UFR Sciences de la vie
THESE DE DOCTORAT
Discipline : Immunologie
Présentée et soutenue par
Michael ESQUERRE
Le 29 novembre 2007
En vue de l’obtention du grade de
DOCTEUR DE L’UNIVERSITE DE TOULOUSE III
INFLUENCE DES LYMPHOCYTES T CD4
+CD25
+REGULATEURS
SUR LA DYNAMIQUE DE FORMATION DE LA SYNAPSE
IMMUNOLOGIQUE ENTRE UN LYMPHOCYTE T CD4
+EFFECTEUR
ET UNE CELLULE PRESENTATRICE D’ANTIGENE
Jury
Pr Joost van Meerwijk
Président
Dr Ignacio Anegon Rapporteur
Dr Claire Hivroz
Rapporteur
Dr Alain Trautmann
Rapporteur
Dr Denis Hudrisier
Examinateur
Au Dr Paulette Recco,
je dédie ce travail
Toulouse le 18 novembre 2007,
Quel difficile exercice que de synthétiser en deux ou trois pages seulement les remerciements que je voudrais adresser aux différentes personnes que j’ai rencontré et apprécié durant ma vie de thésard ainsi, qu’aux personnes que je connais et qui m’entoure depuis bien plus longtemps que ça.
Après avoir enfin achevé la version finale de ce manuscrit, je vais tenter de satisfaire à cet exercice de mon mieux. Toutes mes excuses aux personnes que je pourrais oublier, mes pensées vont vers vous également. En ce début de soirée de dimanche de novembre, instant pourtant plutôt propice au regret de l’agréable week-end qui vient de se terminer et à la terreur de la sonnerie stridente du réveil qui retentira bien trop tôt demain matin, j’espère pourtant que l’inspiration va me gagner…
Je voudrais tout d’abord remercier le Pr Joost van Meerwijk d’avoir accepté d’être le président de mon jury de thèse. Vous avez été mon chef durant mon DEA et vous êtes également présent à la conclusion de mon parcours universitaire. Même si nos rapports ont souffert je pense d’incompréhensions, je tenais à vous dire que j’ai appris beaucoup de choses à vos côtés durant cette dure année qu’a été le DEA. Merci de m’avoir initié au monde du lymphocyte T régulateur.
Je voudrais remercier également mes trois rapporteurs : Claire Hivroz, Ignacio Anegon et Alain Trautmann qui, après avoir lu et analysé de manière chirurgicale mon manuscrit, ont tous répondu présents pour assister à ma soutenance. Veuillez me pardonner Mr Trautmann s’il reste encore quelques fautes d’accords. J’ai confié mon manuscrit à cinq personnes, à priori sans lacunes grammaticales déclarées, qui ont successivement traqué sans relâche la moindre faute et coquille. Un grand merci à toi aussi Denis pour avoir accepté d’être examinateur de ma thèse. Je suis vraiment heureux que tu sois présent le jour de ma soutenance. Tu as été pour moi le meilleur des enseignants que j’ai pu rencontré tout au long de mon parcours universitaire, tu m’as donné le goût de l’immunologie. J’ai pu un peu mieux te connaître durant mon DEA où, en plus de tes grandes qualités scientifiques, j’ai pu apprécier ton immense gentillesse, ton écoute, et je n’oublierais pas non plus tes performances à Altissimo !
Ensuite, j’en arrive logiquement à remercier « Il grande capo » alias « Il dottore Salvatore Valitutti ». Merci de m’avoir accepté dans ton équipe à la fin de mon DEA, de m’avoir fait confiance et de m’avoir donné les moyens scientifiques et matériels pour développer une thématique nouvelle dans ton laboratoire. Je me rappelle encore quand tu as écrit le sujet de ma thèse sur un morceau d’enveloppe. Finalement, après beaucoup d’efforts on est arrivé à en faire quelque chose de tout ce travail qui, je l’espère, sera bientôt publié. Ces trois années de thèse ont été très formatrices, je pense que j’ai pu développé mon indépendance scientifique mais aussi, ma capacité à remettre en cause mes résultats et à penser à faire le contrôle du contôle sans bien sur, oublier les témoins adéquats ! Je pense que l’on a appris à bien travailler ensemble, et je pense qu’on arrivera à produire de bonnes données scientifiques durant l’année qui vient. Comment en pensant à Salvatore ne pas penser à Sabina, merci pour m’avoir initier aux joies de la microscopie confocale, pour m’avoir donné un coup de main sur certaines manips mais également, pour m’avoir fait découvrir la vraie cuisine Italienne.
Maintenant mes pensées vont vers ma tutrice de monitorat, Cécile alias « la Mama » ! A la fac tu ne montres que ton côté obscur et oui, moi aussi tu me faisais un peu peur quand j’étais en DEUG et que tu criais dans l’amphi mais bon, les longues pauses café que tu faisais permettaient de compenser ! Merci de m’avoir aidé dans mon expérience d’enseignement. Merci pour ta bonne humeur au labo, les crises de fou rire qu’on a pu avoir dans le bureau ou même lors d’une surveillance d’exam ! Enfin, merci pour ta relecture de cette thèse. Je pense maintenant à la technicienne de choc du CPTP : Martine Guiraud. Rien ne te fait peur, te jeter sur un pakito, poser en petite tenue au sommet d’une montagne pour un calendrier ou bien faire une cinétique de calcium avec moi au LSRII ! Merci pour ton aide au labo et pour ta gentillesse, on a bien rigolé en se racontant des potins tout en cultivant nos cellules. Tu vas me manquer, je crois que je vais collaborer avec ta nouvelle équipe l’année qui vient. Merci Eric, pour ces bonnes tranches de rigolades en manip, pour toutes ces pauses café avec toujours le petit chocolat qui va bien où l’on a pu discuter de sciences et de pleins d’autres choses. Tu m’as aidé à chaque difficulté que j’ai pu rencontré durant ces trois années de thèse. Tu possèdes de grandes qualités scientifiques et tu es un collègue précieux. Merci aussi Eric pour avoir pris le temps de relire et de corriger ma thèse.
Je pense maintenant à celui qui a soutenu sa thèse avec les honneurs il y a maintenant presque deux mois, j’ai nommé Nico ! Merci pour tout mon cher Nico, mon accolyte au labo et dans les soirées ! Durant tout ce temps où on a travailler ensemble nous avons pu mieux nous connaître, parler de nos
doutes, de notre avenir, mais aussi bien se marrer et je pense au final devenir amis. Merci encore pour ton soutien durant cette année 2007 qui n’aura pas vraiment été facile pour moi.
Merci également à mon stagiaire de compétition, merci Baptiste pour ta bonne humeur et tes manips, je pense qu’on a fait une bonne équipe ensemble, tu es arrivé à un moment crucial dans ce travail. Je crois que tu es encore plus calme que moi…
Merci à toi Florie pour ta joie de vivre au quotidien, c’est un plaisir de travailler avec toi. Je pense d’ailleurs que l’on va travailler ensemble cette année à venir.
Enfin, après avoir passé en revue les membres toujours présents de l’équipe Valitutti, je n’oublie pas ceux qu’ils l’ont été, mais qui ne le sont plus. Je pense en premier lieu à mon ancien colocataire de bureau, mister David, merci pour ton aide lors de mon début de thèse. Tu m’as aidé à m’adapter à ma nouvelle équipe, à passer du monde de la souris C57BL/6 au monde des cellules humaines. Merci pour toutes ces discussions. Je pense également à Aurélie, merci pour ton sang chaud Perpignanais et pour toutes ces discussions où j’ai appris tant de choses, d’ailleurs, depuis que tu es partie, je ne suis plus au courant de rien. Merci également à toi Séb, tu as pris la place de David à la fois sur le projet de la FRAP et dans le bureau. Tu es vraiment quelqu’un de brillant. Les discussions vétos à propos de mon chat me manquent pas mal et en plus, depuis que t’es parti, j’ai plus de gateaux à 16h !
Je pense également aux membres de passage : Iris, la travailleuse acharnée et Leandrito, le plus sympa de tous les biologistes Chiliens.
J’ai une pensée également pour Christelle et Laurent qui ont partagé le même bureau que moi durant ma première année de thèse.
J’espère avoir pu vous donner un peu de tout ce que vous m’avez offert.
Je tiens également à remercier Loïc Dupré et toute son équipe, Ronan, Yovan, Gema, Fanny et Julie, sans oublier Josipa. Merci Loïc pour toutes les discussions que l’on peut avoir dans le bureau, pour tous tes bons conseils, tu m’as d’ailleurs bien aidé à stucturer la dernière partie de ce manuscrit. Mes pensées vont également à Abdel Saoudi, merci pour ton aide précieuse et tes conseils scientifiques avisés. Tu nous as bien épaulés tout au long de ce travail.
Enfin, je tiens à remercier les membres des autres équipes du département Immunologie avec qui j’ai partagé d’agréables moments. Je pense à l’équipe van Meerwijk, à Thibault et ses drôles de dames. Merci à Julie T. et Céline pour avoir repris le flambeau de la gestion du café, car comme chacun sait, au boulot : le café c’est le nerf de la guerre. Mes pensées vont également à Julie R., Ingrid et aussi Olive. Merci Olive pour ton encadrement, pour ton soutien, ta sympathie et ta disponibilité durant mon année de DEA. J’ai beaucoup appris grâce à toi. Depuis que tu es parti, Paris Match a déposé le bilan.
Mes pensées vont également aux membres de l’équipe Guéry, Cyril, Karine avec qui il est toujours agréable de discuter au moment du repas. Et puis je n’oublie pas Sophie également, dommage que tu ne sois pas là pour la soutenance.
Merci au membre de l’équipe Lebouteiller, je pense notamment à Julie pour ta bonne humeur et puis merci aussi pour avoir participé à la relecture de ce manuscrit. Je voudrais remercier Maryse également et pas que pour ses dons culinaires mais également pour m’avoir aidé avec les PCR Foxp3 quand j’essayais de savoir si mes Treg étaient bien des Treg !
Mes pensées vont également à l’équipe de Sylvie Guerder où il est toujours agréable d’aller discuter un peu de sciences ou d’autres choses. Merci Stéphane pour tes mails divertissants et pour les épisodes d’Heroes.
Je voudrais également remercier les personnes en charge des plateaux techniques, Fatima et Valérie pour la cytométrie et le tri cellulaire. Merci pour votre diponibilité et votre gentillesse. Vous m’avez bien aidé pour ces dernières expériences en cinétique de calcium sur le LSRII. Fatima, c’est toujours un plaisir de discuter avec toi, en plus j’apprends et dès fois même, je t’apprends quelques histoires dont tu n’as pas encore eu vent. Merci également à Sophie qui a la charge du plateau d’imagerie, merci pour ta disponibilité et ta gentillesse. Merci à vous toutes de préserver l’intégrité de ce précieux matériel afin de nous permettre à tous d’avançer dans nos travaux.
Je voudrais également remercier Aline, toujours pleines de douces attentions, quand Nicolas était là j’étais n°2, mais depuis que Nicolas est parti je pense que je suis passé n°1.
Merci également à Dominique pour ton sérieux et ton efficacité dans le transfert de nos commandes. Cela nous épargne ainsi beaucoup de travail administratif et nous rend la vie plus simple !
Je voudrais également remercier Yvan, Jacques, Nordine, Joël et Marie qui travaillent dans l’ombre mais qui permettent à la machine de tourner rond.
Mes pensées vont également à Rémy et Delphine. Rémy on a fait quasiment tout notre parcours universitaire ensemble. J’ai toujours été impressionné par ta capacité à comprendre si rapidement les choses, ça ne m’étonne pas que tu travailles maintenant sur des ADN polymérases bizarre. Si je devais me souvenir de quelque chose de toi, ce serait de ton imparable raisonnement et de ta capacité à te transformer en Raymond quand tu buvais un verre de trop dans les soirées, j’en rigole encore.
Merci à toi aussi, Cédric alias Zizouille, le plus grand n°10 du football que le monde est connu après le vrai Zidane. On a sympathisé durant notre licence et depuis, beaucoup de choses se sont passées. Tu es quelqu’un que j’estime beaucoup.
Je voudrais témoigner mon affection à Julien, tu es comme un frère pour moi. J’ai vraiment de la chance de t’avoir rencontré en 1992 au collège du château de l’Hers. Tu as été à mes côtés dans les bons comme dans les mauvais moments de ma vie. Il y aurait beaucoup trop de choses à raconter, merci pour tout ce que tu m’apportes.
Merci à tous mes amis : Charlotte, Biggy, Béa, Rom et les autres. C’est grâce à vous que j’arrive à garder mon équilibre au quotidien, et ma dose d’apéros le dimanche soir. Vous comptez énormément pour moi.
Je tiens également à remercier ma famille et surtout mes parents, vous m’avez toujours aidé, soutenu et apporté ce dont j’avais besoin. C’est grâce à vous si aujourd’hui j’en suis arrivé là, merci pour tout. Je voudrais témoigner mon affection à ma petite sœur Emma, qui n’est plus si petite que ça maintenant. Merci pour tout ce que tu as fais pour moi.
Enfin, mes plus tendres pensées vont à ma petite Camille, merci de m’aimer autant et d’être comme tu es. Merci de m’avoir soutenu jusqu’au bout, en relisant plusieurs fois cette thèse (je crois que tu seras la seule à avoir subi ça). Marcher à tes côtés dans cette grande aventure qu’est la vie est un privilège. Il n’y a pas de mots pour te dire au combien je t’aime et au combien je tiens à toi.
REMERCIEMENTS
SOMMAIRE
2
RESUME
5
ABSTRACT
6
LISTE
DES
FIGURES
7
LISTE
DES
ABREVIATIONS
9
AVANT-PROPOS
10
INTRODUCTION
13
LES LYMPHOCYTES T 14
I. Développement et sélection des lymphocytes T αβ
14II. Récepteur à l’antigène des lymphocytes T αβ
19• Structure du TCR
• Le complexe TCR/CD3
III. Organisation des gènes du TCR
21ACTIVATION DES LYMPHOCYTES T 23
I. Sensibilité et
spécificité
des
TCR
24
• Le
modèle
du
kinetic proof reading
• Le
modèle
du
serial triggering
II.
Activation
de
la
cellule
T
28
• Activation de la cellule T et Valence des TCR
• Le modèle de « kinetic segregation »
• Les microdomaines lipidiques
• Changements
conformationnels et engagement des TCR
• Activation des LT par des super antigènes bactériens
SIGNALISATION ET MISE EN PLACE DES FONCTIONS EFFECTRICES DES LT 40
I. Mise en place de la signalisation et des voies de transduction du signal
II. Fonctions effectrices des LT :
45• LT
CD4
+(TH1, TH2 et TH17)• LT
CD8
+BIOLOGIE ET POTENTIEL THERAPEUTIQUE DES LT CD4+ CD25+ REGULATEURS
I. Caractérisation,
développement
et homéostasie des LT régulateurs
(Treg)
52• Plus de vingt-cinq ans d’efforts à la recherche du marqueur…
• Développement et homéostasie du compartiment T régulateur
II.
Mécanismes
de
suppression
61• Nécessité
du
contact
cellulaire pour la suppression in vitro
• L’implication des cytokines dans la suppression
• Fonctions
physiologiques,
physiopathologiques et signalisation du
TGF-β1
• Autres mécanismes de suppression
III. Fonctions
physiologiques
et physiopathologiques des Treg
70• Tolérance
foeto-maternelle
• Modulation des réponses immunitaires anti-infectieuses
• Effet délétère sur l’immunité anti-tumorale
IV.
Potentiel
clinique
76
• Transplantation d’organes et de tissus
• Pathologies
auto-immunes
et immuno-inflammatoires
• Une aide à la thérapie génique
SYNAPSE IMMUNOLOGIQUE ET DYNAMIQUE DES INTERACTIONS LYMPHOCYTAIRES
I.
Architecture de la Synapse Immunologique
82• La synapse immunologique concentrique
• Diversité des synapses immunologiques
II.
Dynamique à la Synapse et polarisation des lymphocytes T
87• Dynamique de l’interaction lymphocyte T/CPA
• Polarisation des lymphocytes T
III. Synapse
Immunologique
et lymphocytes T CD4
+CD25
+régulateurs
92OBJECTIFS
DU
TRAVAIL
95RESULTATS
97DISCUSSION GENERALE ET PERSPECTIVES
102RESULTATS
PRELIMINAIRES
117
Michael Esquerré
Influence des lymphocytes T CD4+ CD25+ régulateurs sur la dynamique de formation de la synapse immunologique entre un lymphocyte T CD4+ effecteur et une cellule
présentatrice d’antigène
Directeur de thèse : Pr Salvatore Valitutti Toulouse, hôpital Purpan, le 29 novembre 2007
RESUME
La rencontre entre un lymphocyte T et une cellule présentatrice d’antigène (CPA) est un évènement central dans l’initiation et le développement de la réponse immunitaire adaptative. L’interaction entre ces deux cellules entraîne de nombreuses réorganisations moléculaires au niveau de l’aire de contact intercellulaire conduisant à la formation d’une structure dynamique et spécialisée remplissant diverses fonctions biologiques : la Synapse Immunologique (SI). Cette interaction permet à un lymphocyte T CD4+ helper (T
H) de s’activer et de mettre en place une signalisation intracellulaire nécessaire à la
production de cytokines. Le second aspect crucial de cette interaction consiste en la polarisation de la machinerie sécrétoire du lymphocyte TH vers la CPA permettant ainsi une activation sélective de la
CPA présentant l’antigène spécifique et donc une amplification sélective de la réponse immunitaire. Les lymphocytes T CD4+ CD25+ régulateurs naturels (Treg) jouent un rôle capital dans le maintien de la tolérance périphérique au soi, leur absence conduisant au développement de syndromes lymphoprolifératifs auto-immuns. Les Treg sont également impliqués dans le contrôle des réponses immunitaires anti-infectieuses et ont un rôle délétère lors des réponses immunitaires anti-tumorales. Différents mécanismes de régulation impliquant le contact cellulaire ou bien la sécrétion de molécules effectrices solubles ont à ce jour été décrits.
Mon travail de thèse a été de déterminer si les Treg humains pourraient inhiber les réponses immunitaires en altérant la polarisation des lymphocytes TH vers les CPA. Afin de répondre à cette
question, nous avons utilisé des approches de microscopie confocale afin de visualiser un Treg et un lymphocyte TH interagissant simultanément avec une même CPA. Nous avons pu observer que les
Treg inhibent la polarisation de la machinerie sécrétoire des lymphocytes TH (appareil de Golgi et
cytosquelette de tubuline) vers la CPA via la production locale de TGF-β. L’obtention de ces résultats nous a permis d’identifier un nouveau mécanisme de suppression, qui pourrait permettre de mieux appréhender l’incroyable potentiel des Treg à réguler finement les réponses immunitaires.
Mots clés : Synapse Immunologique, Lymphocytes T régulateurs, microscopie confocale, TGF-β.
Discipline : Immunologie.
INSERM U563, Centre de Physiopathologie de Toulouse Purpan, CHU Purpan, BP 3028, 31024 Toulouse Cedex 3
Michael Esquerré
Influence of CD4
+CD25
+regulatory T lymphocytes on the dynamic of the
immunological synapse formation between a CD4
+effector T lymphocyte and
an antigen presenting cell
Thesis supervisor: Pr Salvatore Valitutti Toulouse, Purpan hospital, November 29th 2007
ABSTRACT
The encounter between a T lymphocyte and an antigen presenting cell (APC) is a central event in the initiation and development of adaptative immune responses. Interaction between these two cells leads to multiple molecular reorganizations of the intercellular contact site leading to the formation of a dynamical and specialized structure filling diverse biological functions: the Immunological Synapse (IS). This interaction enables a CD4+ T helper lymphocyte (TH) to activate and to put into place an
intracellular sustained signaling necessary for cytokine production. The second key feature of this interaction consists in TH lymphocyte secretory machinery polarization towards APC thereby allowing
a selective activation of the APC presenting the specific antigen and thus a selective amplification of the immune response. CD4+ CD25+ natural regulatory T lymphocytes (Treg) play a pivotal role in the maintenance of peripheral self tolerance, their absence leading to the development of autoimmune lymphoproliferative disorders. Treg are also involved in controlling anti-infectious immune responses and have a deleterious role during anti-tumoral immune responses. To date, different regulation mechanisms involving cellular contact or the secretion of soluble effector molecules have been described.
My thesis work was to determine if human Treg could inhibit immune responses by altering polarization of TH lymphocytes towards APC. In order to answer this question we used confocal
microscopy approaches so as to visualize a Treg and a TH lymphocyte simultaneously interacting with
a same APC. We were able to observe that Treg inhibit secretory machinery polarization of TH
lymphocytes (Golgi apparatus and tubulin cytoskeleton) towards APC via local TGF-β production. These results enabled us to identify a novel suppression mechanism that could allow to better apprehend the incredible potential of Treg to finely regulate immune responses.
Keywords: Immunological Synapse, regulatory T lymphocytes, confocal microscopy, TGF-β.
Discipline: Immunology.
INSERM U563, Centre de Physiopathologie de Toulouse Purpan, CHU Purpan, BP 3028, 31024 Toulouse Cedex
Figure 1 Engagement vers le lignage lymphocytaire T
Figure 2 Principales étapes du développement thymique des lymphocytes T
Figure 3 Induction de tolérance centrale et périphérique, remparts face à l’autoimmunité Figure 4 Architecture moléculaire du complexe TCR/CD3
Figure 5 Diversité combinatoire et jonctionnelle des TCR
Figure 6 Schématisation du modèle de « kinetic proofreading » pour la signalisation du TCR Figure 7 Le « serial triggering », engagement séquentiel des TCR
Figure 8 Modèle d’hétérodimérisation du TCR Figure 9 Modèle de pseudodimérisation des TCR Figure 10 Modèle des TCR multivalents
Figure 11 Modèle de « kinetic segregation »
Figure 12 Comparaison cristallographique des complexes TCR/CMH/SAg (SEB) et TCR CMH/peptide Figure 13 Voies de transduction du signal en aval du TCR
Figure 14 La voie calcique, signal clé de l’activation lymphocytaire T Figure 15 Un même précurseur, plusieurs destins
Figure 16 Plusieurs marqueurs pour identifier une même population cellulaire Figure 17 Développement intra-thymique du lignage T régulateur (Treg) Figure 17.b Voie de signalisation induite par le TGF-β1
Figure 18 Mécanismes de suppression des Treg
Figure 19 Balance entre lymphocytes TH et Treg : Immunité anti-infectieuse protectrice versus
prévention d’immunopathologie
Figure 20 Les Treg : obstacle majeur à l’immunosurveillance anti-tumorale Figure 21 Architecture moléculaire de la synapse immunologique concentrique Figure 22 Diversité des synapses immunologiques
Figure 23 Dynamique de l’interaction lymphocyte T/CPA
Figure 24 Les Treg inhibent la polarisation de la machinerie sécrétoire des lymphocytes TH via un
mécanisme TGF-β dépendant.
Figure 25 Schématisation du potentiel des Treg et du TGF-β à inhiber la sécrétion cytokinique polarisée au niveau du microenvironnement tumoral
ADN Acide désoxyribonucléique AIRE Autoimmune Regulator
APECED Autoimmune Polyendocrinopathy Candidiasis Ectodermal Dystrophy ARN Acide ribonucléique
CDR Complementary determining regions CMH Complexe majeur d’histocompatibilité CPA Cellule présentatrice d’antigène CRAC Calcium released-activated calcium channels
cTEC Cortical thymic epithelial cell (cellule épithéliale corticale thymique) CTL Cytotoxic T lymphocyte
CTLA-4 Cytotoxic T lymphocyte associated protein-4
DAG (1,2)Diacylglycérol
DC Dendritic cell (cellule dendritique)
DN Double négative
dnTGF-βRII Dominant négatif pour le récepteur de type II au TGF-β
DP Double positive
EAE Encéphalomyélite Autoimmune Expérimentale Erk Extracellular signal-regulated protein kinases ERM Ezrine-Radixine-Moésine
Foxp3 Forkhead box P3
GFP Green fluorescent protein
GM-CSF Granulo-Monocytes-Colony Stimulating Factor GPI Glycophosphatidylinositol
GVHD Graft versus host disease (maladie du greffon contre l’hôte) IDO Indoleamine 2,3 dioxygenase
IFN Interferon
Ig Immunoglobuline IL Interleukine
IPEX Immunedysregulation Polyendocrinopathy Enteropathy X linked ITAM Immunoreceptor tyrosine-based activation motifs
KO Knockout
LAT Linker for Activation of T cells
LFA-1 Lymphocyte function-associated antigen-1 LPS Lipopolysaccharide
MAPK Mitogen-Activated Protein Kinase MOG Myelin oligodendrocyte glycoprotein mTEC Medullar thymic epithelial cell MTOC Microtubule organizing center NFAT Nuclear Factor of activated T cells NFKB Nuclear Factor-kappa B
NK Natural Killer
NO Nitric oxyde
NOD Non obese diabetic
Pi3K Phosphatidylinositol-3-OH kinase RAG Recombination activating gene
SCID Severe combined immunodeficiency
SI Synapse immunologique
SMAC Supra molecular activation cluster TCR T cell receptor
TGF Transforming growth factor
TH T Helper
TIL Tumor infiltrating lymphocyte TLR Toll like receptor
TNF Tumor necrosis factor
Treg Lymphocyte T CD4+ CD25+ régulateur WASP Wiskott-Aldrich Syndrom protein
Le terme immunité (du latin immunis : protégé de…) évoque au premier abord les mécanismes de défenses que notre organisme développe à l’encontre des micro-organismes. La notion de système quant à elle désigne un ensemble d’éléments qui vont interagir de façon hiérarchisée et intégrée afin d’assurer une fonction. Le système immunitaire a pour objectif, dans une situation physiologique, de permettre à un organisme pluricellulaire de maintenir son intégrité, c'est-à-dire de préserver sa cohérence tissulaire et cellulaire en éliminant ses propres constituants altérés.
Dans un contexte physiopathologique, l’objectif à atteindre est identique. Pour cela un ensemble de mécanismes cellulaires et moléculaires va se mettre en place afin de préserver l’intégrité de l’organisme. Ainsi, une infection virale ou bactérienne, l’apparition et le développement d’une tumeur ou la greffe allogénique d’un organe ou d’un tissu peuvent conduire à l’activation du système immunitaire.
Un des éléments clé pour le développement de la réponse immunitaire va être la capacité à distinguer entre, d’une part, les constituants normaux de l’organisme (le soi) qui doivent être préservés et d’autre part, les agents pathogènes (le non soi) et les constituants altérés de l’organisme (le soi modifié). Afin d’assurer cette fonction, deux stratégies différentes ont été développées au cours de l’évolution des êtres vivants. La première ligne de défense va reposer sur le système immunitaire inné. C’est une réaction immédiate mettant en jeu différents acteurs moléculaires, cellulaires et tissulaires agissant au niveau local et systémique. Cette réponse rapide se fait indépendamment de la reconnaissance d’antigènes spécifiques et sans établissement d’une mémoire. Elle est basée sur la reconnaissance de motifs moléculaires conservés à la surface des pathogènes, les PAMP
(Pathogen-Associated Molecular Pattern) par les PRR (Pattern Recognition Receptor) exprimés à la
surface des cellules phagocytaires issues des lignages myéloïdes tels que les monocytes/macrophages ou les cellules dendritiques. Les cellules NK (Natural Killer) douées naturellement d’une activité cytotoxique vont, par la reconnaissance de signaux de stress cellulaire et de danger, participer activement à la réponse innée. Bien que dans certains cas, cette réponse immunitaire soit suffisante pour l’élimination des agents pathogènes, l’absence de spécificité antigénique va être le frein de ce type de réponse. C’est pourquoi, il peut être nécessaire d’activer par le biais de cellules de l’immunité innée des cellules effectrices appartenant à l’immunité adaptative. La réponse adaptative, seconde ligne de défense est apparue avec les vertébrés il y a environ 500 millions d’années. Elle repose sur des cellules d’origines hématopoïétiques, les lymphocytes T et B, portant à leur surface des récepteurs à l’antigène hautement spécifiques et ayant la capacité de développer en cas de réinfection par le même pathogène, une mémoire immunitaire à long terme. C’est une réponse plus lente, nécessitant un processus d’expansion clonale des lymphocytes spécifiques des antigènes du pathogène. Les lymphocytes B reconnaissent des antigènes entiers ou natifs
grâce à leur récepteur, le BCR (B Cell Receptor) qui est une immunoglobuline de surface. Les lymphocytes T quant à eux reconnaissent les antigènes sous forme de fragments peptidiques liés à des molécules du CMH (Complexe Majeur d’Histocompatibilité) présentes à la surface des CPA (Cellules Présentatrices de l’Antigène), cellules de l’immunité innée. Afin de faire face au très large spectre d’agents pathogènes, au cours du développement lymphocytaire dans les organes lymphoïdes primaires, les récepteurs aux antigènes sont générés aléatoirement par recombinaison génique. Cependant, ce processus aléatoire conduit à l’émergence de clones lymphocytaires potentiellement autoréactifs et donc dangereux pour l’organisme. Pour palier à cela, des mécanismes de tolérance centrale au soi sont mis en place afin d’éliminer ces lymphocytes exprimant des récepteurs autospécifiques. Ces mécanismes d’inactivation vont s’avérer cependant imparfaits et des lymphocytes T autoréactifs vont, après avoir échappé à cette sélection négative, pouvoir migrer vers les organes lymphoïdes périphériques et potentiellement déclencher des pathologies auto-immunes. Afin de prévenir cela, il existe des mécanismes de tolérance périphérique passifs, mais également actifs dépendant de l’action de lymphocytes T régulateurs. La population la mieux décrite à ce jour est celle mise en évidence par Sakaguchi en 1995 : la population régulatrice T CD4+ CD25+ Foxp3+. En plus de son rôle physiologique dans le contrôle du répertoire auto-immun, cette population est impliquée dans la modulation des réponses anti-infectieuses, dans la tolérance foeto-maternelle et a un rôle délétère dans l’immunosurveillance des tumeurs.
Mon travail de thèse a consisté à essayer de mieux comprendre les mécanismes d’action employés par ces lymphocytes T CD4+ CD25+ régulateurs pour contrôler la réponse immunitaire lors : de l’interaction entre un lymphocyte T CD4+ (LT CD4+) conventionnel et une cellule présentatrice de l’antigène (CPA), de l’activation de ce lymphocyte T mais également au cours de la mise en place de ses fonctions effectrices.
Les lymphocytes T
En plus des fonctions biologiques effectrices intrinsèques mises en œuvre afin d’éliminer les agents pathogènes, les lymphocytes T αβ s’avèrent cruciaux lors des réponses immunitaires adaptatives en « orchestrant » et en coordonnant ces réponses. En effet, les lymphocytes T CD4+ auxiliaires ou T
H (T helpers) permettent le développement de la majorité des réponses immunitaires humorales et, par la production de cytokines modulent l’activité des phagocytes. Les lymphocytes T αβ sont fortement majoritaires dans l’organisme, il existe toutefois une population de lymphocytes T γδ présente dans le sang, la peau et les muqueuses pouvant représenter de 1 à 10% des lymphocytes T totaux. Au cours de cette étude nous nous sommes intéressés uniquement aux lymphocytes T αβ.
« Ce que l’on tient pour compliqué peut-être expliqué en termes simples, et compris par tous ceux qui ont su garder leur capacité d’émerveillement. »
Jean-Pierre Revillard
I. Développement et Sélection des Lymphocytes T
αβ
L'ontogenèse des lymphocytes T αβ est un long processus qui démarre comme pour toutes les cellules d’origine hématopoïétique au sein de la moelle osseuse. La cellule initiale est la cellule souche hématopoïétique (CSH), cellule multipotente proliférant au contact du stroma médullaire. Cette cellule à l’origine de toutes les cellules sanguines, va progressivement se différencier dans la moelle osseuse en perdant peu à peu son potentiel au cours d’un processus appelé, hématopoïèse. Rapidement au cours de ce processus de différenciation, la cellule peut donner soit un progéniteur myéloïde commun (PMC), soit un progéniteur lymphoïde commun (PLC). Le PLC, cellule multipotente peut générer trois lignages lymphoïdes (B, T et NK) suivant les signaux qu’elle intégre. Le choix d’un lignage T conduit ce PLC à migrer en périphérie et à pénétrer au niveau de la jonction cortico-médullaire thymique pour y poursuivre son processus de différenciation (Figure 1.). Durant ces dernières années il a été suggéré que cet engagement vers le lignage T est un évènement intrathymique qui se réalise après engagement du récepteur Notch1 avec son ligand thymique Delta-1 L (von Boehmer et al. 2003). En effet, des souris déficientes pour Notch1, générées par un « knockout » conditionnel dans les précurseurs lymphoïdes précoces, ne possèdent pas de cellules pro-T mais disposent d’une quantité accrue de cellules pro-B et de cellules B matures. En absence de Notch-1 le choix du lignage se fait « par défaut » et des cellules B sont retrouvées dans le thymus (Wilson et al. 2001). Cette phase de différenciation
en cellule pro-T permet de constituer un stock de thymocytes immatures. Ces thymocytes en développement vont progressivement migrer du cortex à la médulla en subissant une sélection drastique, en effet seulement 2% d’entre eux sortiront du thymus pour constituer le pool périphérique de lymphocytes T matures.
Figure 1. Au cours de l’hématopoïèse, les cellules souches hématopoïétiques (CSH) se différencient en progéniteur lymphoïde commun 2 (PLC 2), cellule pivot dans le choix des lignages lymphoïdes. Une partie du pool de PLC 2 migre au niveau thymique et s’engage dans le développement T après interaction entre le récepteur Notch1 à la surface des précurseurs et son ligand Delta-1 L à la surface du stroma thymique.
Cette sélection s’opère via deux processus, la sélection positive tout d’abord, puis la sélection négative. Le développement lymphocytaire T va être instrumenté par l’expression successive et ordonnée de différents marqueurs de surface dont les corécepteurs CD4 et CD8 (Cluster of Differentiation). Sur la base de l’expression de ces corécepteurs il est d’ailleurs possible de fractionner la maturation des thymocytes en trois étapes principales (Figure 2). Tout d’abord, les thymocytes sont à un stade CD4- CD8- appelé double négatif (Beals et al.), population représentant entre 1 et 5% des thymocytes totaux. Sur la base de l’expression des marqueurs CD25 et CD44, quatre stades différents de thymocytes peuvent être observés, de DN1 à DN4 (Godfrey et al. 1993). Durant les stades DN2 et DN3, ils expriment la machinerie moléculaire nécessaire au réarrangement des gènes V, D et J codant la chaîne β du TCR. Il s’agit notamment des enzymes de recombinaisons RAG1 (Mombaerts et al. 1992) et RAG2 (Shinkai et al. 1992; Shinkai et al. 1993). Si ce processus de recombinaison génique est productif, la chaîne β fonctionnelle peut s’apparier alors avec un substitut de chaîne α, la pré-chaîne α (pTα) pour former à la surface des thymocytes le complexe pré-TCR (Saint-Ruf et al. 1994; von Boehmer 2005). Afin de pouvoir signaler activement, le pré-TCR ne peut pas être exprimé seul à la membrane, il faut qu’il soit associé à d’autres molécules cruciales pour les évènements de signalisation proximale, les
molécules faisant partie du complexe CD3 (von Boehmer and Fehling 1997). L’initiation de la transduction du signal par ce complexe permet de lever le blocage au stade DN3 et réprimer l’expression des gènes codant les enzymes RAG1 et RAG2 conduisant ainsi à l’exclusion allélique du locus β non réarrangé. Le stade DN4 est associé à la perte du marqueur CD25 puis la co-expression des marqueurs CD4 et CD8 effectué en parallèle d’une prolifération cellulaire intense. Après 6 à 8 cycles de divisions, la répression du pré-TCR marque le début de la deuxième étape de maturation des thymocytes, le stade double positif (Alam et al.), soit 80 à 90% des thymocytes totaux (Sebzda et al. 1999). A partir de là, vont débuter les recombinaisons géniques au niveau du locus de la chaîne α du TCR, concernant les segments V et J. Un réarrangement fonctionnel va permettre l’appariement des chaînes α et β du TCR et donc son expression à la surface des cellules DP.
Figure 2. Schéma résumant, d’après des données obtenues à partir de modèles murins, le processus de maturation intra-thymique des précurseurs T ayant pour objectif la génération de lymphocytes T CD4+ et T CD8+ matures. Durant leur développement aux stades double négatif et double positif, les thymocytes vont subir un remodelage génique afin d’exprimer un TCR αβ fonctionnel. Le passage au stade simple positif sera marqué par une sélection positive puis négative des cellules, basée sur les caractéristiques de leur TCR (restriction, spécificité et affinité).
L’expression du TCR αβ mature à la surface des thymocytes marque le début de la première sélection des thymocytes : la sélection positive. Ce TCR généré aléatoirement, pour être sélectionné positivement doit être capable de reconnaître des peptides antigéniques présentés dans un contexte de CMH du soi.
Le paramètre affinité du TCR vis-à-vis des complexes CMH du soi / peptide antigénique aboutira à une élimination de près de 90% des thymocytes DP incapables de les reconnaître. Cette majorité de cellules mourra donc par « négligence » étant dans l’incapacité d’intégrer les signaux de survie nécessaires par leur TCR. La sélection positive décrite dans les années 70 (Zinkernagel and Doherty 1974; Bevan 1977; Fink and Bevan
1978; Zinkernagel et al. 1978) a lieu dans la partie corticale du thymus par les cTEC présentant des antigènes du soi (cellules épithéliales corticales thymiques) (Benoist and Mathis 1989). Plus récemment, une analyse en temps réel par microscopie bi-photonique a permis de visualiser les contacts entre les thymocytes en développement subissant la sélection positive et le stroma thymique (Bousso et al. 2002).
Suite à cela, les thymocytes migrent vers la partie basale du thymus au sein de la médulla où ils subissent la sélection négative. L’objectif étant cette fois d’éliminer les cellules ayant une trop forte affinité pour les complexes CMH/peptide antigénique du soi. En effet, ces thymocytes ayant un TCR potentiellement autoréactif pourraient, une fois sortis du thymus représenter un danger pour l’organisme.
Des travaux utilisant des souris RelB-/- ont montré que l’absence de sélection négative due à un thymus désorganisé et une atrophie médullaire a pour conséquence l’émergence d’auto-immunité périphérique multi-organes (Weih et al. 1995). D’où l’importance capitale de cette induction de tolérance centrale des thymocytes, mise en lumière il y a déjà plus de vingt ans par Polly Matzinger (Matzinger et al. 1984). Depuis, deux mécanismes de sélection négative ont été décrits. Si un thymocyte autoréactif reconnaît un peptide du soi présenté par une CPA médullaire (en majorité des cellules dendritiques (DC)), il subira alors une délétion clonale et sera donc éliminé par apoptose (Page et al. 1996) (figure 3). Une étude quantitative et cinétique des thymocytes sélectionnés positivement montre qu’entre la moitié et les deux tiers d’entre eux sont ainsi éliminés (van Meerwijk et al. 1997).
En revanche, la reconnaissance par un thymocyte d’un peptide du soi à la surface d’une mTec (cellule médullaire épithéliale thymique) aboutira à son inactivation fonctionnelle : l’anergie. En effet, les mTec expriment de manière ectopique au niveau du thymus des antigènes caractéristiques d’oganes périphériques (foie, rein). Cette expression est sous le contrôle de la protéine AIRE (Autoimmune Regulator) régulant l’expression d’une importante quantité de gènes (Anderson et al. 2002). La déficience de AIRE aussi bien chez des souris AIRE-/- que chez des patients atteints d’APECED (Autoimmune PolyEndocrinopathy
Candidiasis Ectodermal Dystrophy), pathologie autosomique récessive monogénique,
conduit à des symptômes auto-immuns similaires (Villasenor et al. 2005).
Malgré l’apparente perfection de cette machinerie thymique, des lymphocytes T potentiellement auto-réactifs et donc dangereux pour l’organisme échappent à cette double sélection et émergent dans la circulation sanguine ainsi que dans les tissus lymphoïdes périphériques. Certains antigènes tissulaires ne seraient pas exprimés via AIRE ou a des niveaux insuffisants pour induire une sélection négative (Klein et al. 2000). Ce défaut thymique peut-être considéré comme une conséquence évolutionniste permettant de maximiser la taille du répertoire T en périphérie, conséquence à double tranchant pour l’organisme, bénéfique dans la lutte contre les pathogènes mais délétère pour la
préservation de son intégrité. Heureusement, des mécanismes de tolérance périphérique au soi existent et ont été décrits (Walker and Abbas 2002; Redmond and Sherman 2005). Il y a tout d’abord des mécanismes de types « passifs » : ignorance, induction d’apoptose ou d’anergie. L’induction de tolérance périphérique qualifiée « d’active » est réalisée par les lymphocytes T CD4+ CD25+ régulateurs (Figure 3.).
Figure 3. Schématisation des processus de sélection thymique. Après avoir réarrangé les chaînes α et β du TCR les thymocytes subissent une première sélection, la sélection positive. La restriction au CMH du soi conditionne la survie des thymocytes. La sélection négative permet ensuite d’éliminer les thymocytes ayant une trop forte autospécificité, la tolérance centrale s’opère par induction d’apoptose (délétion clonale) ou bien par inactivation fonctionnelle (anergie). Une fois la périphérie atteinte, une partie des LT potentiellement autoréactifs est contrôlée par les LT CD4+ CD25+ régulateurs, mécanisme « actif » et majeur de la tolérance périphérique. La tolérance périphérique pourra également s’opérer par des mécanismes qualifiés de passifs. En effet, la séquestration de certains antigènes tissulaires au sein de domaines immunoprivilégiés peut empêcher l’activation de ces clones T autospécifiques.
Les thymocytes ayant survécu à cette sélection impitoyable sortent sous la forme de lymphocytes T simple positif (SP) dits naïfs (car n’ayant jamais été en contact avec l’antigène dont ils sont spécifiques). Les thymocytes possédant un TCR restreint par le CMH de classe II se développent en lymphocytes T CD4+ (Berg et al. 1989), ceux possédant un TCR restreint par le CMH de classe I se développent en lymphocytes T CD8+ (Teh et al. 1988). Une fois sortie du thymus, ces lymphocytes T vont, par voie systémique, circuler continuellement entre les différents organes lymphoïdes secondaires dans l’attente de
rencontrer leur antigène. C’est d’ailleurs dans ces organes que, par exemple dans le cadre d’une viro-infection, une cellule dendritique présentant un peptide viral va activer le lymphocyte T naïf et spécifique de ce peptide antigénique. L’activation de ce lymphocyte T lui permettra de proliférer, de se différencier et d’acquérir ses fonctions effectrices.
II. Récepteur à l’antigène des lymphocytes T αβ
La rencontre entre un lymphocyte T et une CPA et donc, l’interaction privilégiée entre le TCR et le complexe CMH/peptide spécifique va permettre l’activation et l’expansion clonale du lymphocyte T (Davis et al. 1998). La cellule T via son TCR (hétérodimère glycosylé polymorphe) accumule des informations quantitatives et qualitatives sur cet antigène, l’objectif étant ensuite de relayer ces informations à l’intérieur de la cellule. Cependant, l’absence de domaines intracellulaires pour le TCR va nécessiter l’association et le transfert d’informations de ce récepteur aux molécules CD3, protéines non polymorphes dotées d’importants domaines intracellulaires de signalisation. Le complexe TCR/CD3 ainsi formé va pouvoir induire les voies de transduction du signal proximales et précoces permettant ainsi l’activation de la cellule T (Clevers et al. 1988).
Structure du TCR :
Le TCR est composé de deux chaînes transmembranaires polypeptidiques glycosylées α et β. Ces glycoprotéines de type I appartiennent à la superfamille des immunoglobulines (Ig). Chaque chaîne est composée d’un domaine extracellulaire amino-terminal contenant une région variable (V), une région constante (C) ainsi qu’une région charnière (Figure 4.). De proximal en distal font suite, un domaine transmembranaire hydrophobe composé de 5 à 12 acides aminés chargés positivement (favorisant la stabilité du complexe TCR/CD3) ainsi que d’un court domaine cytoplasmique. L’association des deux chaînes α et β se fait par l’intermédiaire d’un pont di-sulfure (Garcia et al. 1996; Schumacher 2002).
L’ensemble des régions variables (Vα et Vβ) ainsi réunies forme le site de reconnaissance à l’antigène. Chaque région variable possède trois régions hypervariables appelées CDR (Complementary Determining Region) (Chothia et al. 1988) par homologie aux CDR identifiés sur les régions variables des immunoglobulines. Les régions CDR1 et CDR2 permettent la liaison aux molécules de CMH, le CDR3 se liant préférentiellement au peptide antigénique (Jorgensen et al. 1992; Sant'Angelo et al. 1996).
Le complexe TCR/CD3 :
Le complexe CD3 peut-être qualifié de plateforme de transduction du signal associé à l’hétérodimère αβ. Il est composé des sous-unités polypeptidiques non polymorphes δ, ε, γ et
ζ qui vont s’associer de manière non covalente, afin de former les hétérodimères CD3δε, CD3γε ainsi que l’homodimère CD3ζζ (Schumacher 2002; Rudolph et al. 2006). A l’exception du domaine CD3ζ, toutes les sous-unités CD3 possèdent une portion transmembranaire chargée négativement qui va permettre l’association avec le TCR et surtout favoriser la stabilité du complexe CD3/TCR (Koning et al. 1990; Manolios et al. 1991).
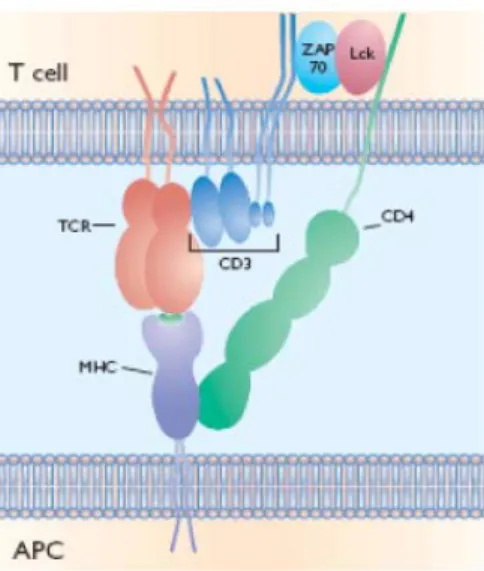
En réponse à l’engagement du TCR par le complexe CMH/peptide, les portions cytoplasmiques des molécules CD3 permettent l’initiation de la signalisation grâce à leurs motifs ITAM (Immunoreceptor tyrosines-based activation motifs : YxxL(X)6-8YxxL) (Figure 4).
Figure 4. Organisation spatiale du complexe TCR/CD3. Le TCR constitué de deux chaînes α (gris) et β (violet) a pour fonction principale la reconnaissance du complexe CMH/peptide antigénique spécifique. Cette reconnaissance permet de recueillir des informations qualitatives et quantitatives qui seront traduites en cascade par des voies de signalisation intracellulaire induites par les molécules CD3. En effet, seules les molécules CD3δε, CD3γε et CD3ζζ vont pouvoir transduire ces signaux grâce à leurs larges domaines cytoplasmiques portant des motifs ITAMs.
Les molécules CD3γ, CD3δ et CD3ε possèdent chacun un motif ITAM, chaque molécule CD3ζ possédant trois motifs ITAM. La phosphorylation des tyrosines de ces motifs ITAM va permettre le recrutement de protéines de signalisation importantes (Samelson et al. 1985). L’assemblage du complexe s’effectue dans le réticulum endoplasmique par des mécanismes rigoureusement contrôlés. La chaîne α du TCR s’associe dans un premier temps à
l’hétérodimère CD3δε, permettant ainsi l’interaction entre l’hétérodimère CD3γε et la chaîne β du TCR. Les deux trimères ainsi formés s’associent pour former l’hexamère suivant TCRαβ/CD3γε/CD3δε. Cet hexamère va enfin s’associer avec l’homodimère CD3ζζ pour former un complexe TCR/CD3 fonctionnel qui pourra être exprimé à la surface du lymphocyte T (Alarcon et al. 1988; Huppa and Ploegh 1997).
La stoechiométrie du complexe TCR/CD3 a longtemps été source de débats, mais il semble aujourd’hui établi qu’un seul hétérodimère TCR αβ s’associe avec un seul hexamère CD3. Les étapes d’assemblages résultant de la formation d’une interface à trois hélices transmembranaires impliquent un résidu basique du TCR et une paire de résidus acides des dimères CD3 (Call et al. 2002; Call and Wucherpfennig 2005; Kuhns et al. 2006).
III. Organisation des gènes du TCR
Afin de se prémunir au mieux face à l’immense diversité des agents pathogènes, l’organisme a eu pour unique solution de générer aléatoirement la spécificité des TCR pour les peptides antigéniques. De cette façon, la taille et la diversité du répertoire des cellules T naïves qui vont émerger du thymus vers la périphérie sont maximisées.
Le réarrangement des gènes codant pour les chaînes α et β est réalisé au cours du développement intrathymique des lymphocytes T. Les processus de recombinaison génique employés vont être très similaires à ceux utilisés pour les réarrangements des chaînes lourdes et légères des immunoglobulines au cours de la lymphopoïèse B. Chez l’homme, les loci α et β sont composés, dans leur configuration germinale de segments V (Variable), J (Joining) et C (Constant) pour la chaîne α et V, D (Diversity), J et C pour la chaîne β.
Figure 5. Réarrangements des segments de gène codant les chaînes α et β du TCR. La chaîne β est la première à être réarrangée, d’abord les segments D et J, puis le segment V sera réarrangé. Un réarrangement productif permettra d’initier le réarrangement de la chaîne α. Des mécanismes enzymatiques additionnels permettront d’accroître la variabilité du répertoire des TCR.
Les gènes de la chaîne β se réarrangent en premier. On observe tout d’abord l’association d’un segment D avec un segment J, puis un segment V est réarrangé avec le segment DJ précédemment obtenu (Figure 5.). Un réarrangement productif de la chaîne β permet son association avec la chaîne pTα ainsi que l’expression à la surface des thymocytes DN (cf.
Développement et Sélection des lymphocytes Tαβ). Suite à cela, le réarrangement des
gènes codant la chaîne α se déroulera de façon similaire. Par ces mécanismes, la multiplicité des segments de gènes permet d’établir une importante diversité combinatoire et jonctionnelle (Arden et al. 1995).
Avec toujours pour objectif d’augmenter cette diversité, des mécanismes enzymatiques additionnels vont intervenir. L’exonucléase (élimination aléatoire de nucléotides) ou la Terminal deoxynucleotidyl Transferase (TdT) (ajout aléatoire de nucléotides) favorisent l’obtention d’un répertoire de TCR encore plus vaste. Il est estimé à plus de 1013 le nombre de TCR pouvant être sélectionnés au cours du développement thymique. Des approches expérimentales pour évaluer cette diversité en périphérie ont donné les résultats suivants : 2x106 TCR chez la souris et 2x107 TCR chez l’homme (Nikolich-Zugich et al. 2004).
Activation des lymphocytes T
Le pré-requis pour l’activation T, comme introduit plus haut, est conditionné par l’interaction productive entre d’une part un lymphocyte T et d’autre part une cellule présentatrice d’antigène. Lors de ce contact cellulaire, il va y avoir reconnaissance par le TCR de la cellule T du complexe CMH/peptide porté par la CPA. Cette interaction moléculaire va permettre l’induction de réponses immunitaires adaptatives hautement spécifiques. L’étude des paramètres contrôlant positivement ou négativement cette interaction cellulaire est donc d’un grand intérêt. De nombreuses équipes de recherche travaillent sur cette problématique qui, malgré toutes les avancées obtenues lors des dernières décennies, suscite encore de nombreuses interrogations. Au premier abord cette interaction semble simple, cependant plusieurs obstacles importants existent.
En effet, la mesure de l’affinité entre le TCR et le complexe CMH/peptide a révélé une faible affinité de liaison avec une constante de dissociation comprise entre 10-4 et 10-7 M (Weber et
al. 1992; Alam et al. 1996). L’analyse par technologie BIAcore a permis de mesurer la
demi-vie d’association entre le TCR et son ligand, cette demi-demi-vie n’étant que de quelques secondes (Matsui et al. 1994; Davis et al. 1998). Ces études quantitatives mettent en lumière un paradoxe : comment avec des paramètres cinétiques si défavorables (faible affinité et demi-vie d’association rapide) l’interaction TCR/CMH/peptide peut-elle engendrer une réponse moléculaire de signalisation aussi importante et sophistiquée ?
De plus, un autre point négatif va être le faible nombre de complexes CMH/peptides spécifiques d’un TCR présent à la surface d’une CPA. La majorité des molécules de CMH à la surface de la CPA exprimeront des peptides du soi issus du catabolisme de protéines intracellulaires ou provenant du milieu extracellulaire. Toutefois, entre 60 et 200 complexes CMH/peptide antigénique spécifique à la surface d’une CPA suffisent à déclencher une réponse lymphocytaire T (environ 0,03% des complexes CMH/peptide exprimés à la membrane plasmique d’une CPA) (Demotz et al. 1990).
Le lymphocyte T va donc devoir scanner la surface de la CPA à la recherche de son ligand antigénique. La probabilité que les complexes CMH/peptique spécifique se trouvent dans la zone initiale de contact entre ces deux cellules est relativement faible. L’initiation de la transduction du signal en aval du TCR est donc due à une interaction moléculaire hautement sensible (une faible quantité de ligand devant engendrer une forte réponse) et spécifique (très peu de ligands spécifiques parmi un océan de ligands non spécifiques). Tous ces éléments réunis soulèvent donc une nouvelle interrogation sur les mécanismes développés par la cellule T afin d’assurer une détection efficace de faibles concentrations antigéniques.
Un paramètre compliquant également l’interaction TCR/CMH/peptide est la taille réduite du TCR par rapport aux autres molécules membranaires, le complexe TCR/CMH ne mesurant environ que 15 nm (Garcia et al. 1996). La taille réduite du TCR ne va pas être un atout pour interagir avec son ligand sachant que d’autres molécules membranaires comme la phosphatase CD45 ont une taille de 45 nm (Cyster et al. 1991). Cela suggère que des réarrangements moléculaires membranaires à l’aire de contact cellule T/CPA doivent s’opérer.
Un dernier point rendant encore plus complexe la mécanistique de cette interaction est la nécessité de la soutenance de la signalisation, pour l’activation et la mise en place des fonctions effectrices des lymphocytes T (Goldsmith and Weiss 1988; Valitutti et al. 1995; Iezzi et al. 1998).
Tous ces éléments sur la mécanistique d’activation des lymphocytes T ont soulevé de nombreuses questions. Afin d’essayer d’y répondre différents modèles ont été proposés.
I. Sensibilité et spécificité des TCR
Le modèle du « Kinetic Proofreading » (McKeithan 1995)
Le modèle de « kinetic proofreading » a été à l’origine proposé par Hopfield pour expliquer la redoutable efficacité lors des processus de réplication de l’ADN et de transcription/traduction conduisant à la synthèse protéique (Hopfield 1974). S’appuyant sur ce concept, c’est en 1995 que McKeithan transpose le modèle du « kinetic proofreading » aux voies de signalisation induites en aval du TCR (McKeithan 1995). Les bases de ce modèle reposent sur la notion de temps d’interaction entre le TCR et son ligand. Il explique tout d’abord dans son travail, que l’interaction entre le TCR et son ligand ne se fait pas parallèlement à l’activation de la cellule T mais nécessite un certain temps pour engendrer cette réponse. Ce délai se justifie biologiquement par la nécessité d’étapes successives de phosphorylations sur des tyrosines essentielles à la mise en place de voies de signalisation productives. Ceci permet de comprendre qu’une interaction entre un TCR et un complexe CMH-peptide non spécifique ayant un taux de dissociation très rapide ainsi qu’un temps d’interaction très court ne peut conduire qu’à l’obtention d’un signal abortif. Dans les mêmes concentrations antigéniques, l’interaction TCR-CMH/peptide spécifique plus stable pourra générer un signal d’activation.
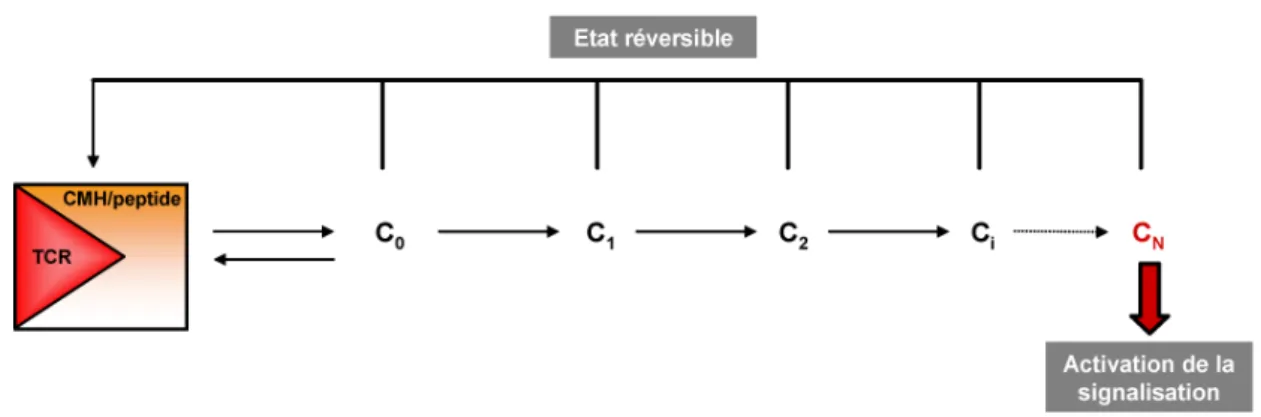
La schématisation du modèle est la suivante (Figure 6.) : l’interaction TCR/CMH/peptide spécifique ou non spécifique forme un complexe (C0). Ce complexe pouvant à tout moment être dissocié, est converti en une série d’intermédiaires (Ci) avec pour but d’obtenir un complexe actif (CN) qui permet d’induire la réponse biologique. Chaque étape est nécessaire
et consomme de l’énergie car elle implique des processus enzymatiques de phosphorylation sur tyrosines. Toutefois chacun des intermédiaires peut-être dissocié et donc chaque étape peut-être réversible. Dans le cas de la signalisation du TCR, des phosphatases permettront le processus inverse en induisant un retour à l’état basal pour chacun des intermédiaires réactionnels.
Pour des complexes TCR/CMH/peptide non spécifique, le taux de dissociation est suffisamment important pour empêcher la formation du complexe actif (CN).
Figure 6. Schématisation du modèle de « kinetic proofreading » adapté à la transduction du signal en aval du TCR. Le complexe C0 formé entre le TCR d’une part et le complexe CMH/peptide d’autre part va subir N
modifications afin de produire le complexe actif CN seul capable d’induire la signalisation. Chaque étape
est réversible, la dissociation du complexe entraînant un retour des intermédiaires à leur état basal.
Ce modèle proposé paraît donc intéressant car il permettrait théoriquement d’expliquer la haute sélectivité du TCR. En effet, la transduction du signal ne peut être mise en place que si l’interaction TCR/CMH-peptide est suffisamment spécifique pour durer assez longtemps afin que le complexe actif CN se forme. Depuis, de nombreux travaux utilisant des variants peptidiques induisant soit un signal inhibiteur (peptide antagoniste) soit un signal partiel (peptide antagoniste partiel) ont aidé à mieux comprendre les mécanismes d’engagement du TCR. Dans le cas des variants « peptidiques altérés » ou APL (Altered Peptide Ligand) qui ont une affinité de liaison plus faible, il y aura stimulation du TCR mais absence de signalisation soutenue permettant l’obtention du complexe actif. Un peptide antagoniste induira une rapide dissociation du complexe TCR/CMH/peptide, l’interaction sera donc non productive (Lyons et al. 1996). Ainsi, avec ce modèle prenant en compte surtout les aspects sélectifs et affins de l’interaction, une faible concentration de complexes CMH/peptide de haute affinité sera plus efficace qu’une forte concentration de CMH/peptide de basse affinité. Ce modèle met donc en avant l’affinité d’interaction comme paramètre crucial conditionnant le temps d’interaction entre le TCR et son ligand pour activer la signalisation cellulaire.
Cependant, ce modèle ne permet pas de déterminer la durée d’interaction suffisante pour induire une réponse biologique.
En 1996, Rabinowitz a apporté une nouvelle pierre au modèle du « kinetic proofreading » en proposant le modèle de « kinetic discrimination » (Rabinowitz et al. 1996). Il propose que l’utilisation d’un peptide antagoniste en plus de ne pas aboutir au travers de la cascade d’intermédiaires au complexe actif CN va avoir un effet inhibiteur sur l’activation de la cellule T. Une des zones d’ombre de ce modèle de « kinetic proofreading » réside dans l’identification moléculaire des intermédiaires réactionnels. Récemment une équipe a montré suite à la stimulation d’un lymphocyte T par un peptide agoniste, un changement conformationnel (détecté par l’utilisation de l’anticorps APA1/1) au niveau de la chaîne CD3ε non observable après stimulation par un peptide antagoniste (Risueno et al. 2005). Pourrait-on identifier ce changement de cPourrait-onformatiPourrait-on lié à la pleine activatiPourrait-on du lymphocyte T comme un des intermédiaires réactionnels du « kinetic proofreading » ?
Ce modèle pourrait prendre toute sa dimension biologique au cours de la sélection thymique où l’affinité de l’interaction TCR/CMH/peptide conditionne le devenir des thymocytes en développement (Alam et al. 1996).
Le modèle du « Serial Triggering » (Valitutti, 1995)
La sensibilité de détection de l’antigène est permise pour les lymphocytes B par une haute affinité de liaison entre leur récepteur (BCR : B Cell Receptor) et l’antigène spécifique. Le paradoxe des lymphocytes T est d’assurer une haute sensibilité de détection avec une faible affinité d’interaction. En effet, des lymphocytes TH (T helpers) ou T auxiliaires sont capables de s’activer, de proliférer ainsi que de produire des cytokines en réponse à une CPA présentant entre 50 et 100 complexes CMH/peptide spécifique (Demotz et al. 1990). Des études plus récentes ont montré qu’un seul complexe CMH/peptide est capable d’induire une signalisation calcique dans un lymphocyte T CD4+ (Irvine et al. 2002) et que trois complexes CMH/peptide sont capables d’engendrer la réponse cytotoxique des CTL (Cytotoxic T
Lymphocyte) (Purbhoo et al. 2004). Ces résultats suggèrent que l’activation T va se dérouler
au niveau d’une zone de contact privilégiée où peu de complexes CMH/peptide spécifique au milieu d’une masse de complexes CMH/peptide non spécifique pourraient permettre la pleine activation du lymphocyte T. Le rôle de ce site de contact entre le lymphocyte T et la CPA dans l’induction de la signalisation a été documenté par plusieurs travaux morphologiques (Donnadieu et al. 1994; Valitutti et al. 1995; Wulfing et al. 1997). Les résultats obtenus ont permis de comprendre que les lymphocytes T n’étaient pas passifs dans la reconnaissance du ligand antigénique mais scannaient activement la CPA à la recherche du complexe CMH/peptide spécifique grâce notamment au cytosquelette d’actine
(Valitutti et al. 1995). Il a également été montré que l’internalisation et le recyclage du TCR faisaient suite à l’interaction du TCR avec son ligand spécifique (Padovan et al. 1993).
S’appuyant sur ces éléments, en 1995 Valitutti et al. ont proposé un nouveau concept d’activation des lymphocytes T : le « serial triggering » ou engagement en série des TCR. Ce concept a permis d’expliquer le mécanisme sous-jacent au fait que seulement un petit nombre de complexes CMH/peptide spécifique liant avec une faible affinité le TCR, peuvent pleinement activer un lymphocyte T (Valitutti and Lanzavecchia 1997). En effet, dans ce travail les auteurs ont pu observer que lors d’un contact lymphocyte T/CPA aboutissant à la mise en place d’une signalisation soutenue, le nombre de TCR engagés était supérieur au nombre de complexes CMH/peptide spécifique. Ce modèle se base donc sur le fait que grâce à un taux de dissociation entre le TCR et son ligand assez important, un complexe CMH/peptide après dissociation d’un premier TCR peut engager séquentiellement un deuxième TCR (Figure 7.). Ce modèle exige une dynamique moléculaire importante au niveau du site de contact entre la cellule T et la CPA, notamment par l’apport régulier de nouveaux TCR à engager afin de maintenir la signalisation soutenue. Le temps d’interaction TCR/CMH/peptide doit être assez long pour induire un signal tout en étant assez court pour permettre la dissociation et l’engagement rapide d’un nouveau TCR afin de soutenir la signalisation. Le résultat de cet engagement en série des TCR peut-être quantifié expérimentalement en mesurant l’internalisation des TCR qui va varier proportionnellement à la concentration en complexes CMH/peptide spécifique présentés par la CPA (Valitutti and Lanzavecchia 1997; Coombs et al. 2002).
Figure 7. Modèle d’engagement en série des TCR ou « serial triggering ». Un seul complexe CMH/peptide spécifique peut engager séquentiellement plusieurs TCR durant l’interaction prolongée entre le lymphocyte T et la CPA. L’engagement d’un TCR va permettre le recrutement de protéines de signalisation et la mise en place des voies de transduction du signal. Puis le TCR engagé se libère de l’interaction avec le complexe CMH/peptide qui est alors libre de pouvoir engager un nouveau TCR. Les TCR engagés (en bleu) sont finalement internalisés.
Un peptide agoniste sera donc optimal dans ce modèle pour l’activation d’un lymphocyte T au contraire d’un peptide antagoniste qui occupera un TCR sans permettre l’induction de la signalisation. Le « serial triggering » permet donc de mieux appréhender les effets inhibiteurs des peptides antagonistes, car lorsque ces derniers ont engagé des TCR, ces TCR ne sont plus disponibles pour lier des peptides agonistes. D’autres études, par l’utilisation de variants peptidiques dont le taux de dissociation est diminué ou augmenté par rapport au peptide agoniste de référence, ont mieux documenté les conséquences de la spécificité de l’antigène sur l’activation T (Kalergis et al. 2001). En 1998, Hudrisier et al. ont montré l’importance de la fréquence de l’engagement en série des TCR pour la réponse fonctionnelle des CTL. En effet, la liaison covalente donc permanente entre le TCR et le complexe CMH/peptide abolit la signalisation soutenue dans les lymphocytes T (Hudrisier et al. 1998).
Le « serial triggering » est donc un modèle intéressant permettant de comprendre comment, avec si peu de ligands antigéniques spécifiques à leur surface et une faible affinité de liaison de ces ligands avec le TCR les interactions lymphocyte T/CPA peuvent être aussi productives. De plus dans ce modèle, la cinétique d’engagement des TCR doit être optimale pour pouvoir induire la signalisation soutenue, exigeant donc un peptide agoniste hautement spécifique. Cet autre aspect du modèle permet donc de comprendre la haute spécificité des réponses T in vivo résultat d’une sélection drastique pour la CPA offrant la stimulation optimale.
Il serait tentant ainsi de spéculer sur l’existence de deux stratégies différentes pour la reconnaissance antigénique entre BCR et TCR. Le BCR serait plutôt sélectionné sur le paramètre affinité, l’interaction la plus affine étant la plus productive, alors que le TCR serait plus finement sélectionné pour une cinétique d’engagement de son ligand optimale.
II. Activation de la Cellule T
Activation de la Cellule T et Valence des TCR
La nécessité de la multimérisation des TCR pour permettre l’activation des lymphocytes T est un dogme qui a récemment été remis en question. Cette théorie a été établie suite au constat que l’utilisation d’anticorps anti-CD3, afin d’activer une cellule T, était très efficace du fait de leur potentiel à agréger ou « cross-linker » les TCR (Kappler et al. 1983). Plus tard des travaux ont supporté ce prérequis de multimérisation en montrant que l’induction des réponses calciques dans les lymphocytes T nécessite l’utilisation de multimères solubles de complexes CMH/peptide (Boniface et al. 1998).
Plusieurs évidences remettent toutefois en discussion cette notion de multimérisation des TCR pour l’activation des lymphocytes T. En effet, les complexes CMH/peptides spécifiques