THESE
Présentée devant
L’UNIVERSITE PAUL SABATIER DE TOULOUSE (SCIENCES)
En vue de l’obtention du
Doctorat de l’Université Paul Sabatier- Toulouse 3
Spécialité : Biomécanique par
Franck ACCADBLED
Docteur en Médecine, Chef de Clinique-Assistant
CHU TOULOUSE
Contribution à l'analyse du transport convectif dans le segment
vertébral: étude de la perméabilité macroscopique
du plateau vertébral
Soutenance le 25 septembre 2007 devant le Jury d’examen :
Pr J.P. Cahuzac, PU-PH, Hôpital des Enfants, Toulouse 3 Examinateur Pr P. Chabrand, PU, Université de la Méditerranée Rapporteur Pr C. Glorion, PU-PH, Hôpital NeckerEnfants Malades, Paris V Rapporteur Pr J. Sales de Gauzy, PU-PH, Hôpital des Enfants, Toulouse 3 Examinateur
Pr P. Swider, PU, Toulouse 3 Examinateur
Remerciements
Je tiens à remercier sincèrement :
Monsieur le Professeur Christophe Glorion pour m’avoir accueilli dans son service et avoir accepté d’être le rapporteur de ce travail.
Monsieur le Professeur Patrick Chabrand, pour avoir accepté d’être le rapporteur de ce travail.
Monsieur le Professeur Eric Viguier pour son accueil à l’Ecole Vétérinaire de Lyon.
Monsieur le Professeur Jean-Philippe Cahuzac qui m’a donné le goût pour la recherche, clinique et fondamentale.
Monsieur le Professeur Jérôme Sales de Gauzy, pour son soutien et ses conseils. Monsieur le Professeur Pascal Swider pour sa disponibilité et son aide précieuse.
Monsieur le Professeur André Autefage, Mlle Sophie Palierne de l'Ecole Nationale Vétérinaire de Toulouse pour leur accueil et leur collaboration.
Merci à Dominique Ambard, notre association en binôme a été particulièrement constructive et agréable au quotidien.
Merci à Jean-Michel Laffosse pour sa précieuse participation à ce travail. Merci à Thierry Odent pour une collaboration qui va certainement perdurer
Merci à Jérôme Briot et Erik Estivalèzes, du Laboratoire de Biomécanique EA 3697 de l’Hôpital Purpan à Toulouse pour leur collaboration, leur aide et leurs conseils.
Enfin je remercie bien sûr ma famille, Stéphanie et tous mes amis ainsi que mes collègues du Women’s and Children’s Hospital d’Adélaide.
SOMMAIRE
RESUME………...………5
ABSTRACT………..6
INTRODUCTION & OBJECTIFS……….7
CHAPITRE 1 : Contexte, étude bibliographique…………..………9
1.1 : Vertèbre et disque intervertébral………9
1.1.1 : Développement………..…………..9
1.1.2 : Anatomie………..………..11
1.1.3 : Histologie, Biochimie………....12
a - Disque intervertébral b - Plateau cartilagineux 1.1.4 : Fonction & biomécanique………..15
a- Disque intervertébral b - Plateau cartilagineux 1.1.5 : Maturation,dégénérescence………...……….16
1.2 : Considérations sur les transports convectifs dans le segment vertébral………...17
1.2.1 : Comportement d’un milieu poreux élastique………...17
a - Relation contrainte, déformation, pression de fluide b - Relation débit, pression de fluide, perméabilité 1.2.2 : Application au plateau vertébral, implication dans la physiologie rachidienne………20
1.2.3 : Etude qualitative du transport convectif………..…..21
1.2.4 : Etude quantitative du transport convectif………..….……...21
1.3 : Considérations sur les approches expérimentales……….….….…….22
1.3.1 : Modèle animal……….…..22
1.3.2 : Conservation des échantillons………..….22
a- Etat frais b- Congélation 1.4 Synthèse………..24
CHAPITRE 2 : Développement d’un dispositif de mesure de la perméabilité du plateau vertébral, application à un modèle animal porcin 2.1 : Objectif……….25 2.2 : Matériel et méthodes……….….. 25 2.2.1 : Echantillons biologiques………..……..25 2.2.2 : Considérations théoriques……….…….26 2.2.3 : Dispositif expérimental……….……….27 2.3 : Résultats………...………….29
2.3.1 : Evaluation de la précision du dispositif de mesure………29
2.3.3 : Perméabilité macroscopique du plateau vertébral……….………..…..30
2.4: Discussion et conclusion………...….31
CHAPITRE 3 : Influence de l’âge, de la localisation et du sens du flux sur la perméabilité du plateau vertébral; application à un modèle animal ovin 3.1 : Objectif……….…....34
3.2 : Matériel et méthodes………..…...34
3.2.1 : Echantillons biologiques……….….…..34
3.2.2 : Dispositif de mesure &procédure expérimentale………..35
3.3 : Résultats………....36
3.3.1 : Perméabilité………..…….36
3.3.2 : Etude microscopique……….……….39
3.4 : Discussion et conclusion………...40
CHAPITRE 4 : Influence d’une épiphysiodèse asymétrique sur la perméabilité du plateau vertébral; application à un modèle animal porcin 4.1 : Objectifs………42
4.2 : Scoliose expérimentale, étude bibliographique………...…….42
4.3 : Matériel et méthodes……….44
4.3.1 : Instrumentation et prélèvement des échantillons………..…….44
4.3.2 : Dispositif de mesure et procédure expérimentale………..……45
4.4 : Résultats………..……..45 4.4.1 : Perméabilité macroscopique………..46 4.4.2 : Etude microscopique………....………..47 4.5 : Discussion et conclusion………...………47 CONCLUSIONS ET PERSPECTIVES………..….49 REFERENCES………52 ANNEXES………...57
Les travaux présentés ici ont été réalisés conformément aux textes réglementaires relatifs à la protection et l’utilisation des animaux à des fins expérimentales.
RESUME
Le comportement mécano-biologique du segment vertébral, sain et pathologique, est un problème complexe. La préoccupation clinique première de notre travail concerne la pathologie du rachis chez l'enfant et notamment la scoliose idiopathique de l’adolescent. L’hypothèse centrale a été que la modification de nutrition et d’homéostasie des disques pouvait être associée à une perturbation du transport convectif au niveau du plateau vertébral. Nous avons développé, à partir de modèles animaux, une démarche de recherche progressive, associée dans son questionnement à la problématique clinique chez l'enfant. L'étude bibliographique préliminaire a permis de mettre en évidence le rôle significatif des transports de masse (fluides, protéines, nutriments) dans le développement sain et pathologique du rachis. Ceci a abouti aux quatre points clés de la stratégie de recherche: variabilité topographique de la perméabilité, influence du sens de flux, rôle de la croissance et influence d'une perturbation mécanique associé à une courbure imposée. Dans la deuxième partie, une méthode originale de mesure de perméabilité macroscopique d'échantillons de tissus biologiques a été développée puis évaluée sur un modèle animal porcin. Cette étude a montré que la perméabilité macroscopique du plateau vertébral n'était pas uniforme, la zone centrale étant plus perméable que la périphérie. L'étude microscopique associée a montré une corrélation entre l'augmentation de la perméabilité et la diminution des épaisseurs de cartilage. Dans un troisième temps, l'évolution des perméabilités de plateaux vertébraux a été mesurée sur un modèle animal ovin en croissance normale. Il a été mis en évidence une diminution de la perméabilité macroscopique du plateau vertébral avec la maturation des tissus. Le flow-out, (exsudation de fluide hors du disque), a été associé à des perméabilités plus grandes qu'en flow-in (réhydratation du disque). Il est apparu que la séquence des tissus constitutifs et leurs propriétés intrinsèques (porosité, tortuosité évolutive) pouvaient modifier la perméabilité macroscopique du plateau vertébral. En dernier lieu, un modèle de croissance asymétrique de segment (épiphysiodèse en L1-L2) a été mis en place sur un modèle animal
porcin. Ce modèle a confirmé les résultats précédents et les perméabilités globales des plateaux mécaniquement perturbés ont été inférieures à celle des témoins. L'effet de pincement dissymétrique a eu tendance à faire décroître la perméabilité des tissus comprimés. En conclusion, nous avons montré que la perméabilité macroscopique du plateau vertébral était dépendante de la zone d'intérêt, du sens de l'écoulement de fluide et dépendante de la maturité des tissus et donc de la croissance. Il est donc apparu que les propriétés de transport convectif du plateau pouvaient constituer un point clé du remodelage mécano-biologique du rachis en croissance, sain et pathologique.
ABSTRACT
The mechano-biological behaviour of the healthy or pathological vertebral segment is a complex problem. The clinical concern of our work related to the idiopathic scoliosis in adolescent.
The central assumption was that the modification of nutrition and homeostasis of the intervertebral disc could be associated to a disorder of the convective transport through the vertebral end plate. We designed a progressive research based on experimental animal models keeping in mind the clinical relevance of our approach. The preliminary bibliographical study highlighted the significant role of mass transport (fluids, proteins, nutrients) in the physiological and pathological growth of the spine. This allowed four key points of our strategy to be defined: geometrical variability of the permeability, influence of the fluid-flow direction, role of the growth and influence of a mechanical disturbance associated to an imposed curve. In the second part of the study, an original method of measurement of the macroscopic permeability has been developed then evaluated in a porcine animal model. This study showed that the macroscopic permeability of the vertebral plate was not uniform, the central zone being more permeable than the periphery. Associated microscopic study demonstrated a correlation between the permeability increase and the reduction of cartilage thickness. Third, the variation of the vertebral plate permeability during growth was measured in the ovine animal model. We highlighted a decrease of the macroscopic permeability with tissue maturation. Flow-out, (exudation of fluid from the disc), showed greater permeability than flow-in (disc rehydration). It appeared that the sequence of constitutive tissue and their intrinsic properties (porosity, evolutive tortuosity) could modify the macroscopic permeability of the vertebral plate. Finally, an asymmetrical growth tether model was designed in L1-L2 in
a porcine animal model and confirmed the preceding results. The permeability of the mechanically modified plates was lower than that of their unaffected counterparts. The effect of dissymmetrical growth tended to decrease the permeability of compressed tissue.
In conclusion, we showed that the macroscopic permeability of the vertebral plate was dependent upon the zone of interest, the direction of fluid flow and the tissue maturity (growth). Thus, it appeared that the properties of convective transport of the end plate could constitute a key point of the mechano-biological remodelling in the physiological or pathological growth.
INTRODUCTION & OBJECTIFS
Le disque intervertébral (DIV) est un élément anatomique majeur du rachis. Il est structurellement constitué d'un squelette solide saturé par une solution aqueuse. La matrice solide est essentiellement composée de fibres de collagène et d’agrégats de protéoglycanes (Pg), dont les proportions varient suivant les différentes régions du DIV. Les cellules, chondrocytes et fibroblastes, participent à son homéostasie en synthétisant (ou en dégradant) les fibres de collagène et les chaînes de Pg. Le métabolisme cellulaire fournit l'énergie nécessaire (glucose, oxygène, …). Mais le DIV n'étant pas vascularisé, les nutriments indispensables à la croissance et à l'homéostasie du segment vertébral transitent majoritairement au travers des plateaux cartilagineux disposés de part et d'autre du DIV. Ce transport est effectué par la solution aqueuse saturante. Les déchets de la respiration cellulaire, doivent eux aussi être évacués via la phase liquide. Le transfert de ces nutriments est réalisé par différents phénomènes physiques, telle que la diffusion moléculaire, phénomène relativement lent, et par transport convectif. De plus, l'activité physique et les phases de repos entraînent des cycles de chargement mécanique et provoquent des circulations de fluide entre le DIV et les corps vertébraux. Cet effet de pompage est le vecteur principal de transport de nutriment vers le DIV.
Le comportement mécano-biologique du segment vertébral, sain et pathologique, est donc un problème complexe et nous pouvons constater qu'il constitue encore à ce jour un problème scientifique et clinique totalement ouvert.
Notre problématique clinique première est la scoliose idiopathique chez l'enfant. Dans ce contexte les interactions entre courbure, stimuli mécaniques et homéostasie des DIV ont été très peu étudiées. L'étude quantitative du transport convectif à l’interface DIV - corps vertébral demeure également peu développée or l'analyse rigoureuse des transferts moléculaires devrait contribuer à une meilleure compréhension de la pathologie et à l'amélioration de son traitement chirurgical notamment le choix du niveau inférieur de l’instrumentation.
L’hypothèse centrale du projet consiste donc à supposer que la modification de nutrition et d’homéostasie des DIV peut être associée à une perturbation du transport convectif au niveau du plateau vertébral et le schéma synoptique de la Figure 1 permet de résumer la stratégie mise en place. Nous pensons donc que la perméabilité macroscopique du plateau vertébral peut être un point clé de la boucle de contrôle mécano-biologique du rachis. Nous pouvons observer que le point initial de la démarche est la scoliose idiopathique de l’adolescent, mais il est prématuré de développer des études quantitatives à partir des données cliniques. Dans ce cas une approche parallèle sur modèle animal est développée. Cette stratégie permet de développer les techniques expérimentales et grâce à une progression dans la complexité, elle s'efforce de jalonner le parcours qui devrait permettre à terme d'obtenir des conclusions robustes utilisables chez l'enfant scoliotique.
Dans un premier temps, le contexte général de l'étude est explicité dans le chapitre 1. Celui-ci est consacré à une description anatomique, biologique et fonctionnelle du DIV ainsi qu’à une analyse des travaux de la littérature concernant le transport convectif dans le segment vertébral et plus particulièrement, la perméabilité de tissus biologiques.
Dans le deuxième chapitre, nous décrivons un dispositif original de mesure de la perméabilité intrinsèque du plateau vertébral et sa validation par une étude pilote sur un modèle animal porcin.
Plusieurs facteurs sont susceptibles de modifier la perméabilité du plateau vertébral : l’âge du sujet, la région intéressée sur le plateau ou le sens d’écoulement. Ces facteurs sont étudiés dans le chapitre 3 par une expérimentation sur l’agneau en croissance normale.
Dans le chapitre 4, nous étudions l’effet d’une épiphysiodèse vertébrale asymétrique sur la perméabilité dans le cas d'un modèle animal porcin en croissance.
En dernier lieu, une synthèse des résultats obtenus est présentée, ainsi que des perspectives de développement concernant les aspects scientifiques et cliniques de l'étude dans le cadre général des recherches interdisciplinaires Sciences / Santé.
Etude du transport convectif dans des modèles animaux
Scoliose idiopathique chez l'enfant
Modèle porcin perméabilité macroscopique des
plateaux vertébraux Modèle ovin
perméabilité macroscopique des plateaux vertébraux en croissance normale
Modèle porcin
perméabilité macroscopique des plateaux vertébraux en croissance pathologique
Fig. 1 - Schéma synoptique de la stratégie de recherche aide au diagnostic & traitement chirugical
CHAPITRE 1 - Contexte, revue bibliographique
1.1 : Vertèbre et disque intervertébral
1.1.1 : Développement
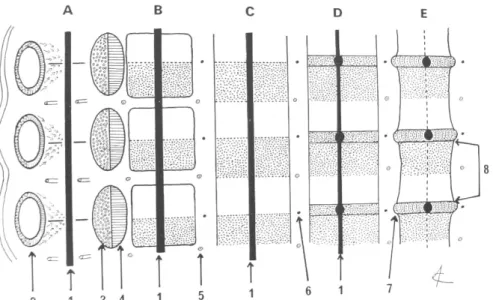
La priorité accordée au rachis dans la morphogénèse du squelette souligne son importance et notre appartenance au groupe des vertébrés. Le développement du DIV et des vertèbres adjacentes débute très tôt durant la vie intra-utérine et sont étroitement liés. A la 3ème semaine, l’embryon est constitué de 3 feuillets : entobaste, ectoblaste et mésoblaste. Le mésoderme forme latéralement de part et d’autre de la chorde 2 colonnes longitudinales : le mésoderme para axial. Ce mésoderme se segmente pour former les somites (Fig. 2).
Chaque somite se creuse ensuite d’une cavité qui permet de distinguer du côté interne les sclérotomes qui vont former les vertèbres, et du côté externe les myotomes qui vont former les muscles, les dermatomes qui vont former le derme et le tissu sous cutané. Chaque sclérotome se clive ensuite horizontalement. La partie crâniale est à l’origine du DIV. Elle fusionne à la partie caudale du sclérotome sus-jacent qui est à l’origine du corps vertébral. Chaque vertèbre se développe donc à partir de 2 sclérotomes adjacents. La vertèbre est une
structure intersegmentaire (Fig. 3).
La chorde va régresser au sein des corps vertébraux lors de leur différenciation cartilagineuse puis osseuse. Sa dégénérescence mucoïde va former le nucleus pulposus (NP). Le disque péri-notochordal mésenchymateux se différencie en deux zones : une zone périphérique fibreuse faite de fibroblastes et de fibres de collagène et une zone centrale plus lâche. Ces 2 zones vont former l’annulus fibrosus (AF). Le DIV (NP + AF) est donc une
structure d’origine mixte, chordale et somitique, mais elle n’est pas intersegmentaire. Fig. 2 : Embryon 3 semaines
(d’après JM. Clavert : Chirurgie et Orthopédie du Rachis, Sauramps24)
La maturation post-natale se traduit par la disparition des vestiges chordaux et la transformation du NP en fibrocartilage à mesure qu’il se charge en fibres de collagène (Fig. 4). Chez l’adulte, vers l’âge de 30 ans, le NP est solide, fibreux et n’est plus démarqué aussi clairement de l’AF. On parle alors, notamment en IRM, de complexe central ou interne composé du NP et de la partie centrale de l’AF, et de complexe périphérique constitué de la partie périphérique de l’AF qui s’insère à la périphérie des plateaux vertébraux. Quant à la croissance de la vertèbre, elle débute dès le 3ème mois in utero pour se terminer vers la 25ème année de vie (Fig. 5). Elle va se faire dans 2 directions, en hauteur et en largeur par ossification enchondrale.
Fig. 3 : La vertèbre est une structure inter segmentaire.
A : Migration du sclérotome. B : Condensation mésenchymateuse. C : Organisation inter segmentaire. D : Morphogénèse du disque. E : Structure vertébrale définitive. 1. Chorde. 2. Somite. 3. Dermatome. 4. Myotome. 5 Artère inter segmentaire. 6. Nerf spinal. 7. Disque intervertébral. 8. Corps vertébral. (d’après JM. Clavert : Chirurgie et
Orthopédie du Rachis, Sauramps24)
Fig.4 : Régression chordale et formation
du NP (d’après Diméglio et Bonnel. Le
Les vertèbres ont une croissance différentielle; l’activité des cartilages de croissance croît du segment cervical au segment lombaire. Le phénomène d’ossification débute très tôt vers la 7ème semaine du développement pour se terminer à la fin de la croissance. Elle se fait progressivement par transformation des zones cartilagineuses. On retrouve 3 zones d’ossification pour chaque vertèbre : 1 point d’ossification volumineux par corps vertébral et 1 point d’ossification par hémi-arc postérieur. Ces 3 centres primaires sont retrouvés à la naissance. Durant la 1ère année de vie, les 2 hémi-arcs postérieurs fusionnent et entre 3 et 6 ans, l’arc postérieur fusionne avec le corps vertébral. La séparation entre corps vertébral et arc postérieur est matérialisée par le cartilage neurocentral, nécessaire à l’adaptation du canal vertébral à la taille de la moelle épinière. On dénombre 5 centres d’ossification secondaires: 1 pour l’apophyse épineuse, 1 pour chaque apophyse transverse et 1 plaque par plateau vertébral. En fin de croissance, les disques représentent 1/4 de la hauteur totale du rachis cervico-dorsal, et 1/3 de la hauteur totale dans la région lombaire.
Le développement du rachis est directement lié aux contraintes auxquelles il est soumis et la verticalisation est nécessaire à une anatomie normale30. Il a été observé une hauteur plus importante des corps vertébraux et un diamètre antéro postérieur diminué chez des enfants grabataires ainsi que des DIV d’une épaisseur moindre et des NP hypotrophiques30,38.
1.1.2 : Anatomie
Les corps vertébraux sont unis entre eux par l'intermédiaire des DIV dont l'épaisseur varie entre 3mm (premiers disques thoraciques), 5 à 6mm (disques cervicaux) et 10 à 15mm (disques lombaires). Dans les régions cervicale et lombaire, le DIV est plus épais en avant qu'en arrière. Le DIV est une structure non vascularisée et peu innervée. Le plateau cartilagineux ou "cartilage end-plate" (CEP) s'intercale entre les plateaux vertébraux et le disque auquel il adhère très intimement. Il est constitué de cartilage hyalin et d’une mince couche d’os sous chondral. Son épaisseur moyenne chez l’humain est de 0,6mm (0,1 à 1,6mm) elle est d’environ 1mm en périphérie en regard de l’AF puis diminue en regard du NP68,88,89. Cette dépression de la partie centrale est appelée « stress bearing zone » de Schmorl30. L’épaisseur augmente des vertèbres crâniales vers les vertèbres caudales jusqu’en L3-L4. Elle est identique pour les plateaux supérieur et inférieur de chaque vertèbre.
Les autres liaisons entre deux vertèbres adjacentes comportent les processus articulaires en postérieur, et un système ligamentaire. Ces ligaments (jaune, capsulaire, interépineux et supraépineux, longitudinaux postérieur et antérieur, intertransverse) sont liés à une unité
fonctionnelle (ensemble de deux vertèbres et des tissus de liaison) représentée Figure 6. Fig.5 : Disque et vertèbre chez le
nouveau-né (d’après Diméglio et
Bonnel Le rachis en croissance,
Springer29)
Corps vertébral
Disque
Le DIV, participant à la flexibilité et à la stabilité du rachis, est constitué de deux éléments anatomiques distincts : l'AF et le NP. L’AF est une puissante enveloppe fibreuse fixée en périphérie aux plateaux vertébraux par des fibres de Sharpey. Elle est bordée par les ligaments longitudinaux antérieurs et postérieurs. Le NP est une matrice gélatineuse incompressible fortement hydratée localisée à la partie centrale du plateau vertébral.
1.1.3 : Composition, histologie
Le NP et la partie centrale de l’AF sont en contact direct avec le cartilage hyalin du plateau vertébral.
a – Disque intervertébral
Le NP est un gel hydrophile contenant plus de 90% d’eau dans l’enfance, cette valeur diminuant avec l’âge, environ 80% chez l’adulte jeune et moins de 70% chez le sujet âgé. Il représente environ 50% du volume d’un DIV adulte. La matrice extracellulaire est composée de collagène et de Pg. Le collagène constitue 20% du poids sec du NP, dont 80% de collagène de type II, 15% de type VI et 5% de collagènes IX et XI. Il ne contient pas de collagène de type I (qui est le collagène des structures tendineuses). Les Pg représentent 50% du poids sec du NP, ils lui confèrent son caractère hydrophile et sa consistance en gel visqueux. On observe avec l’âge, une diminution du pourcentage de Pg parallèlement à la perte d’eau. Le DIV contient peu de cellules, dont 3000/mm3 dans le NP. On distingue 2 types cellulaires : de petites cellules arrondies de type chondrocytaire et de grosses cellules vacuolaires qui synthétisent Pg et collagène de type II.
L'AF est formé de 7 à 15 lamelles concentriques constituées de fibres de collagène de type I disposées de façon oblique d’une couche à l’autre et formant un angle de 120° entre elles et de 30° par rapport au plan du DIV. Entre ces lamelles sont enchâssées les cellules et leur matrice. L’AF contient 60 à 70% d’eau, cette valeur reste relativement constante au cours de la vie. Environ 65 à 70% de son poids sec proviennent du collagène. Le collagène de type II est un des principaux constituants de la matrice extra cellulaire. Les collagènes I et II représentent 80% du collagène total, mais l’AF contient également du collagène de type III, V, VI, IX et XI. Le rapport collagène de type I/collagène de type II diminue de la partie centrale vers la périphérie. Les Pg représentent 10 à 20% du poids sec et sont essentiellement concentrés dans la partie centrale de l’AF. La cellularité de l’AF est faible elle aussi avec
Fig. 6 : Une unité fonctionnelle:
DIV + 2 vertèbres adjacentes et leurs liens ligamentaires
9000/mm3 alors que le plateau cartilagineux en contient 1500/mm3. Il s’agit de petites cellules rondes semblables à celles du NP.
Les 2 principaux constituants de la matrice extracellulaire du DIV sont les Pg et le collagène. La majorité des Pg sont capables de former des agrégats, ce sont principalement les agrécanes et en plus faible quantité les versicanes. Les agrécanes du DIV ont des propriétés physicochimiques semblables à celles des agrécanes du cartilage articulaire et peuvent être synthétisées par le NP comme par l’AF. Les agrécanes sont des molécules complexes formées d’une protéine centrale (core protein) possédant 2 domaines globulaires G1 et G2 dans la région N-terminale et un domaine globulaire G3 dans la région C-terminale. Entre G2 et G3, des glycosaminoglycanes (kératanes sulfates et chondroïtine sulfates) se fixent sur cette protéine porteuse3. Les glycosminoglycanes sont des sucres chargés négativement qui attirent les molécules d’eau et confèrent au DIV ses capacités de résistance à la compression. Le second élément essentiel de la matrice extracellulaire est le collagène. Il confère au DIV ses capacités de résistance en traction.
b - Plateau cartilagineux
Le CEP est intimement accolé au DIV. Il est constitué d’une fine couche de cartilage hyalin associé à une épaisseur d’os sous-chondral88. Duncan et al.32 ont défini l’os sous chondral comme étant la zone séparant le cartilage articulaire de l’os spongieux. Il est constitué de 2 couches : la partie calcifiée du cartilage articulaire et une couche d’os lamellaire. Le CEP est directement accolé à la plaque de croissance. Ainsi, il n’existe pas à proprement parler chez l’humain d’épiphyse vertébrale contrairement à d’autres mammifères. (Fig. 7).
Les fonctions biologiques et mécaniques du CEP sont sous-tendues par une architecture
complexe qui, comme celle du DIV évolue avec sa maturation et son vieillissement.
Les fibres de collagène du DIV semblent se poursuivre dans le CEP, formant un angle de 120° avec l’AF périphérique et de 90° avec le NP (Fig. 8). Ainsi, il existe des connections directes entre CEP et DIV, alors que le CEP et le corps vertébral sont nettement distincts68. Au niveau de la partie périphérique de l’AF, il existe une ligne basophile ou « tidemark » soulignant une couche de cartilage calcifié. Dans la région du NP, cette couche est très mince voire indiscernable (Fig. 8).
Espace intervertébral (DIV réséqué) Cartilage hyalin Os sous-chondral Plaque de croissance Os spongieux
Fig. 10 : Contact direct entre
espace médullaire (A) et CEP (B)
(d’après Nachemson et al71)
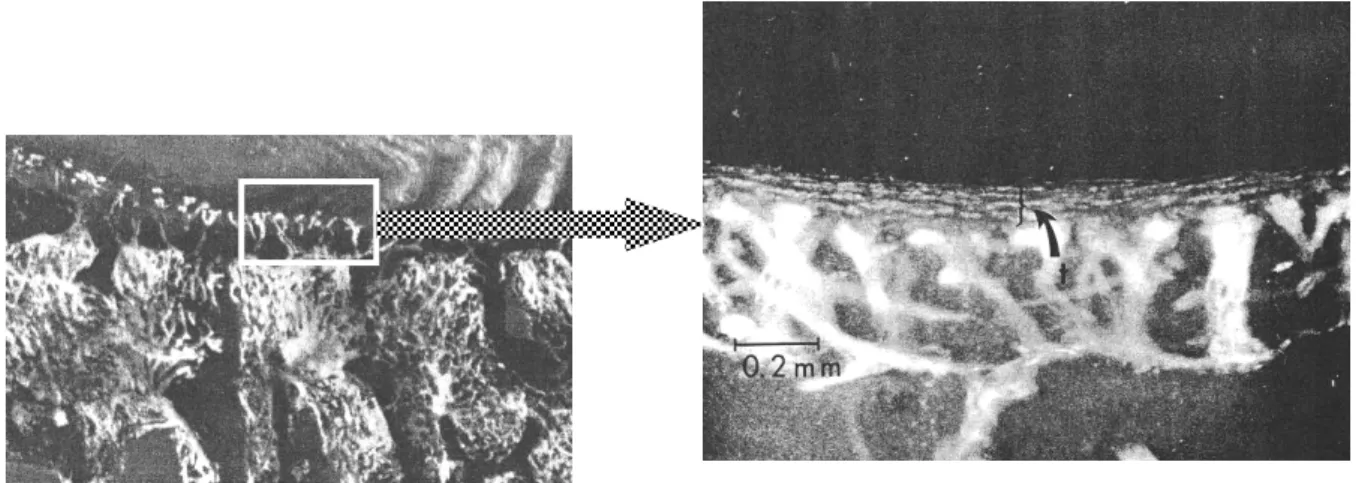
Il existe un lit capillaire à l’interface CEP-corps vertébral. Des terminaisons capillaires sessiles viennent au contact du cartilage et réalisent un véritable réseau plexiforme (Fig. 9).
Il existe également des zones de contact direct entre moelle osseuse et le CEP appelées "marrow contacts" 27 (Fig. 10). Les terminaisons capillaires ainsi que les zones de contact direct sont plus nombreuses dans la région centrale du plateau, en regard du NP, qu’à la périphérie, en regard de l’AF19,71,88,100. A la partie la plus périphérique de l’AF, ce réseau est moins riche voire inexistant par endroit avec des terminaisons moins nombreuses et plus grêles27. Oki et al.75 ont étudié la morphologie des bourgeons vasculaires en microscopie électronique au contact du CEP. Le nombre de vaisseaux en regard du NP n’était pas plus important qu’en regard de l’AF, mais ils présentaient plusieurs boucles (tels des ressort), offrant ainsi une surface d‘échange plus importante, tels les glomérules rénaux. Vaisseau sanguin Nucleus pulposus Annulus fibrosus
Fig.8 : Représentation de l’histologie normale (D’après Robert et al.88)
Disque intervertébral
Cartilage end-plate CEP calcifié Os sous chondral
On retrouve les mêmes éléments constituant le CEP vertébral et le cartilage articulaire, avec cependant un plus faible pourcentage d’eau pour le 1er90. Il n’existe pas de différence significative de composition entre les plateaux supérieurs et inférieurs ni entre les différentes vertèbres de L1 à S186. La matrice du CEP est composée d’un gel de Pg hydratés, renforcé d’un réseau de fibres de collagène et associé à d’autres substances inorganiques et organiques68. Le contenu du CEP en Pg est semblable à celui du DIV à savoir 0,2g pour 1g d’eau. Le contenu biochimique du CEP varie en fonction de la localisation88. Plus on se rapproche du corps vertébral, plus la quantité de Pg et la teneur en eau diminuent et plus la quantité de collagène augmente. Il existe une variation similaire dans le plan horizontal vers l’AF. Les fibres de collagène ont un diamètre de 300 à 800 Å avec des lacunes encore plus grandes; ainsi c’est le gel eau-Pg avec des pores très petits de 20 à 50 Å qui gouverne la porosité du milieu.
La composition biochimique du CEP est importante pour le maintien de l’intégrité du disque4,5,10,45,50,68,74,88,89,102, notamment les Pg50,74,89 qui jouent un rôle fondamental dans la régulation des échanges du DIV. Ainsi, quand la quantité de Pg diminue dans le CEP, elle diminue aussi dans le NP. Les Pg contrôlent la distribution des ions et des macromolécules, La diffusion des solutés à travers le CEP est directement liée à la concentration en Pg. Plusieurs auteurs ont impliqué la survenue de perturbations de la composition biochimique du CEP durant la croissance dans le développement de déformations scoliotiques77,87.
1.1.4 : Fonction & biomécanique
a- Disque intervertébral
La fonction principale du DIV est de maintenir un espace déformable entre les corps vertébraux offrant ainsi une mobilité mais aussi d’agir comme un amortisseur des forces compressives. Le DIV est plus résistant à la rupture que le corps vertébral ce qui explique la survenue de fractures sans lésions discales20.
Le comportement biomécanique d’un segment rachidien est hautement non linéaire. Ceci est grandement lié à la composition complexe du DIV qui montre des propriétés à la fois viscoélastiques et poroélastiques54. L’hydratation du DIV varie selon la charge à laquelle il est soumis. Ainsi les échanges liquidiens jouent un rôle majeur dans le comportement mécanique du DIV49.Après une nuit de repos, les activités de la vie quotidiennes entrainent une augmentation de la pression intradiscale du fait de la gravité mais aussi et surtout des contractions musculaires. Il en résulte une fuite d’eau du NP et donc une augmentation de la concentration en Pg. La concentration discale en Pg est à l’origine d’une "pression de gonflement" c’est-à-dire d’un appel d’eau par phénomène osmotique jusqu’à ce qu’un équilibre soit atteint. Ainsi, in vivo, une période de repos ou « de décharge » de 7 à 24 heures est nécessaire au DIV pour retrouver son contenu hydrique63 et sa hauteur67,85. La mesure de la pression intradiscale in vivo a été initiée par Nachemson dans les années 6070 puis confirmée et précisée plus récemment par Wilke et al.106. La pression chez un sujet adulte allongé est de l’ordre de 0,08 MPa, assis ou debout au repos de 0,50 MPa et jusqu’à 1,72 MPa en portant une caisse de 20kg. La valeur de pression communément admise pour une activité modérée est 1 MPa23.
b- Plateau cartilagineux
Il existe 2 grandes voies d’échanges liquidiens et d’apports nutritionnels entre DIV et circulation sanguine19,45,66 : (1) à travers les vaisseaux sanguins à la face périphérique de l’AF, (2) à travers le CEP par l’intermédiaire de bourgeons vasculaires à son contact (Fig.11).
Cette dernière représente la principale voie d’apport de nutriments du corps vertébral vers le DIV19,35,71,93. Si elle est compromise, cela peut entrainer la mort cellulaire du NP69,83. Parallèlement, la perméabilité du CEP contrôle la fuite de catabolites et autres produits de dégradation de la matrice du DIV. Ses propriétés de transport dépendent de la composition de sa matrice et en particulier du contenu en Pg50,74,88,89. Le CEP prévient également la fuite de Pg de faible poids moléculaire15.
La pression absorbée par le DIV est redistribuée partiellement vers le CEP18,80. Des forces compressives importantes, en particulier si elles sont répétées, peuvent être à l’origine de lésions irréversibles du CEP. Il apparaît que l’intégrité du cartilage et de l’os sous-chondral, plutôt que le degré de dégénérescence discale détermine l’importance des lésions générées46. Le CEP empêche enfin la protrusion du NP dans l’os spongieux du corps vertébral80.
1.1.5 : Maturation, dégénérescence
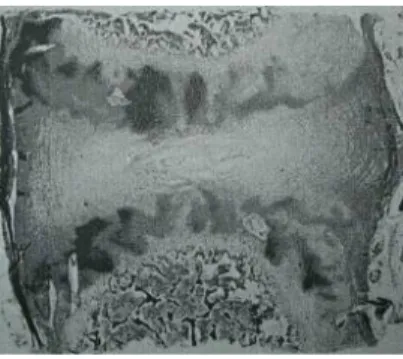
L’histologie et la composition du DIV et du CEP évoluent de manière importante après la naissance par une maturation progressive jusqu’à l’âge adulte (Fig. 12). Débute ensuite une dégénérescence en particulier au niveau lombaire, d’intensité variable selon les sujets.
Le CEP d’un nouveau né est composé d’une plaque de croissance et d’un cartilage articulaire bordant le DIV. Le cartilage de croissance est caractérisé par des colonnes parallèles de cellules prolifératrices et hypertrophiques. A la naissance, la densité cellulaire des chondrocytes augmente. Entre 3 et 10 ans, le nombre de vaisseaux diminue et ils s’oblitèrent progressivement. A l’âge de 10 ans, l’épaisseur du cartilage de croissance diminue ainsi que le nombre de cellules prolifératrices organisées de manière plus
irrégulières. A 18 ans le cartilage de croissance est calcifié. Après 45 ans, le cartilage calcifié est ossifié, les capillaires et espaces médullaires au contact du CEP sont presque totalement occlus. Le CEP est réduit après 65 ans à une mince couche de cartilage au contact du DIV14 (Fig. 13 et 14).
Fig. 11 : Les deux voies d’apport nutritif du DIV (d’après Holm et al.45)
Fig. 12 : Section parasagittale du corps vertébral d’un nouveau né. Le DIV a une forme biconcave. Le CEP et le cartilage de croissance sont très épais. D’après Donish et al.
Fig. 12c : 18ans
NAg NAg
Fig. 14 : Minéralisation progressive du CEP (d’après Bernick et al.10)
PAS NAg
Fig. 12a : nouveau-né Fig. 12b : 10ans NAg PAS
Fig. 12d : 35 ans NAg PAS
Fig. 13 : Evolution de la structure du CEP avec l’âge. A : cartilage articulaire (CEP), B : os sous-chondral, NP : nucleus pulposus, G : cartilage de croissance. (Grossissement x 50) (d’après Bernick et al.10)
Fig. 12e : 65 ans
NAg PAS
La composition biochimique du CEP et du DIV évolue avec l’âge parallèlement à ces modifications structurales. A l’âge adulte, la différenciation entre NP et AF est moins évidente, on parle alors de complexe central composé du NP et de la partie centrale de l’AF et de complexe périphérique constitué de la partie périphérique de l’AF qui s’insère sur le listel marginal du corps vertébral. La dégénérescence discale est caractérisée par une diminution des Pg agrégés et une augmentation des métalloprotéases lysant le lien protéique qui unit et stabilise l’acide hyaluronique au corps du Pg51. Il s’ensuit une perturbation du système osmotique discal et par conséquent une perte en eau et en éléments nutritifs. Cette déshydratation s’accompagnerait de mouvements discaux excessifs entrainant des microfissures qui augmenteraient la concentration discale en collagène. La dégénérescence discale est appréciée par la classification anatomopathologique de Nachemson106 :
1 : NP nettement individualiséet sain
2 : Minuscules ruptures du NP qui est moins bien délimité 3 : NP fibreux, non plus distinct de l’AF. Fissures de l’AF
4 : Larges fissures et cavités intéressant NP et AF. Présence d’ostéophytes sur les corps vertébraux adjacents.
Si l’âge est la cause principale de la dégénérescence discale, l’application d’une contrainte
mécanique est aussi un facteur déterminant qui entraine l’apoptose ou mort cellulaire au sein du CEP59,60.
1.2 : Considérations sur les transports convectifs dans le segment vertébral
1.2.1 : Comportement d'un milieu poreux élastique
a - Relation contrainte, déformation, pression de fluide
Le milieu poreux élastique déformable peut être modélisé par la superposition spatio-temporelle d'une matrice solide poreuse et d'un fluide la saturant (Fig. 15)13,26.
La mise en équation de chaque état (non développée ici) et leur superposition, permettent de mettre en place la loi de comportement d'un élément de volume macroscopique du milieu poreux. Comme le montre l'équation (1), le champ de contraintes total σ résulte d'une combinaison linéaire entre les déformations observables ε de la partie solide (squelette poreux) et la pression p du fluide. Les paramètres E, ν, K sont les propriétés mécaniques du
=
+
contrainte effective σ dans le domaine solide
pression p dans le domaine fluide
contrainte effective nulle dans le domaine solide pression p dans le domaine fluide
contrainte effective σ dans le domaine solide
pression nulle dans le domaine fluide Fig. 15 : Superposition des états de comportement mécanique.
milieu poreux : module d'élasticité, coefficient de Poisson et coefficient de compressibilité. Cette relation peut être écrite sous la forme (2) qui permet de faire apparaître le coefficient de Biot noté b, G étant le module de cisaillement.
(
)(
)
(
)
0 0 0 0 0 0 I 0 1 I 1 2 1 1 s E tr E K p Kυ
σ
ε
ε
υ
υ
υ
⎛ ⎞ ⎜ ⎟ ⎝ ⎠ = + − − − + + (1) 0 0 2 2 0 3 I I G K tr G b pσ
⎛⎜ − ⎞⎟ε
ε
⎝ ⎠ = + − (2)Sous l'hypothèse d'incompressibilité de la matrice solide, le coefficient Ks tend vers l'infini et par conséquent b tend vers 1. Cette hypothèse est souvent utilisée pour la modélisation des structures biologiques et implique que le milieu poreux se déforme sous l'action des gradients de pression.
b - Relation débit, pression de fluide, perméabilité
La loi de comportement reliant la vitesse de filtration qf dans un milieu poreux saturé au
gradient de pression p, est la loi empirique de Darcy exprimée par la relation (3). Le coefficient κ est la perméabilité intrinsèque ou spécifique (m2) et μ est la viscosité du
fluide (Pa.s). On peut définir également la perméabilité relative κ/μ (m4/Ns). grad f q p μ
κ
= − uur uuuuur (3) La perméabilité κ est une propriété intrinsèque du milieu poreux et elle est donc pardéfinition indépendante des propriétés du fluide et du procédé de mesure. Ce paramètre est principalement dépendant des caractéristiques géométriques du squelette poreux dont les paramètres les plus représentatifs peuvent être résumés à la porosité, le type et la topologie de leurs connexions (tortuosité équivalente unidimensionnelle). Le développement de modèles prédictifs de perméabilité à partir de la connaissance de la topologie des pores demeure un problème scientifique ouvert31.
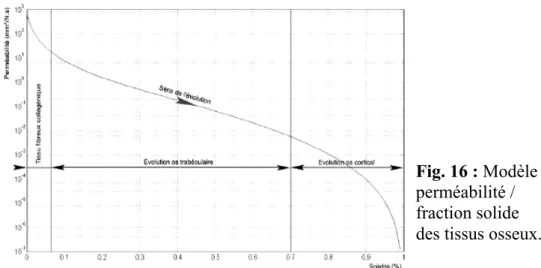
Dans le cas des applications au tissu osseux, on peut relever assez peu de travaux dans ce domaine. On peut noter toutefois certains résultats intéressants et notamment ceux de Arramon et al.6 établissant une loi empirique basée sur la porosité instantanée locale et reprenant l'approche de Carman-Kozeny basée sur des modèles capillaires22.La mesure de la porosité et de la perméabilité d'échantillons osseux (humains) a permis d'établir la loi de comportement (4) qui est représentée Figure 16.Ici la loi est établie en fonction de la fraction solide φs du milieu poreux (φs=1- φf); φf: étant la fraction fluide correspondant à la porosité
saturée par le fluide.
( )
104 2 B A Sφ
κ φ
= avec S=323 939 1340 1010 288φ
−φ
2+φ
3−φ
4+φ
5 (4) et A = 2.6 10-5 , B = 4.671.2.2 : Application au plateau vertébral, implication dans la
physiologie rachidienne
Les solutés dissous peuvent être transportés dans et à travers la matrice du DIV par un mécanisme passif de diffusion régi par des gradients de concentration des métabolites, indépendamment d’échanges de liquide2,10,19,45,50,52,65,66,68,71,73,74,81,93,100. Les corps dissous peuvent également être transportés au cours des échanges de fluides "pompés" à l’intérieur et à l’extérieur du disque lors des mouvements ou de la mise en charge du rachis. Ce mécanisme actif, appelé "bulk-flow" ou convection 19,35,45,52,53,65,66,68,71,89,93,99,100, est dépendant d’échanges liquidiens. Un faible volume liquidien est mobilisé lors des modifications de charge physiologiques au niveau des disques : moins de 0,01 % du volume discal est échangé pendant un cycle de marche et on estime que 3 à 20 % du volume total du disque est échangé au cours d’un cycle nycthéméral. Ces échanges liquidiens sont le fait de la compression en position debout ou assise qui chasse le liquide hors du DIV et de la « dépression » lors de la position allongée qui à l’inverse crée un afflux de liquide19,35,100. On décrit ainsi de petits échanges oscillatoires au cours des mouvements et de la marche qui se surajoutent aux échanges plus importants au cours du cycle nycthéméral. Ferguson et al35. ainsi que Sélard et al.93 ont étudié l’influence des modifications de la perméabilité du CEP sur la composition discale. Ils ont démontré que des perturbations de cette perméabilité influencent les transports moléculaires aussi bien par diffusion que par convection, compromettant ainsi le métabolisme cellulaire par défaut d’apport en nutriments. La mort cellulaire qui en résulte avec risque de dégénérescence discale peut retentir sur la fonction mécanique du DIV93.
La voie principale de nutrition du disque intervertébral se fait au travers du CEP19,35,45,71,93,100, comme l’ont démontré des études colorimétriques. Ferguson et al.35 ont montré que si on réduit la perméabilité du cartilage end-plate à 0, on constate une nette diminution du flux liquidien de 40 à 54 % alors qu’une perméabilité nulle en périphérie de l’AF ne réduit le flux que de 13 à 20 %. Selard et al. en 200393 ont développé un modèle en éléments finis de la diffusion de trois molécules (oxygène, glucose et lactate) à l’intérieur du DIV humain. Ils ont ainsi pu montrer que la concentration en corps dissous du NP est surtout sensible aux conditions aux limites au niveau du CEP et très peu au niveau de l’AF. La perméabilité du CEP a donc un rôle primordial dans les apports nutritionnels aux cellules discales et dans l’élimination des catabolites de la matrice88,89. Le transport de ces molécules dépend de nombreux facteurs liés aux vaisseaux (nombre, surface de contact)10,27,50,71, aux solutés (poids moléculaire, forme, charge)2,50,64,66,74,81,93,99,100, et au CEP (calcification4,40,79,
Fig. 16 : Modèle
perméabilité / fraction solide des tissus osseux.
composition en Pg50,74,88,89). La composition du CEP est variable en fonction de la région, centrale ou périphérique. De même, sa perméabilité n’est pas uniforme.
1.2.3 : Etude qualitative du transport convectif
Brodin19 le premier a étudié dès 1955 la perméabilité du CEP en injectant du fluorochrome en intraveineux à des lapins de 4 semaines. Il a ainsi mis en évidence les 2 voies possibles d’échanges entre DIV et circulation sanguine. La partie centrale du CEP, en regard du NP, était nettement plus perméable au colorant que sa partie périphérique. Ces résultats ont été confirmés sur des plateaux humains in vitro en 1970 par Nachemson et al.71 qui ont également observé que la perméabilité était liée à la présence d’espaces médullaires et de bourgeons vasculaires au contact du CEP27,53,66,71,88,89. Ces zones sont plus abondantes dans la région centrale des plateaux en regard du NP et diminuent vers la périphérie en regard de l’AF central. Elles sont quasi-inexistantes à la partie la plus périphérique des plateaux19,71,88,100. Environ 7 à 10% de la surface totale du CEP sont tapissés d’espaces médullaires directement à son contact et 20 à 30% soit directement à son contact, soit séparés du CEP par une épaisseur d’os sous chondral inférieure à 25µ66. Ces espaces étaient plus nombreux en regard du NP. L’examen microscopique de la partie latérale des plateaux vertébraux retrouvée imperméable a montré que le colorant était arrêté au niveau de la zone calcifiée du cartilage. Cette zone constitue une barrière pour le colorant comme elle pourrait l’être pour d’autres molécules71,100. L’étude in vitro de la perméabilité au glucose au niveau de plateaux vertébraux lombaires humains a montré une différence significative entre la partie centrale (0,58.10-6 cm2/s) et la partie latérale (0,18.10-6 cm2/s).
Urban et al.100 ont utilisé la diffusion de traceurs radioactifs pour montrer que 85% de l’interface DIV-corps vertébral est effectivement perméable au niveau du NP, 35% en regard de l’AF central alors que l’AF périphérique est pratiquement imperméable. Ces valeurs calculées sont corrélées avec les résultats histologiques. Il n’y avait pas de différence entre plateaux vertébraux supérieurs ou inférieurs. Ce sont surtout l’épaisseur de la couche de cartilage calcifié et le nombre de vaisseaux et d’espaces médullaires qui semblent gouverner la perméabilité du CEP93.
Ayotte et al.8 ont étudié l’influence de la direction du flux liquidien sur la perméabilité de CEP de moutons. La pression hydrostatique induite par les contraintes lors d’activités quotidiennes est de l’ordre de 0,6 à 1 MPa alors que la pression osmotique « de gonflement » du DIV est d’environ 0,15-0,2 MPa. Cette dernière est plus faible que la pression hydrostatique, alors que la période de repos (environ 8 heures) est plus courte que les périodes d’activité (environ 16 heures), le DIV retrouve une hydratation normale. Il semble donc qu’il existe une influence de la direction du flux sur les résistances à l’écoulement8. Ayotte et al. ont ainsi trouvé que la résistance à l’écoulement est près de 40 fois supérieure dans le sens DIV-corps vertébral (flow-out) que dans le sens corps vertébral-DIV (flow-in).
1.2.4 : Etude quantitative du transport convectif
Les études sur la diffusion au niveau des DIV, du cartilage articulaire ou des CEP sont nombreuses. Il peut s’agir de méthodes semi quantitatives grâce à des reconstructions IRM associées à l’injection de produits de contraste2,50,56,73,74,81, ou l’utilisation de colorants19,71, ou de solutés marqués53,89,99,100,102. Ces méthodes, si elles apportent des informations intéressantes ne permettent pas d’approcher quantitativement la perméabilité.
Gu et al.41, ont mesuré la perméabilité de l’AF dans différentes directions, et pour différents états de dégénérescence. Ils ont utilisé des rachis lombaires de cadavres humains avec comme étalon la mesure au niveau du cartilage articulaire de bovin (où k = 1.408 ±
0,135.10-15 m4/Ns). Cette valeur est cohérente avec celle trouvée par Mansour et al.64. Ils ont ainsi pu mesurer une perméabilité axiale de 1.530 à 1.640.10-15 m4/Ns, une perméabilité radiale de 1.675 à 1.924.10-15 m4/Ns et une perméabilité circulaire de 1.147 à 1.618.10-15 m4/Ns. Best et al.11, puis Houben et al.47 et enfin Iatridis et al.48, ont calculé par une méthode indirecte à l’aide de tests en compression confinée, la perméabilité radiale de l’AF de DIV humains ou canins et ont étudié son caractère anisotrope. Les valeurs retrouvées au cours de ces trois études sont très proches, respectivement 1 à 4.10-16 m4/Ns47, de 1,5 à 3.10-16 m4/Ns11, 1 à 2,3.10-15 m4/Ns48.
Néanmoins pour plusieurs auteurs7,48, il semble que de telles mesures ne soient pas très fiables et que l’on doive leur préférer des mesures « directes » comme le permet la manipulation originale de Gu et al41. Le dispositif comprend 2 chambres étanches, un vérin et un capteur de pression. Un échantillon d’AF de 6mm sur 0,3mm est inséré entre 2 plaques perméables de polyéthylène. Il est alors soumis à un flux liquidien continu et à une compression correspondant à une pression de 0,1 à 0,15MPa pour éviter la fuite de Pg. Le débit nécessaire pour maintenir une pression de 69kPa est utilisé pour calculer la perméabilité de l’échantillon.
La mesure directe de la perméabilité du CEP n’a, à notre connaissance, jamais été rapportée à ce jour.
1.3 : Considérations sur les approches expérimentales
1.3.1 : Modèle animal
Les rachis ovins et caprins sont souvent utilisés dans les études biomécaniques animales25,84,105,107, malgré des différences anatomiques et morphométriques. Wilke et al.105 ont comparé l’anatomie des rachis ovin et humain. Ils ont conclu de leur étude qu’il existe des similarités importantes entre les deux espèces en ce qui concerne les principales dimensions des vertèbres et les variations en fonction de l’étage concerné. Les étages thoraciques et lombaires représentent des modèles raisonnables de rachis humain. Au niveau lombaire, les corps vertébraux du mouton sont plus hauts que chez l’humain et les DIV moins épais. Ce même auteur a également réalisé des tests statiques sur banc de segments rachidien de mouton et a validé le modèle ovin pour la chirurgie discale mais aussi l’instrumentation rachidienne42,43,107. Une étude de la composition biochimique du DIV ovin a confirmé la validation du modèle84.
Ainsi même s’il s’agit d’un quadrupède ne présentant pas la même répartition des masses et n’étant pas soumis aux mêmes forces gravitationnelles qu’un bipède, l’ovin est un modèle animal fiable.
Le modèle porcin a quant à lui largement été utilisé dans la littérature76, y compris pour l’instrumentation rachidienne28,55,61. Les propriétés mécaniques de l’os porcin se rapprochent beaucoup de celles de l’humain1.
1.3.2 : Conservation des échantillons
Les manipulations nécessaires à la mesure de la perméabilité d’échantillons organiques demandent souvent beaucoup de temps. La conservation des tissus est donc une préoccupation majeure.
a - Etat frais
Dans leur étude sur les disques ovins, Costi et al.25 ont remarqué que les échantillons conservés dans de l’eau à 37°C commençaient à présenter des signes de décomposition après 4 à 5 heures. Si les propriétés mécaniques ont été peu modifiées jusqu’à la 8ème heure, elles se sont nettement dégradées à partir de la 20ème heure. De même, Rodrigo et al.91 ont montré que la viabilité des chondrocytes de cadavres conservés à 4°C sans milieu de culture chute rapidement de quasiment 100% à 6 h, à 80% à 12 h, 70 % à 24 h pour chuter ensuite plus rapidement. Malinin et al.62 ont montré que des lésions tissulaires n’apparaissent qu’après 20 jours de conservation à + 4°C dans un milieu de culture adapté.
Le Ringer Lactate (solution contenant des concentrations physiologiques de sodium, potassium, chlorure et bicarbonate) est fréquemment utilisé comme solution de conservation des échantillons osseux et cartilagineux. Pourtant il semble préférable de conserver ces échantillons à 4°C dans un milieu de culture stérile contenant les nutriments nécessaires au métabolisme des chondrocytes108. Ce milieu doit être renouvelé toutes les 48 à 72 heures ce qui permet un apport régulier de nutriments tout en assurant l’élimination des enzymes de dégradation des chondrocytes lysés. Wayne et al.104 ont pu conserver des échantillons ostéochondraux jusqu’à 60 jours à 4°C dans une solution de conservation changée deux fois par semaine17. Ils n’ont pas trouvé de différence significative des propriétés mécaniques ou biochimiques (quantité et distribution des Pg). Ils ont montré par ailleurs que la conservation dans un milieu de culture à 37 °C induisait des lésions tissulaires. Williams et al.108 ont confirmé ces résultats mais une différence significative du nombre et de la densité de chondrocytes viables apparaissait dès après 14 jours de conservation. De la même façon, ils n’ont pas retrouvé de différence significative concernant les propriétés mécaniques (perméabilité, module d’élasticité) entre les échantillons conservés 7 ou 28 jours, laissant supposer que le délai d’attente n’a pas induit de lésions tissulaires importantes. Il ressort de l’analyse de l’ensemble de ces études qu’il est raisonnable de conserver des prélèvements ostéocartilagineux 14jours dans un milieu adapté à 4°C.
b - Congélation
On retrouve des résultats contradictoires quant à l’influence de la congélation des échantillons sur les résultats des expérimentations dans la littérature8,9,108. L’origine des différences retrouvées n’est pas encore bien élucidée. Il semble que la congélation produise des dommages tissulaires soit directement par la formation de cristaux de glace, soit indirectement par la mort cellulaire8,9,91,97,98,104. Ces cellules relarguent dans la matrice des enzymes qui détériorent les Pg du cartilage. Ainsi, après congélation, on retrouve dans le cartilage une augmentation de la proportion des Pg de petite taille et une diminution de la quantité des Pg agrégés9. Ces altérations sont similaires à celles retrouvées au sein des disques de sujets âgés.
Williams et coll.108 ont montré que la congélation à -80°C entraine la mort des
chondrocytes. Ces observations ont été confirmées par les travaux de Enneking et Campanacci33,34. Bass et al.9 ont étudié l’effet de la congélation sur les propriétés biomécaniques de DIV de porc. Deux groupes d’échantillons ont été différenciés. Le premier était constitué de spécimens conservés à + 4°C. Les échantillons du deuxième groupe étaient congelés à -20 °C sans cryoprotecteur pendant 6 à 14 jours puis étudiés après décongélation rapide à température ambiante. Ils ont ainsi montré que la congélation modifie de façon très importante les propriétés mécaniques des échantillons; la perméabilité des disques congelés était 82% plus importante que celle des échantillons frais et la pression de gonflement des disques 25% plus faible. De plus, lors d’une série de 5 mesures consécutives sur un
échantillon congelé, même si la première mesure est proche de celle retrouvée sur le spécimen frais, les autres divergent et ce d’autant plus que l’on répète les cycles, avec des valeurs de plus en plus importantes. Ceci confirme que cette différence de comportement est certainement secondaire à des lésions tissulaires.
Ainsi, la majorité des auteurs s’accordent sur l’utilisation de cryoprotecteurs ou des techniques de congélation plus perfectionnées62,91,92,97,98,104,108 qui évitent la formation de
cristaux de glace et préviennent le choc osmotique induit par la déshydratation cellulaire lors de la congélation. Tomford et al.98 ont étudié la toxicité des cryoconservateurs. Le glycérol est plus toxique que le DMSO (DiMéthylSulfOxyde). Le meilleur taux de survie est obtenu avec une phase de pré-congélation par immersion des spécimens dans une solution de DMSO de 4 à 12 % pendant quelques minutes suivie d’une diminution lente de la température qui permet l’établissement d’un équilibre des milieux extra et intracellulaires, évitant ainsi les lésions cellulaires62,91,92,97,98,104,108. Il n’a pas été prouvé qu’une congélation à -196°C soit plus efficace qu’à -80°C92.
En ce qui concerne la décongélation, les auteurs pratiquent pour la plupart une décongélation rapide à température ambiante, dans une solution de Ringer (avec ou sans DMSO) à laquelle sont ajoutés des inhibiteurs enzymatiques41,92,108. Ceci semblerait améliorer les résultats en évitant la dégradation de la matrice cartilagineuse par les enzymes relargués lors de la mort cellulaire.
Néanmoins, ces notions ne sont applicables qu’aux chondrocytes isolés. Le problème semble plus complexe pour les pièces anatomiques car intervient la matrice extra-cellulaire qui constitue un obstacle à la diffusion du DMSO98 avec pour conséquence des taux de survie
inférieurs92. Une étude colorimétrique a cependant montré que le DMSO peut diffuser dans des échantillons ostéochondraux de taille conséquente en moins de 2 heures97. Ainsi, ce protocole de congélation semble néanmoins acceptable pour des échantillons de petite taille de l’ordre de 5mm. Gu et al.41 ont conservé leurs segments lombaires à -20°C avant de procéder aux mesures sur les échantillons et leurs résultats n’en ont pas été altérés. Notons que dans une étude de 1998, Gleyzes et al. ne trouvaient aucune influence de la congélation sur les propriétés biomécaniques de DIV d’agneaux37.
1.4 : Synthèse
Cette première partie a permis de développer une description anatomique, biologique et fonctionnelle du disque intervertébral. Le rôle des transferts de masse dans le segment intervertébral et notamment dans les problèmes de calcification des plateaux a été abordé. Les transferts de fluide, de protéine et de nutriments sont liés à la superposition complexe de phénomènes osmotiques et de convection - diffusion. Il apparaît donc que ces interactions entre transfert et comportement sain et pathologiques du rachis constituent actuellement un problème scientifique et clinique totalement ouvert.
Notre démarche a été focalisée sur l'analyse des propriétés convectives des tissus et plus particulièrement de la perméabilité. Les données de la littérature, notamment en termes de propriétés des tissus biologiques, sont relativement dispersées car associées à des modèles et à des techniques expérimentales variées. De plus elles sont souvent éloignées de notre problématique clinique première qui concerne le rachis en croissance et la scoliose idiopathique.
Cette étude bibliographique a donc permis de mettre en évidence l'intérêt de mener des études quantitatives des perméabilités de l'interface disque - vertèbre dans un cadre scientifique rigoureux notamment en utilisant des modèles animaux en croissance et des techniques expérimentales contrôlées.
CHAPITRE 2 : Développement d’un dispositif de mesure de la
perméabilité du plateau vertébral; application à un modèle animal porcin
2.1 : Objectif
L’objectif a été de mettre en place une procédure théorique et expérimentale de mesure de la perméabilité macroscopique d'échantillons cylindriques prélevés dans le plateau vertébral. Après validation du dispositif expérimental nous avons mené une étude pilote sur un modèle animal porcin.
2.2 : Matériel et méthodes
2.2.1 : Echantillons biologiques
Les échantillons biologiques ont été prélevés sur un cochon âgé de 4 mois. Une dose d’héparine de bas poids moléculaire (Innohep® tinzaparine 4500UI/ml, Léo Pharma) a été injectée en sous cutané les 2 jours précédents le prélèvement. Cette anticoagulation avait pour but de prévenir la formation de caillots sanguins au sein des échantillons et donc un biais potentiel de mesure de leur perméabilité57,103. Sous anesthésie générale et analgésie adaptée, les corps vertébraux de L4 et L5 ont été prélevés en bloc, puis l’animal a été sacrifié. Les 2
plateaux vertébraux de chaque vertèbre ont été découpés à scie oscillante avec une épaisseur de 6 mm. Le DIV a été réséqué à l’aide de bistouris ophtalmologiques sous microscope57,71. Cinq échantillons cylindriques de 3,5 mm de diamètre ont alors été prélevés dans le plateau caudal de L4 et le plateau crânial de L5 à l’aide d’un emporte pièce (Fig. 17a). Chaque
échantillon était donc constitué d'une séquence d'empilement de tissus biologiques : CEP, os sous chondral, os spongieux assimilable à une épiphyse et cartilage de croissance.
La hauteur des échantillons a été contrôlée par l’utilisation d’un guide de coupe pour la scie oscillante. La précision des coupes a été de 0,2 mm avec une marge d’erreur sur un échantillon de 6 mm de 0,2 / 6 ≈ 3 %.
Les échantillons (Fig.17b) ont été placés dans un tube en silicone (longueur l, diamètre intérieur ri, diamètre extérieur r0) (Fig. 17c), identifiés et immergés dans une solution de
(c)
(b)
Fig.17 : Plateau vertébral (face discale).
(a) 5 prélèvements cylindriques (notés S1 à S5) ont été effectués. Chaque échantillon
cylindrique (b) était introduit dans un tube en silicone (c). Les 9 coupes de l’étude microscopique sont visualisées en (a) (pointillés)
chlorure de sodium isotonique héparinée à 100UI/ml. Ils ont été conservés à 6°C et toutes les mesures ont été effectuées dans les 36 heures suivant le prélèvement.
2.2.2 : Considérations théoriques
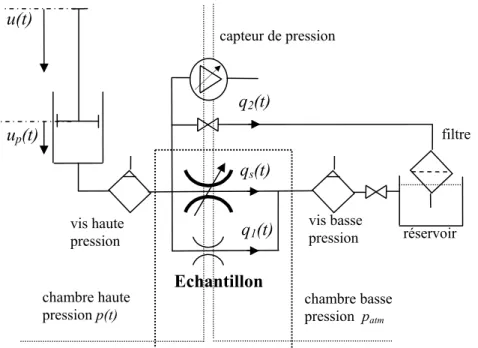
Le schéma hydraulique du dispositif expérimental est présenté Figure 18. Un débit de fluide transitoire était généré dans le circuit et en saturant totalement l'échantillon biologique, il permettait de déduire la perméabilité macroscopique à partir de la mesure de pression relative. Le déplacement contrôlé du vérin était quasi statique et l’effet de masse du à l’accélération du liquide négligeable.
L’équation (5) exprime la conservation de la masse liquide dans la zone à haute pression (Fig. 18). Le produit de la section du piston sp et de sa vitesse de déplacement u&preprésente le
débit généré dans la chambre à haute pression.
0 = − + + 1 2 p p f 0 p q q s u B v & & (5)
Le débit du à la compressibilité du liquide Bf en transformation isotherme est dépendant
dep&, la variation de pression dans le temps et de v0, la vitesse moyenne du fluide. Les
variables qs, q1 et q2 représentent respectivement le débit à travers l’échantillon, les fuites
autour de l’échantillon et les fuites de la durite.
Le débit est laminaire avec un faible nombre de Reynolds. Le débit à travers l’échantillon (section ss, longueur ls) dépend de la pression relative p, de la perméabilité intrinsèque κs et de
la viscosité du liquide μ, en appliquant la loi de Darcy (6a)31. De la même manière, les fuites sont proportionnelles à la pression différentielle en utilisant les coefficients λ1 et λ2 selon (6b)
et (6c). p l s q s s s s μ κ = (a) q1 =λ1 p (b) q2 =λ2 p (c) (6) La déformation des composants du montage mis en pression est prise en compte de manière globale dans l’équation (7) exprimant la différence entre le déplacement imposé u et
q2(t) qs(t) q1(t) capteur de pression filtre vis basse pression vis haute pression Echantillon réservoir chambre haute
pression p(t) chambre basse pression patm
up(t) u(t)
le déplacement effectif du piston up, k étant le coefficient de raideur linéaire globale du
montage (piston, durite…).
( )
p p
p s =k u u− (7)
L’introduction des équations (6a), (6b), (6c) et (7) dans l’équation (5) permet d’exprimer la pression relative p(t) par l’équation différentielle de 1er ordre, équation (8). La constante A représente la perte de charge due au passage du fluide à travers l’échantillon et aux fuites éventuelles (λ1, λ2). La constante B représente la compressibilité, Bf et v0 sont respectivement
la compressibilité du liquide et sa vitesse moyenne.
u B p τ p&+1 = 1 & (8) avec 1 A τ = B (a)
(
)
1 s s 1 2 p s p s A s l s κ λ λ μ = + + (b) 2 1 0 p p f s v B s B k ⎛ ⎞ = ⎜⎜ + ⎟⎟ ⎝ ⎠ (c)L’équation (8) montre que la variation de pression dépendait de la variable u&. La loi
cinématique du vérin comprend donc 2 phases principales (Fig. 23). Au cours de la phase 1, Le piston se déplace à vitesse constanteu&entre 0 et t1. La pression p1(t1) est atteinte et la
solution de l’équation (8) est exprimée par (9). La phase 2 débute en t1, la vitesse u&est nulle et
le déplacement u constant. La relaxation de la pression amont à partir de la pression p1(t1) est
exprimée par l’équation (10).
(
τ)
κ1 t 1 e u t p ( )= & 1+ − avec[
]
1 1 t t u u& = & ∀ ∈ 0 ; (9)(
t τ)
tτ 1 2 e e u t p ( )= 1+ 1 −κ& avec u&=u&2 =0 ∀t ∈
[
t1 ; +∞[
(10)L’acquisition de la pression au cours du temps est représentée par le couple (ti, pi). Le
paramètre τ est obtenu en utilisant une approximation par la méthode des moindres carrés appliquée à l'exponentielle décroissante (10), comme exprimé dans l’équation (11). En introduisant l’équation (8b) dans (10) on obtient la perméabilité de l’échantillon, et celle-ci est exprimée par la relation (12).
L’utilisation préalable d’un échantillon non poreux (κs = 0) et complètement imperméable
permet de déterminer par les équations (11), (12) et (8a) la perméabilité du dispositif (λ1 +λ2)
et son coefficient de compressibilité B, tous les autres paramètres étant contrôlés.
∑ −∑ ∑ ∑ − ⎟ ⎠ ⎞ ⎜ ⎝ ⎛ ∑ = = = = = = n i n i i n i i i i n i i n i i p t p t n t n t 1 1 1 1 2 2 1 ln ln τ (11)
(
)
2 2 2 1 1 1 1 1 1 1 ln ln 1 2 1 n n n n n n i i i i i i i i i i i i i t t p t p n t t p s p t s 1 s s l s u e e s τ κ μ λ λ = = = = = = ⎛ ⎞ ⎛ ⎞ ⎜ ⎛ ⎞ ⎟ ⎜ − ⎟ ⎜ −⎜ ⎟ ⎟ ⎜ ⎟ ⎜ ⎜ ⎟ ⎟ ⎝ ⎠ ⎝ ⎝ ⎠ ⎠ ⎡ ∑ ∑ ∑ ∑ ∑ ∑ ⎤ ⎛ ⎞ ⎢ ⎥ =⎢ + −⎜ ⎟⎥× + ⎝ ⎠ ⎢ ⎥ ⎣ ⎦ & (12)2.2.3 : Dispositif expérimental
Le dispositif expérimental correspondant au schéma hydraulique est présenté Figure 19. Le fluide utilisé était un chlorure de sodium isotonique (Baxter®) à température ambiante (22° +/-10%). La pression d’amont était contrôlée par un vérin hydraulique (Joucomatic®, diamètre 10 mm, course 10 mm, section 804,25 mm2) actionné par une machine d'essai électromagnétique asservie en déplacement (Elf 3200 Enduratec®). Durant la phase 1 ( u=u t & &1 ∀ ∈
[ ]
0;t1 ), un débit constant de 2mm3/s était généré pendant 120 s pour atteindre une pression d’amont p1 (t1) de 1Mpa. Le vérin était immobilisé dans cette position pendant 1080
s représentant la phase 2 ( u=u = t & &2 0 ∀ ∈
[
t1;+∞[
). L’évolution de la pression d’amont était mesurée par un capteur (TME®, 0-5bars) et elle était enregistrée par le logiciel Labview® (National Instruments). Le fluide traversait l’échantillon saturé et la perméabilité macroscopique était obtenue à partir du post-traitement de l’évolution de la pression en phase 2 (équation 12). Le fluide rejoignait par la suite, le réservoir basse pression (pression atmosphérique).L’échantillon biologique préalablement glissé dans le tube silicone (§ 2.2.1), était introduit dans une entretoise en aluminium d'alésage intérieur très légèrement conique. Une deuxième entretoise de serrage, plus étroite était mise en place (Figure 2) et le tube silicone était soigneusement
u specimen 1 silicon tube 2 z r θ w0 ri ro l ELF 3200 Enduratec® échantillon joint torique tube silicone entretoise conique entretoise de serrage vérin réservoir
(b)
(c)
capteur de pression(a)
Fig.19 : (a) Dispositif de mesure. (b) Positionnement de l’échantillon dans la chambre étanche. (c) Modélisation de l’échantillon.