Surdité unilatérale : approches psychoacoustique, qualité de vie et neuroimagerie fonctionnelle par IRMf
263
0
0
Texte intégral
Figure

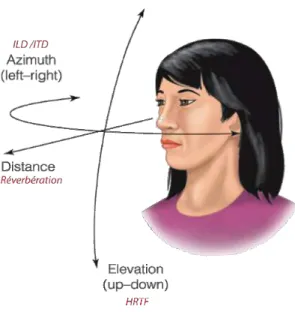

![Figure 2.14 – Schématisation de l’expérience de stream ségrégation de Van Noorden illustrée par McAdams [120]](https://thumb-eu.123doks.com/thumbv2/123doknet/2240093.17065/35.892.324.569.450.725/figure-schematisation-experience-stream-segregation-noorden-illustree-mcadams.webp)
+7

Documents relatifs