 [100].
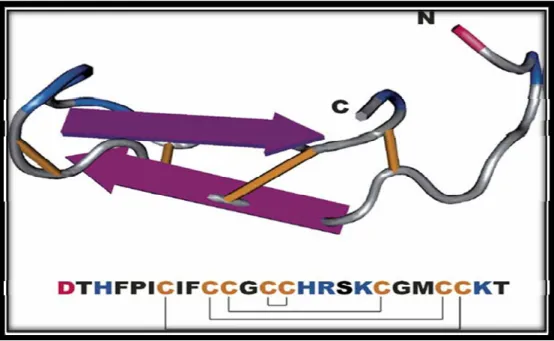
L’étude par spectrométrie de masse montre que le peptide natif de 25 acides aminés est riche en feuillets β et contient 8 cystéines formant 4 ponts disulfures [100]. Au cours de l’évolution, ces cystéines ont été très conservées et sont retrouvées dans différentes espèces [104] (Figure 8).
Par ailleurs, l’étude par résonance magnétique nucléaire, montre que l’hepcidine 25 se présente en forme d’épingle à cheveux dont les deux feuillets β anti-parallèles sont stabilisés par 4 ponts disulfures. L’un des ponts disulfures est formé par deux cystéines adjacentes situées dans le coude β de l’épingle, ce qui tend à renforcer extrêmement cette structure et à conférer une stabilité pour l’hepcidine en circulation [111] (Figure 9).
5.3 Sites d’expression de HAMP
HAMP est principalement exprimé au niveau du foie [102, 105], notamment dans les hépatocytes [112]. Également, l’expression de l’hepcidine est détectée dans d’autres sites, mais à plus faibles concentration, comme la rate, le cœur, l’estomac, le cerveau, les poumons et le pancréas. La détection de l’hepcidine dans les reins a été rapportée, mais son rôle dans ce tissu reste inconnu [113]. Par ailleurs, l’hepcidine est exprimée dans les macrophages activés, tels que les macrophages spléniques [114], les macrophages dérivés des monocytes [115] et les macrophages pulmonaires [116]. Enfin, une étude récente montre que le tissu adipeux des patients obèses présente des taux élevés d’hepcidine par rapport au tissu adipeux d’individus non obèses, ce qui peut contribuer à l’hyposidérémie observée chez 70 % de ces patients [117].
Figure 8. Séquences de l’hepcidine chez les mammifères et les poissons. Les espèces de mammifères sont : L'Homme (h), cochon (p), rat (r) et la souris (m). Les poissons sont : Bar (b), flet (f) et long-jawed mudsucker (g). Les cystéines conservées sont encadrées en rose. Adapté de [118].
Figure 9. Structure de l’hepcidine humain.
L’hepcidine 25 se présente en forme d’épingle à cheveux dont les deux feuillets β anti-parallèles contiennent 8 cystéines et sont stabilisés par 4 ponts disulfures. L’un des ponts disulfures est formé par deux cystéines adjacentes situées dans le coude β de l’épingle. Les ponts disulfures et les cystéines sont représentés en jaune. Adapté de [78].