Pour l'obtention du grade de
DOCTEUR DE L'UNIVERSITÉ DE POITIERS UFR des sciences fondamentales et appliquées
Institut de paléoprimatologie, paléontologie humaine : évolution et paléoenvironnements - IPHEP (Poitiers)
(Diplôme National - Arrêté du 7 août 2006)
École doctorale : Sciences pour l'environnement - Gay Lussac (La Rochelle) Secteur de recherche : Paléontologie des vertébrés
Cotutelle : Université de Chulalongkorn
Présentée par :
Kantapon Suraprasit
Paléoenvironnements et reconstitutions paléoclimatiques du
Pléistocène moyen de Thaïlande et leur impact sur la
biodiversité et la distribution des espèces : la contribution de
la faune de vertébrés du gisement de Khok Sung (Province du
Khorat)
Directeur(s) de Thèse :Stéphane Ducrocq, Somsak Panha, Jean-Jacques Jaeger Soutenue le 16 novembre 2015 devant le jury
Jury :
Président Patrick Vignaud Professeur, IPHEP, Université de Poitiers Rapporteur Herve Bocherens Professeur,Université de Tübingen, Allemagne
Rapporteur Colin Groves Professor, Australian National University, Canberra, Australia Membre Stéphane Ducrocq Directeur de recherche CNRS, IPHEP, Poitiers
Membre Somsak Panha Professor, ASRU, Bangkok
Membre Jean-Jacques Jaeger Professeur émérite, IPHEP, Université de Poitiers
Membre Yaowalak Chaimanee Ingénieur de recherche, CNRS, IPHEP, Université de Poitiers
gisement de Khok Sung (Province du Khorat) [En ligne]. Thèse Paléontologie des vertébrés. Poitiers : Université de Poitiers, 2015. Disponible sur Internet <http://theses.univ-poitiers.fr>
DOCTEUR DE L’UNIVERSITÉ DE POITIERS Faculté des Sciences Fondamentales et Appliquées
(Diplôme National – Arrêté du 7 août 2006)
Ecole Doctorale Gay Lussac Sciences pour l’environnement Secteur de recherche : Terre solide et Enveloppes superficielles
Discipline : Paléontologie des vertébrés Par
Kantapon SURAPRASIT
Paléoenvironnements et reconstitutions paléoclimatiques du
Pléistocène moyen de Thaïlande et leur impact sur la biodiversité
et la distribution des espèces: la contribution de la faune de
vertébrés du gisement de Khok Sung (Province du Khorat)
Sous la direction de Stéphane DUCROCQ Jean-Jacques JAEGER Olivier CHAVASSEAU
Soutenue le 16 Novembre 2015 Devant la commission d’Examen
JURY
Colin P. GROVES Professeur, ANU, Canberra Rapporteur Hervé BOCHERENS Professeur, HEP, Tübingen Rapporteur Somsak PANHA Professeur, ASRU, Bangkok Examinateur Jean-Jacques JAEGER Professeur émérite, iPHEP, Poitiers Examinateur Patrick VIGNAUD Professeur, iPHEP, Poitiers Examinateur Stéphane DUCROCQ Directeur de recherche CNRS, iPHEP, Poitiers Examinateur Yaowalak CHAIMANEE Ingénieur de recherche CNRS, iPHEP, Poitiers Examinateur John de VOS Associé de recherche, NBC, Leiden Examinateur
Doctor of Philosophy
Faculty of Fundamental and Applied Sciences (National Diploma – Decree of August 7th 2006)
Ecole Doctorale Gay Lussac Sciences pour l’environnement Research sector : Terre solide et Enveloppes superficielles
Discipline : Vertebrate paleontology
Kantapon SURAPRASIT
Paleoenvironmental and paleoclimatic reconstructions of the Thai
Middle Pleistocene and their impacts on the biodiversity and the
species composition and distribution: the contribution of Khok
Sung (Khorat province, Thailand) vertebrate community
Supervisors Stéphane DUCROCQ Jean-Jacques JAEGER Olivier CHAVASSEAU
Defense date : 16th Novembre 2015
Committee in charge :
Colin P. GROVES Professor, ANU, Canberra External reviewer Hervé BOCHERENS Professor, HEP, Tübingen External reviewer Somsak PANHA Professor, ASRU, Bangkok Committee Jean-Jacques JAEGER Professor emeritus, iPHEP, Poitiers Committee Patrick VIGNAUD Professor, iPHEP, Poitiers Committee Stéphane DUCROCQ Research director (CNRS), iPHEP, Poitiers Committee Yaowalak CHAIMANEE Research engineer (CNRS), iPHEP, Poitiers Committee John de VOS Research associate, NBC, Leiden Committee
I wish to thank all of the supervisors and colleagues who have helped me to complete this research and to attain my Doctor of Philosophy degree in paleontology. I would like to express deepest gratitude to my thesis supervisor, Prof. Dr. Jean-Jacques Jaeger, who provided me all opportunities to complete my study in France, led me down research avenues, and supported me to maintain the necessary persistence to explore them. I would like to extend special thanks to my Thai co-supervisor, Prof. Dr. Somsak Panha (Chulalongkorn University), for his advices, encouragement, and financial supports throughout this research project. I would particularly like to thank Dr. Olivier Chavasseau for his constructive improvements and reviews of this work, as well as all previously related researches. I am grateful to Dr. Stéphane Ducrocq for his greatly supervisory responsibility and his helps in providing some references through this study. I am obligated to Prof. Dr. Patrick Vignaud who has helped me manage an official registration and all academic requirements since my first arrival in France, and has kindly offered me financial supports from the laboratory for research studies. I also appreciate the assistance of Dr. Mouloud Benammi and his insightful suggestions for the paleomagnetic data. I am indebted to Dr. Yaowalak Chaimanee, who gave me the first opportunity to study mammalian fossils in her care and became a part of my future as a result of coming to France, and for that it was the best decision of my life. She always encouraged and inspired me to step beyond the traditional field of geology into the fantastic world of vertebrate paleontology.
The completion of this research would not have been possible without the assistances of several museums, departments, and laboratories and their associated personnel. I would like to thank all of the curators, who permitted me access to the collection and provided me the comparative fossil and extant material, including Dr. Joséphine Lesur (MNHN), Chen Jin (IVPP), Natasja den Ouden (RMNH), Dr. Frank Zachos (NMW), Alexander Bibl (NMW), Gassner Georg (NMW), Dr. Reinhard Ziegler (SMNS), Michael Hiermeier (ZSM), and Cholawit Thongcheroenchaikit (THNHM). I would like to express our grateful thanks to Dr. John de Vos (NBC) who provided me some additional information on the Pleistocene mammalian taxonomy during my visit to the Dubois collection. I am grateful to Prof. Dr. Hervé Bocherens (Universität Tübingen) for allowing me to carry out an isotope analysis on the Mae Moh and Khok Sung faunas at his biogeochemistry laboratory and for his assistance and helpful discussions on the stable isotope results. In addition, I thank Christoph Wissing, Martin Cotte, and Bernd Steinhilber from Universität Tübingen for their valuable helps in
Valentin for his help in making fossil replicas, and to Mana Rugbumrung (DMR) for his help in identifying some fossil specimens and for his technical assistance. I thank Pannipa Tian (DMR), Chotima Yamee (DMR), and Bernard Marandat (ISEM) for their assistances in the fieldwork. I also thank Supparerk Ruckchart for his assistance with statistical analyses. I would like to express my sincere appreciation to the laboratory secretaries: Ghislaine Florent, Guylaine Reynaud, and Carine Noël (former), who always organized transportation and accommodation abroad for my internships, scientific collection visiting, and fieldworks. I take this opportunity to record my sincere thanks to all members of the iPHEP laboratory (Université de Poitiers) and the Animal Systematics Research Unit (Chulalongkorn University) for scientific discussions and communications, as well as for our friendship.
I am obligated to my dad, mom, and brothers, who gave me financial and moral supports, as well as encouragement, when I needed them. This work is dedicated to my second mom, Thongmuan Puangsri (Armooh), who is without a doubt one of the most important person in my life. In addition to her assistance with the data collection, she made many sacrifices to raise me up and had a positive influence on my life since I was born. I especially thank Armand and Michel for supporting me everything. I also thank my beloved friends and cousins: Aphinand (Sun), Chatchalerm (Kendo), Manit (Meng), Chonlada (Kate), Patchara (P’Ae), Chawin (P’Nam), Karanta (Fern), and Suppharun (Champ), who graciously helped me collect the data at the museums and generate the PhD thesis database. I especially thank Preusamon for her encouragement and dissertation assistance. I really enjoyed sharing an office with Salamet and Florian who stayed together during the late-night work. I am beyond grateful to have all friends here, including Tiphaine, Anne, Abdou, Ghislain, Dimitri, Emilie, Clarisse, Mikael, Yamina, Mathieu, God, Weddy, Sonia, Lisa, Fabien, Johanne, and Vincent, who always taught me everything about the French language, culture, food, and life, and the coursework of Master’s degrees (M1 and M2). I especially acknowledge all academic institutitions and departments for all facilities and financial supports through the research study: iPHEP (Université de Poitiers) and Chulalongkorn University, and for allowing me access to the collection: the Department of Mineral Resources and the Khok Sung subdistrict municipality. Finally, this research could not have succeeded without the financial supports funded by the following grants: “The 90th Anniversary of Chulalongkorn University fund”,
impacts on the biodiversity and the species composition and distribution: the contribution of Khok Sung (Khorat province, Thailand) vertebrate community
The Khok Sung sand pit, Nakhon Ratchasima province, has yielded the richest Pleistocene vertebrate fauna of Thailand, where more than a thousand fossil mammals and reptiles (skulls, isolated teeth, and postcranial remains) were recovered. The mammalian fauna, which is described in details hereby, consists of at least 17 identified species (12 genera), including a primate, proboscideans, rhinoceroses, suids, bovids, cervids, and carnivores, which are characterized by mostly extant elements associated to some completely and locally extinct taxa. The age of the Khok Sung fauna is tentatively attributed to the late Middle Pleistocene as either 188 or 213 ka, based on the paleomagnetic data and on the faunal comparisons. The Khok Sung mammal assemblage yields the Stegodon-Ailuropoda faunal association, most similar in composition to that of Thum Wiman Nakin, supporting the hypothesis that northeastern Thailand was a biogeographic gateway of the Sino-Malayan migration route from South China to Java. An analysis of stable carbon isotopes extracted from the tooth enamel of fossil ungulates reveals evidence of niche partitioning among megaherbivores and within cervids. The enamel carbonate 13C values also suggest a considerable
amount of C4 plants in the dietary use of ruminants, indicating that grasslands had significantly expanded in Thailand at that time during which anthropic impacts on the ecosystems were absent. The stable oxygen isotope results, obtained from the serial sampling of large mammal enamel, combined with the cenogram analysis reflect significant seasonal variation in precipitations and temperature for Khok Sung, associated to a relatively humid climate.
Keywords: large mammals, biochronology, paleobiogeography, paleoenvironments, paleoclimate, Middle Pleistocene, Quaternary, Thailand, mainland Southeast Asia
Paléoenvironnements et reconstitutions paléoclimatiques du Pléistocène moyen de Thaïlande et leur impact sur la biodiversité et la distribution des espèces: la contribution de la faune de vertébrés du gisement de Khok Sung (Province du Khorat)
La sablière de Khok Sung, dans la province de Nakhon Ratchasima, qui a livré plus d'un millier de fossiles de mammifères et de reptiles (cranes, dents isolées et restes post-craniens), abrite la faune de vertébrés du Pléistocène la plus riche de Thaïlande. La faune mammalienne qui est décrite ici en détail, se compose d'au moins 17 espèces identifiées (12 genres), y compris un primate et des proboscidiens, rhinocéros, suidés, bovidés, cervidés et carnivores. Elle compte principalement des taxons encore représentés de nos jours, ainsi que quelques taxons globalement ou localement éteints. A partir des données paléomagnétiques et des comparaisons fauniques, l'âge de la faune de Khok Sung est estimé au Pléistocène Moyen tardif, vers 188000 ou 213000 ans. Par rapport aux autres faunes diversifiées du Pléistocène d'Asie du Sud Est, l'assemblage de Khok Sung est caractérisé par une association des taxons Stegodon-Ailuropoda, comparable en cela au site de Thum Wiman Nakin, ce qui supporte l'hypothèse selon laquelle le Nord Est de la Thaïlande était un corridor biogéographique appartenant à la route migratoire Sino-Malaise, entre la Chine du Sud et l'île de Java. L'analyse des isotopes stables du carbone à partir de l'émail des ongulés fossiles révèle la présence d'une partition de niches entre les méga-herbivores et au sein des cervidés. Les valeurs du 13C de
l'émail suggèrent également que les ruminants ont consommé une grande quantité de plantes en C4, ce qui indique que les prairies à graminées étaient particulièrement répandues en Thaïlande à cette époque où les écosystèmes n'étaient pas encore soumis à l'influence anthropique. La mesure des isotopes stables de l'oxygène, obtenue par échantillonnage sérié de l'émail des dents de grands mammifères, et l'analyse du cénogramme de la localité de Khok Sung reflètent une importante variation saisonnière des précipitations et de la température, associée à des conditions climatiques relativement humides.
Mots-clés: grands mammifères, biochronologie, paléobiogéographie, paléoenvironnements, paléoclimat, Pléistocène moyen, Quaternaire, Thaïlande, Asie du Sud-Est continentale
Acknowledgments
Abstract
Introduction
I.1) Problematic aspects of paleoenvironmental and paleoclimatic studies in mainland
Southeast Asia 1
I.2) The discovery of the Khok Sung fossiliferous site 2
I.3) Regional geologic setting 5
I.4) Objectives of the study 6
I.5) Thesis plan 7
I.6) Southeast Asian history and biogeographic provinces 8 I.7) Zoogeographic history of Southeast Asian mammalian faunas 11 I.8) Pleistocene mammalian fossil sites in mainland Southeast Asia 14
I.8.1) Myanmar 15
I.8.2) Laos 17
I.8.3) Cambodia 18
I.8.4) Vietnam 18
I.8.5) Peninsular Malaysia 20
I.8.6) Thailand 21
Chapter 1: A complete skull of Crocuta crocuta ultima indicates a late Middle
Pleistocene age for the Khok Sung (northeastern Thailand) vertebrate fauna
1.1) Abstract 35
1.2) Introduction 36
1.3) Geological setting and previous chronological framework 39
1.4) Materials and Methods 42
1.5) Systematic paleontology 42
1.8.1) Age and paleobiogeographic records of C. crocuta in Asian mainland and adjacent
areas 52
1.8.2) Contribution to the age of Khok Sung fauna 55
1.8.3) Paleoecological and paleoenvironmental implications of Khok Sung locality 56
1.9) Conclusion 57
Chapter 2: The Middle Pleistocene vertebrate fauna from Khok Sung (Nakhon
Ratchasima, Thailand): biochronological and paleobiogeographical implications
2.1) Abstract 70
2.2) Introduction 71
2.3) Material and methods 74
2.4) Systematic paleontology 76
- Stegodon cf. orientalis 76
Material description 76
Taxonomic remarks and comparisons 80
- Elephas sp. 86
Material description 86
Taxonomic remarks and comparisons 86
Past and current distribution of stegodontids and elephantids 87
- Rhinoceros sondaicus 90
Material description 90
- Rhinoceros unicornis 94
Material description 94
Taxonomic remarks and comparisons of Khok Sung Rhinoceros 94
Past and current distribution of Indian and Javan rhinoceroses 96
- Bos sauveli 99
Material description 99
- Bos javanicus 101
- Bubalus arnee 106
Material description 107
Taxonomic remarks and comparisons of Khok Sung large bovids 109
Past and current distribution of Southeast Asian large bovids 126
- Capricornis sumatraensis 129
Material description 129
Taxonomic remarks and comparisons 130
Past and current distribution of Capricornis sumatraensis 131
- Axis axis 133
Material description 134
Taxonomic remarks and comparisons 137
Past and current distribution of Axis axis 142
- Panolia eldii 143
Material description 143
- Rusa unicolor 147
Material description 147
Taxonomic remarks and comparisons of Khok Sung large cervids 150
Past and current distribution of Panolia eldii and Rusa unicolor 153
- Sus scrofa 155
Material description 155
- Sus barbatus 157
Material description 157
Taxonomic remarks and comparison of Khok Sung Sus 158
Past and current distribution of Sus scrofa and Sus barbatus 160
- Macaca sp. 163
Material description 163
Taxonomic remarks and comparisons 165
- Crocodylus cf. siamensis 168
Material description 168
Taxonomic remarks and comparisons 170
- Python sp. 170
Material description 170
Taxonomic remarks and comparisons 171
- Varanus sp. 172
Material description 172
Taxonomic remarks and comparisons 173
2.5) Faunal composition of Khok Sung vertebrate assemblage 173 2.6) Mammalian fauna comparisons: description and Raup-Crick similarity analysis 176
2.7) Discussion 180
2.7.1) Age of Khok Sung fauna 180
2.7.2) Relevance of the Khok Sung vertebrate fauna for the dispersal from mainland
Southeast Asia to Java 180
2.8) Conclusion 184
Chapter 3: Paleoecological and paleoclimatic reconstructions of Pleistocene
Khok Sung vertebrate fauna (Nakhon Ratchasima province, Northeastern
Thailand): stable carbon and oxygen isotope investigations of ungulate tooth
enamel
3.1) Abstract 263
3.2) Introduction 264
3.3) General principles and applications of stable isotopes in the study of paleoecology
and paleoenvironments 266
3.3.1) Stable carbon isotopes in mammalian diets and habitats 266
3.3.2) Stable oxygen isotopes in tooth enamel of mammals 268
3.3.3) Intra-tooth sampling: records of seasonal variation 269
3.4.3) Cenogram application 272
3.5) Results and interpretations 272
3.5.1) Bulk carbon and oxygen isotope analyses 272
3.5.1.1) Bovids 275
3.5.1.2) Cervids 275
3.5.1.3) Rhinoceroses 277
3.5.1.4) Proboscideans 278
3.5.1.5) Suids 279
3.5.2) Serial carbon and oxygen isotope records from the Khok Sung large mammals 279
3.5.3) Cenogram analysis 283
3.6) Discussion 284
3.6.1) Spatial and temporal C3/C4 vegetation and paleoenvironments in eastern Asian mainland during the Pleistocene: evidence from the stable carbon isotopes of mammalian
tooth enamel 284
3.6.1.1) China 285
3.6.1.2) South China 285
3.6.1.3) Mainland Southeast Asia 286
3.6.2) Niche partitioning of sympatric Khok Sung ungulates 289
3.6.3) Niche partitioning between late Middle Pleistocene and modern large mammals in
Thailand 291
3.6.4) Paleoclimatic implications of Khok Sung 291
Chapter 4: Conslusions and future perspectives
4.1) Age 306
4.2) Taxonomy and faunal composition 308
4.3) Paleobiogeography 309
4.4) Paleoecology, paleoenvironments, and paleoclimate 311 4.5) Taphonomic and paleoanthropological perspectives 313
Introduction
I.1) Problematic aspects of paleoenvironmental and paleoclimatic studies in mainland Southeast Asia
The Middle Pleistocene (781 to 126 ka) is a critical period in the history of Earth characterized by a cyclic occurrence of high amplitude glacial periods alternating with interglacial periods (Huybers, 2007), whose intensity is under the control of astronomic cycles and oceanic paleocurrents. The climatic characteristics of that period have been largely analyzed in the Northern Hemisphere, where the glaciations are directly expressed by the variation of inlandsis surface and extension (e.g., Varsteegh, 1997; Jahn et al., 2003). Paleoclimatic evidence has been also deeply investigated in the world oceans (e.g., Billups and Schrag, 2003; Ferreira et al., 2014; DeNinno et al., 2015) because of the possibility of having access to more global information, which is less affected by continental and coastal conditions.These important data contribute to understanding of the climatic changes that have occurred in the world. The impact of these climatic events on the biodiversity is also well-known in the Northern hemisphere continents and surrounding oceans, having considerably modified the composition of the living communities, by the way of distribution, extinction, and speciation events, reduction and/or extension of landscapes and habitats. However, the understanding of these events is much less advanced in tropical areas. In Africa, in relation to human evolution, a great number of data have been collected so far and have shown the close relationships between vertebrate and plant communities, composition, distribution, and climatic changes. It is now clear that, at least in tropical Africa, the glacial periods corresponded to significantly more open environments than during the interglacial phrase, stable isotopes and microfossils from lake sedimentary cores having broadly contributed to that understanding (e.g., Smith et al., 2004; Lézine et al., 2005; Lee-Thorp et al., 2007). On the other hand, the current situation is much more confused in Asia, where only two regions (Northern China and Indonesia) have been investigated in details. In Northern China, the study of loess deposits (e.g., An et al., 1990; Sun et al., 2006; Peng et al., 2015) and the stable isotopic analysis of terrestrial mammal tooth enamel (e.g., Gaboardi et al., 2005; Biasatti et al., 2010) have allowed to unravel some aspects of climatic changes in the Northern part of Asia. Some proxy data have been obtained from Sundaland (Indonesia), mostly the island of Java, in relation to the rich records of fossil hominids (Homo erectus) (Dubois, 1894) in that island (e.g., van der Kaars and Dam, 1995, 1997; van der Kaars, 1998;
Bettis et al., 2009; Sémah et al., 2010; Brasseur et al., 2015). However, the data obtained from extensive chronological stages (Early to Late Pleistocene) are still scarce, often limited to short time intervals, and become only more detailed for the late Pleistocene, since 126,000 years ago.
Since Southeast Asia has been currently known as a “biodiversity hotspot” with a high diversity of plants and animals including rare and unique species and/or even specific landforms, glacial refugia could contain the last remaining individuals of species that were widely distributed but have now mostly disappeared. It is possibly hypothesized that some population of animals and plants have been separated from the rest of their species within these refugia in this region during the glacial events and have subsequently increased or decreased their genetic diversity. In Thailand and other countries, this information is poorly known due to the extreme scarcity of data. In addition, this current geographic area is under the control of the monsoon, which is classically considered to have been directly related to the uplift of the Tibetan plateau since the last 20 million years ago (Ruddiman and Kutzbach, 1989, 1990; Passey et al., 2009). The paleobotanic evidence obviously demonstrates that the vegetation zones dropped of about 1000 meters of elevation and that lowlands were mostly covered by grasslands with tropical Dipterocarp forests relicts in some areas during the last glacial maximum (e.g., Morley, 1982, 2012; Sun et al., 2002; Harrison and Prentice, 2003; Raes et al., 2014). However, the fragmentation of the Dipterocarp forest did not reach the maximum levels at all of the periods, as exemplified by the fact that orangutans and giant pandas were widespread in mainland Southeast Asia (e.g., Olsen and Ciochon, 1990; Tougard et al., 1996, Tougard and Ducrocq, 1999; Ibrahim et al., 2013; Harrison et al., 2014). But this iterative mechanism of contraction and extension of Dipterocarp forests has been considered as the source of numerous allopatric speciation events whose evidence has to be still demonstrated. Therefore, the fauna and flora enclosed on the Pleistocene deposits continue to play an important role to understand these environmental and climatic phenomena.
I.2) The discovery of the Khok Sung fossiliferous site
The Khok Sung fossiliferous site (N 15°06'17", E 102°06'38.2") is situated in land tenure of the Korat Yongsanguan Rice Mill Co. Ltd. owned by Mr. Somchai Tirasetphakdee. This rice mill is located in Ban Khok Sung subdistrict, Nakhon Ratchasima province (also
called “Khorat”), at about 15 km north of Nakhon Ratchasima city, close to the Highway no. 205 (Fig. I.1).
Figure I.1. Map of Thailand showing the location of the Khok Sung fossiliferous site in Nakhon Ratchasima province, northeastern Thailand (modified from the Royal Thai Survey Department topographic map: scale 1:50,000, sheet 5439III, series L7017). Red lines indicate highways of Thailand.
In 2005 (March), remains of vertebrate fossils have been first recovered, at the depth of 5-7 meters below the surface, during which the fluviatile terrace deposits behind the rice
mill were digging out for the construction of a reservoir (50 long × 50 wide × 15 deep) (Fig. I2A). Following the discovery of large bone fragments by workers, this site was then rapidly excavated by the Thai-French paleontological research team and geologists from the Department of Mineral Resources. While the water was pumped out of the sand pit, the fossils have been continuously exposed and searched by using the water spraying technique (Fig. I2B). The excavation work has revealed an extraordinary discovery of vertebrate fossils, particularly mammals and reptiles, in terms of quantity and preservation (Fig. I2C).
Figure I.2. The sand pit of Khok Sung during the paleontological excavation (in March, 2005): (A) a general view of the sand pit, (B) an area of vertebrate fossils recovered, and (C) in situ remains of Stegodon. The photos were taken by Yaowalak Chaimanee during the fieldwork. The blue arrow indicates the location of the fossiliferous layer (above the dark gray layer) and the black one points to the non-marine Mesozoic bedrock.
I.3) Regional geologic setting
The Khok Sung sand pit is geographically positioned on the Khorat Plateau (about 150 m above sea level for an average elevation), which is located at the north-central part of the Indochinese Peninsula. The Khorat Plateau, which covers an area of 180,000 km2,
exhibits a saucer-shaped basin that was structurally developed over the sequence of Mesozoic to Tertiary clastic sediments (Sattayarak, 1985; Sattayarak et al., 1998). It fundamentally comprises two major sedimentary basins, the large Khorat basin (south) and the small Sakon Nakhon basin (north), separated by the northwestern to southeastern Phu Phan mountain range (Dheeradilok and Kaewyana, 1986). The central part of the southern Khorat basin with an average elevation of 120 m contains a deeply incised mature river valley, which has been filled by Quaternary sediments (Löffler et al., 1984). Other incised valleys covered by more recent sediments indicate that the flat appearance of the Khorat Plateau is today characteristics of the infilling sediments during the Late Pleistocene to Holocene (Löffler et al., 1983). The Quaternary tectonic activity on the Khorat Plateau has therefore contributed to the alteration of present-day river drainage patterns (Löffler et al., 1983; Hutchison, 1989).
The Plateau’s landscape is drained by two main rivers, Mun and Chi, which flow eastward into the Mekong River. The Mun River drainage joins the Mekong River near the eastern border of the Khorat Plateau. However, the ancient Mun river drainage is supposed to have been also connected to the Chao Phraya River prior to the Middle Pleistocene (Hutchison, 1989; Attwood and Johnston, 2001; Glaubrecht and Köhler, 2004). Its paleocurrent was possibly flowing from east to southwest, in the direction of Chao Phraya drainage systems (Claude et al., 2011). The Khok Sung sand pit is located closest to the Mun River and has been therefore considered as corresponding to the ancient Mun River terrace deposits, which consist of Quaternary alluvial sediments underlain by the Mesozoic red beds (Chaimanee et al., 2005) (Fig. I2A). The undulating bedrock surfaces underlying the Mun River floodplain and the interfluvial areas are a result of the weathered shales and sandstones of the Mahasarakham Formation (Löffler et al., 1984) (see the next chapter for the detailed geological context of the locality).
I.4) Objectives of the study
The proposed subject of this research is mainly focused on the understanding of the paleoenvironments and paleoclimates in mainland Southeast Asia during the Pleistocene based on the study of mammalian fossils recovered from the Khok Sung sand pit, Nakhon Ratchasima province in northeastern Thailand. Three main objectives of the thesis have been proposed as follows;
First, to collect all available data and to review literatures concerning the Pleistocene mammal faunas of Southeast Asia, in order to improve our knowledge regarding the history of the extant fauna and the faunal exchanges that have occurred during the Pleistocene, to compare the geographic distribution ranges of mammals between the Pleistocene and the present day, and to analyze the possible causes of extinction.
Second, to identify precisely the taxonomic status of these vertebrate fossils, which are exceptionally well-preserved and complete, in opposition to fragmentary remains of the cave deposits. This would allow to elaborate a new and complete fauna list of Khok Sung, only some turtles, soft-shelled turtles, and gharials having been previously described. The completeness of several species remains, which are represented by complete skulls, horns and antlers, tooth rows, and postcranial bones will constitute a reference for the identification of the karst fissure fragmentary bones and teeth in the future.
The material is currently stored in the Department of Mineral Resources (DMR), Bangkok and also in the local Museum of the village (Ban Khok Sung), Nakhon Ratchasima province. The age of the Khok Sung fauna proposed here is based on the biochronological information coupled with paleomagnetic data. The fauna composition of Khok Sung could be compared in details with other Middle to Late Pleistocene faunas that have already been published from South China, Myanmar, Laos, Vietnam, Cambodia, and Peninsular Malaysia, as well as from other localities in Thailand. Differences in geographic distribution of vertebrate taxa between the Pleistocene and today may reveal more details regarding the nature of climatic changes that took place during that period.
The third objective of this study is to analyze stable isotopes (13C and 18O) of tooth
enamel of these mammals in order to constraint their paleodiets (exclusive C3, C4 plants or mixed diet) and habitats (open, closed or intermediate landscape), to evaluate niche partitioning of Khok Sung vertebrate community, and to reconstruct the air temperature,
humidity, and animal-drinking behavior. High resolution sampling through growth lines along mammalian hypsodont crowns may also help to quantify the importance of the seasonality. These isotopic results could be compared with data from other localities in Southeast Asia and from other parts of the world. Finally, the cenogram construction based on the weight distribution of the Khok Sung mammals could provide information regarding the paleoenvironments of the locality, possibly allowing a higher resolution of paleoclimatic interpretation of the region, when combined with those of stable isotopic results.
Overall, the amount of proxy data obtained through this study would allow to reconstruct the regional paleoenvironments and paleoclimates, which have been poorly known, especially for most of the tropical old world countries at least during the Middle to Late Pleistocene. Finally, this research may shed some critical new light on the natural events that affected and shaped extant Southeast Asian vertebrate communities during the Pleistocene and that led to better understand the origin of present-day faunas and floras in mainland Southeast Asia.
I.5) Thesis plan
This thesis has been divided into 4 main chapters (three research studies and a summary of the results) regarding the chronological, taxonomic, paleobiogeographical implications of the Khok Sung fauna and the paleoenvironmental and paleoclimatic reconstructions of that locality.
Chapter 1: A complete skull of Crocuta crocuta ultima indicates a late Middle Pleistocene age for the Khok Sung (Northeastern Thailand) vertebrate fauna
This chapter presents the results of the taxonomic study of the fossil spotted hyaena, whose material is highly complete, recovered from the Khok Sung sand pit. The spotted hyaena fossil, which represents a good biochronological index for the Middle to Late Pleistocene, has been used here to estimate the age of the fauna. In addition, I combined paleomagnetic data with biochronological information in order to determine more precisely the age of the fauna. These results have been officially published in the journal of “Quaternary International”.
Citation: Suraprasit, K., Jaeger, J.-J., Chaimanee, Y., Benammi, M., Chavasseau, O., Yamee, C., Tian, P., Panha, S., 2015. A complete skull of Crocuta crocuta ultima indicates a late Middle Pleistocene age for the Khok Sung (northeastern Thailand) vertebrate fauna. Quaternary International 374, 34–45.
Chapter 2: The Middle Pleistocene vertebrate fauna from Khok Sung (Nakhon Ratchasima, Thailand): biochronological and paleobiogeographical implications
The main goal of this chapter is to describe a large amount of the Khok Sung fossil specimens in terms of taxonomy in order to constraint the age of the fauna based on the faunal comparisons with other Pleistocene localities and to better understand the paleobiogeographic affinities of Pleistocene large mammal faunas across Southeast Asia. This part is considered to be published as a monograph, which is able to present comprehensive taxonomic details of the fauna.
Chapter 3: Paleoecological and paleoclimatic reconstructions of Pleistocene Khok Sung vertebrate fauna (Nakhon Ratchasima province, Northeastern Thailand): stable carbon and oxygen isotope investigations of ungulate tooth enamel
In the context of this chapter, I described the paleoecological significances of Khok Sung ungulates and provided the paleoenvironmental and paleoclimatic informations obtained from the stable carbon and oxygen isotope analyses of fossil tooth enamel. This part has been designed as a form of scientific research articles for publishing.
Chapter 4: Conclusions and future perspectives
Based on the obtained data from this study, I finally summarized the main results and gave more comprehensive ideas on some geological and paleontological applications in order to better understand all these important aspects and/or to extend our results into other related fields.
I.6) Southeast Asian history and biogeographic provinces
Southeast Asia constitutes a subregion of Asia and is divided into two geographic regions (mainland and insular Southeast Asia). Mainland Southeast Asia consists of the countries including Myanmar, Thailand, Peninsular Malaysia, Laos, Cambodia, and Vietnam,
whereas insular or maritime Southeast Asia includes the countries of Malaysian Borneo, Singapore, Brunei, East Timor, Indonesia, and the Philippines. The Southeast Asian continental block largely comprises elements, which have been broken off from the southern supercontinent Gondwanaland (Gatinsky and Hutchison, 1987). Tectonically, the extant geography of Southeast Asia has been formed by the collision between Sinoburmalaya and Cathaysia plates from the event of the Late Triassic Indosinian Orogeny (Hutchison, 2005). Additionally, the collisions between Burma plate and Shan highlands during the Cretaceous and between India and Eurasia during the Early Tertiary have been evidenced (Hutchison, 2005). Until the Miocene, the latter collision gradually resulted in the North-South trending mountain ranges of Western Yunnan, Myanmar, and Peninsular Malaysia at the eastern part and in the uplift of the Himalayas and the Qinghai-Tibetan Plateau at the northern part (Whitmore, 1987). The uplift of the Himalayas and the Qinghai-Tibetan Plateau has caused the alteration of climates, resulting in the development of the Asian monsoon system during the Miocene (25-22 Ma). However, the existence of the older Asian monsoon related to the enhanced greenhouse conditions is supposed to have occurred in north and southern part of the Tibetan-Himalayan orogen during the late Eocene (Licht et al., 2014). Another relatively more recent tectonic collision occurred between Southeast Asia and Australia at about 15 Ma, leading to the formation of the Lesser Sunda islands, as well as the appearance of islands of Sulawesi and the Philippine Archipelago (Hutchison, 1989).
With regards to biogeographical terms, Southern Asia coincides with the Indo-Malayan region (Udvardy, 1975), which is divided into 5 subregions: the Indian, Indochinese, Sundaic, Philippines, and Wallacean subregions (Lekagul and McNeely, 1988; Corbet and Hill, 1992). The Indochinese subregion includes the Indochinese Peninsula (Myanmar, Thailand, Laos, Cambodia, and Vietnam and South China), whereas the Sundaic subregion comprises the southern part of Thailand, Malaysia, Sumatra, Java, and Borneo (Lekagul and McNeely, 1988) (Fig. I3).
The climate between these two subregions is markedly different, the Indochinese province displaying a stronger seasonality with lower rainfall (Whitmore, 1984; Gray et al., 1994). According to Wallace (1869), biogeographers have defined a major transition between Indochinese and Sundaic zoogeographic subregions at the Thai-Malay Peninsula near the Isthmus of Kra (10° 30´ N, Peninsular Thailand), where both of the northern and southern faunas meet (Fig. I3). This biogeographic boundary is recognized by differences in biotas, with a rapid turnover. There is about 50% turnover at the species level in forest-associated
birds between the northern and southern of the Kra Isthmus (Hughes et al., 2003; Round et al., 2003). Botanists recognized a similar floristic pattern but more significant transition at about 500 km south, where the Southeast Asian and Malayan plants meet (e.g., Whitmore, 1984; Lekagul and Roung, 1991; Ashton, 1992; Baker et al., 1998; Morley, 2000). These differences in biotas are also valid for insects, amphibians, and mammals (e.g., Corbet and Hill, 1992; Corbet and Pendlebury, 1992; Gray et al., 1994; Inger, 1999).
Figure I.3. Map of Southern Asia showing the biogeographical regions and subregions (modified after Lekagul and McNeely (1988), Corbet and Hill (1992), and Antoine (2012)).
The mammalian fauna is sufficiently rich to support biogeographic analyses due to the presence of over 500 extant species in Southeast Asia. The extant mammal transition is also suggested to occur near the Isthmus of Kra (Chasen, 1940; Musser and Newcomb, 1983; Cranbrook, 1988; Lekagul and McNeely, 1988; Corbet and Hill, 1992). The species diversity of national terrestrial mammals in Southeast Asia has been estimated: ~300 species in Myanmar, ~251 species in Thailand, ~273 species in Vietnam, and ~210 species in Malaysia (WCMC, 1992; SAMD, 2006; Sterling et al., 2006). In addition, the taxonomic abundances, habitats, distributions, and natural history of these mammals have been documented by a century of enquiry, atlases, and books (Chasen, 1940; Medway, 1983; Lekagul and McNeely,
1988; Corbet and Hill, 1992; Francis, 2001; 2008). However, the understanding of the history of present-day mammals in the region is most linked to information on the Pleistocene megafaunas (e.g., biodiversity, distribution, and extinction), whose evidence is still under investigation today. The Khok Sung fauna, which has yielded numerous remains of large mammals, possibly provides a high amount of data regarding all of those aspects.
I.7) Zoogeographic history of Southeast Asian mammalian faunas
Since the late 20th century, the Quaternary history of Southeast Asia is known to have
been sufficiently detailed and to have allowed for some reliable inferences of the role of the geographic events in faunal distribution. Several studies (e.g., Chaimanee, 1998; Chaimanee and Jaeger, 2000; Tougard, 2001; Tougard and Montuire, 2006) on the Pleistocene mammalian faunas in mainland Southeast Asia made the significant progress in explaining paleobiogeographic affinities and could be considered as references to the broader faunal comparisons. Fossils of murine rodents and squirrels are useful to reconstruct the regional palaeoenvironments and their data can be interpreted as an indicator for the past boundaries of forest and savannah communities in mainland Southeast Asia(Chaimanee, 1998; Tougard and Montuire, 2006). They also have evidenced past migrations between Indochinese and Sundaic subregions, which occurred in Thailand. Distribution patterns inferred from northern taxa that dispersed southward into the Sundaic subregion and from some peninsular endemic taxa, whose distribution has taken place in the Northern Province, suggest severe shifts of the boundary betweenthese two biogeographic subregions during the Pleistocene. Similar to those of large mammals, their fossil records have indicated that the transition between Indochinese and Sundaic taxa was located further south of the Isthmus of Kra during some parts of the Pleistocene. Some species dispersed further north or south than they do today (e.g., Tougard, 2001; Bacon et al., 2004; 2006; 2008a; 2008b; 2011), in relation to the climatic fluctuations during the glacial-interglacial. Overall, mainland Southeast Asia appears to have been cooler and more seasonal than today during the major part of the Pleistocene (Chaimanee, 1998; 2007; Penny, 2001; White et al., 2004). The vegetation type of mainland Southeast Asia during that period is likely to have been dominated by evergreen, semi-evergreen and coniferous forests with considerable amounts of grasses and some herbaceous vegetation (Louys and Meijaard, 2010). However, the paleogeographic situation in mainland Southeast Asia is still less advanced, compared to those of islands (e.g., Java), where the
Pleistocene faunas and floras have been studied in more details in relation to the presence of Homo erectus. The paleodistribution of large terrestrial mammals in insular Southeast Asia has been well-documented, almost related to a land corridor (known as the “Sunda shelf”) connecting the mainland Southeast Asia and islands of Indonesia (Fig. I4), allowing an overland migration of large mammals during the glacial periods. Glacio-eustatic fluctuations have therefore played a significant role on Southeast Asian geography (e.g., size, number, and degree of isolation of islands) and biota (e.g., faunal turnover and endemism rate), depending on the timing and magnitude of sea level related to the glacial and interglacial climatic conditions.
Figure I.4. Map of Southeast Asia showing the land exposure (sky blue color), at sea level of 120 m below the present day, over Sundaland and Indochina during the Pleistocene (modified from Voris (2000)).
The Sundaland is a biogeographical region that comprises the Indonesian islands of Borneo, Sumatra, Java, and Bali, as well as Malay Peninsula. The largest geographic part of this region lies hidden below sea level today. Early geographic emergence of the Sunda shelf (geological term) or “Sundaland” (biogeographical term) in Southeast Asia has been first
reconstructed by Wallace (1876) and Molengraaff (1922), and has been further studied and discussed by many authors (e.g., Whitmore, 1987, 1998; Hall and Holloway, 1998; Woodruff, 2003; Bird et al., 2005; Meijaard and Groves, 2006). Although many details remain controversial, the major patterns of geographic and sea-level changes are principally accepted now. Early Pleistocene faunas related to the expansion of the Sundaland are almost known exclusively from Java, where the uplift process started during the Late Pliocene due to the combination of tectonic and volcanic activities (van Bemmelen, 1949). Both the flora and the fauna between the mainland and insular Southeast Asia are supposed to have been exchanged during the Early Pleistocene (before 800 ka) by dispersing through the emergence of Sundaland, when the sea level dropped of about 70 m on average below the present day (Prentice and Denton, 1988; van den Bergh et al., 2001). These archaic fossils recovered from the Sangiran Formation indicate an unbalanced fauna and suggest Siva-Malayan characteristics that originated from Siwaliks and Myanmar (Sondaar, 1984). The paleoenvironmental conditions during the Early to Middle Pleistocene in Java were characterized by riparian forests, savannahs, and open woodlands (Bettis et al., 2009; Sémah et al., 2010). A major environmental change that occurred in Java might have taken place toward the end of the Middle Pleistocene, as exemplified by the presence of new tropical rainforest faunas in Punung (van den Bergh et al., 2001; Westaway et al., 2007). The large expansion of Sundaland could generally provide an unequal opportunity for the distribution of mammals because forest-associated mammals are suggested to be better adapted to various types of forests (e.g., perhumid rainforest and seasonal, monsoonal, and mangrove forests) (Woodruff and Turner, 2009). The floodplain area of Sundaland might have been vegetated by Pinus savannah woodlands or grasslands, rather than by the rainforests (Whitmore, 1987, 1998; Heaney, 1991; Morley, 2000, 2007). Therefore, mammals that are restricted to the primary rainforests could have crossed this area through riparian forest corridors and subsequently became isolated by ecological barriers (Gorog et al., 2004). However, the central Sundaland that was exposed due to the lower sea level during the last glacial maximum (LGM, 21 ka) harbored optimal environmental conditions for Dipterocarpaceae and was probably covered by rainforests (Raes et al., 2014).
Although the boundary and range of the distribution of savannah lands and rainforests in Southeast Asia are still controversial (Sun et al., 2000, 2003; Meijaard, 2003; Bird et al., 2005; Morley, 2007; Raes et al., 2014), the impacts of the habitat modification on the past mammalian geographic distribution are linked directly to these scenarios. Studies on the
paleodiets and habitats of ancient mammals are therefore crucial to understand their distribution patterns and responses to environmental and climatic changes and needs to be taken into account in future analyses. Here we summarize the key findings from literature works and also elucidate an ongoing progress on the Pleistocene fossil records of large mammals in mainland Southeast Asia, in terms of faunal composition, chronology, paleoenvironments, and paleoclimate.
I.8) Pleistocene mammalian fossil sites in mainland Southeast Asia
A particular assemblage of mammals, so-called the “Ailuropoda-Stegodon fauna complex” or the “Sino-Malayan fauna”, has been known since the early 20th century and first
described as a representative of the Middle Pleistocene in South China (Young, 1932; Pei, 1935; Teilhard de Chardin, 1935; Bien and Chia, 1938; Granger, 1938; von Koenigswald, 1939; Kahlke, 1961; de Vos, 1984). However, it is now proven to have represented a characteristic of the long period ranging from the Early to Late Pleistocene of the Indochinese subregion (e.g., Tougard, 2001; Rink et al., 2008; Zeitoun et al., 2010; Bacon et al., 2011; 2015). This faunal association fundamentally yields Asian taxa, which are endemic or closely related to tropical environments, including Stegodon, Indian elephants, rhinoceroses, Gigantopithecus, orangutans, suids, cervids, and bovids. The carnivores commonly include a spotted hyaena, a tiger, a panther, a dhole, an Asiatic black bear, and a giant panda. This faunal complex has been first identified in South China (Matthew and Granger, 1923), based on the occurrence of tropical taxa such as Hylobates and Tapirus, followed by the same discovery in Vietnam (Patte, 1928), Laos (Fromaget, 1936), Myanmar (de Terra, 1938), Cambodia (Beden and Guérin, 1973), and Thailand (Pope et al., 1981; Ginsburg et al., 1982). The distribution of the faunal complex is widespread in Indochinese subregion, expanding latitudinally from the Yangtse River (Pei, 1957) to Kra Isthmus (Tougard, 2001). Their fossils have been mostly found from the karst topolographies including sinkholes, caves, and underground drainage systems. Interestingly, this faunal association was sometimes recovered together with human remains and/or stone artefacts, allowing further the understanding of the anthropic dynamics in this part of the world. The Ailuropoda-Stegodon faunal characteristics are highly crucial to establish a regional biochronology and stratigraphy, at least for the Pleistocene. However, nowadays its precise duration and subdivision are peculiarly nonspecific. In mainland Southeast Asia, the absolute datings are
scarce and relatively inaccurate. Bacon et al. (2015) proposed a Middle to Late Pleistocene biochronological division in mainland Southeast Asia with three evolutionary stages: (1) the presence of lineages of extant mammals (modern faunas), (2) the occurrence of some extinct taxa (e.g., Megatapirus augustus and Stegodon orientalis) (archaic faunas), and (3) the appearance of new incomers. However, the biochronological ages based on the evolutionary stages have not been justified yet. A precise biochronologic time scale will become critical to reconstruct scenarios of the mammalian evolution and extinction, as well as paleoenvironments and paleoclimates of the region, if such a well constrained timing is more finely subdivided.
According to the numerous paleontological and archaeological discoveries of Pleistocene fossil site in mainland Southeast Asia since the late 20th century, new data have
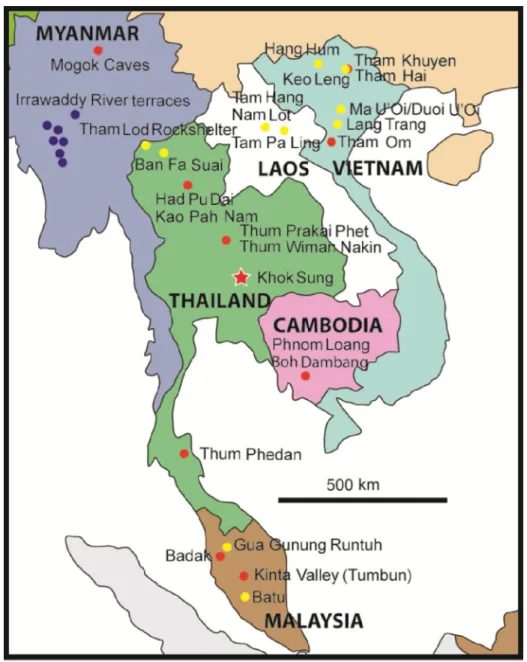
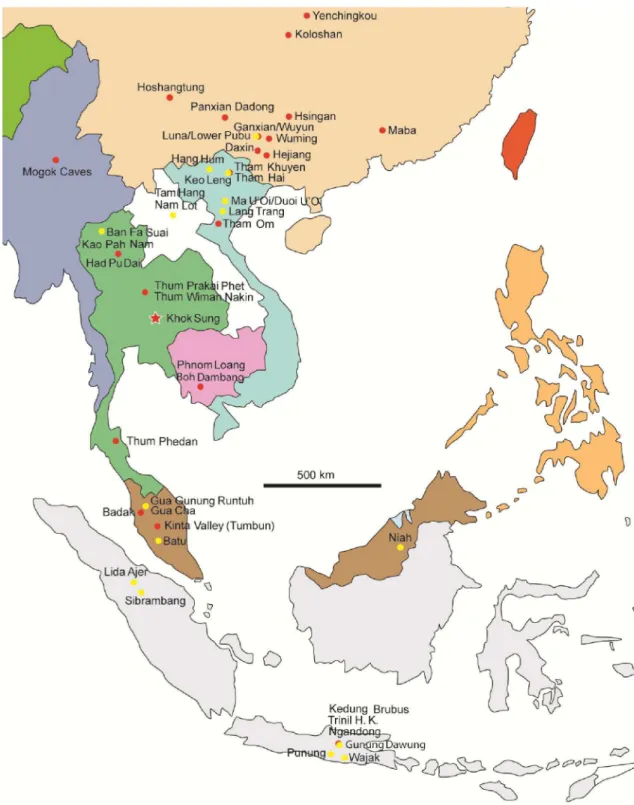
become obtained rapidly and numerous pieces of the puzzle involving the faunal composition, ages, and paleoenvironments are stepwise reconstructed. The latest informations on these aspects are hereby updated for the following countries in mainland Southeast Asia (Fig. I5).
I.8.1) Myanmar
Sites along the Irrawaddy River terraces and localities of Mogok Caves are representatives of the Early to Middle Pleistocene mammalian faunas in the country (Colbert, 1943) (Fig. I5). The ages of these two faunas have been poorly established since the 20th
century excavation, only based on the presence of Stegodon and other archaic taxa (Colbert, 1943), without radiometric dating analyses. The Upper Irrawaddy fauna, being of Early Pleistocene in age (Stamp, 1922; Colbert, 1938), consists of obviously archaic species such as Stegodon insignis, Elephas hysudricus, Rhinoceros sivalensis (later regarded as junior synonym of Rhinoceros unicornis by Antoine (2012)), Equus yunnanensis, and Hexaprotodon iravaticus (Colbert, 1943; Takai et al., 2006). The taxonomic validity of Rhinoceros sivalensis is still controversial (Yan et al., 2014). Only two modern species, Rhinoceros sondaicus and Capricornis cf. sumatraensis, were also present in this assemblage (Colbert, 1938; Zin-Maung-Maung-Thein et al., 2006; Takai et al., 2006). However, the presence of Hipparion cf. antelopinum and Stegodon elephantoides in the Early Pleistocene of the Upper Irrawaddy fauna (Colbert, 1943) is ambiguous. They might have been possibly collected from the older strata (the Lower Irrawaddy Formation, late Miocene to Pliocene in age) according to Takai et al., (2006).
Figure I.5. Map of mainland Southeast Asian countries showing the location of the Early (blue), Middle (red) and Late (yellow) Pleistocene fossil sites.
Although Hooijer (1950) considered the faunas of Mogok Caves as also being of Early Pleistocene age, the extinct proboscideans, Stegodon orientalis, and Palaeoloxodon namadicus, rather suggest a Middle Pleistocene age (Colbert, 1943), in agreement with Louys et al. (2007). However, Takai et al. (2006) argued that the material of Palaeoloxodon, described by Colbert (1943), possesses the similar morphological features of extant Asian elephants and should be assigned to the genus Elephas. Moreover, the presence of a fossil giant panda (Ailuropoda melanoleuca) was probably documented from nearby caves (Colbert, 1943). The paleoenvironments of these two faunas are interpreted to have
corresponded to closed habitats due to the synecological (community-based) methods (Louys and Meijaard, 2010). However, this interpretation is probably a result of the limited number of taxa recovered from the sites. Louys and Meijaard (2010) further suggested that the mixed and open habitats would have been more consistent for these sites because the climatic fluctuations of the Pleistocene resulted in expanding and contracting forests within a more open vegetation matrix. On the other hand, evidence of grasslands was documented during the Early Pleistocene based on the palynological records, indicating a more widespread expansion of savannah vegetation, as well as charred grass cuticles (Morley, 1998).
I.8.2) Laos
The paleoenvironmental information for the Pleistocene of Laos is mostly achieved from the fissure filling deposits of Nam Lot (Bacon et al., 2012) and Tam Hang South (Bacon et al., 2008a, 2011) (Fig. I5), where the ages have been attributed to around the early Late Pleistocene (86-72 ka and 94-60 ka, respectively) on the basis of the luminescence and U-series dating methods combined with the faunal correlations (Bacon et al., 2015). The composition of the Nam Lot and Tam Hang South assemblages consists of modern and extinct taxa, the former site yielding further remains of Homo sp. (Bacon et al., 2011, 2012). The cave of Nam Lot is suggested to have been younger than that of Tam Hang South on the basis of the presence of two modern taxa, Muntiacus muntjak and Sus scrofa, which represent more advanced evolutionary stages (Bacon et al., 2015). The paleoenvironments of these two sites were interpreted as corresponding to mixed habitats with open seasonal deciduous forests and grassy covers based on the presence of numerous cervids, bovids, and suids (Bacon et al., 2015) and on the synecological (community-based) methods (Louys and Meijaard, 2010).
According to the most recent debated topics on the modern human migrations into Asia, modern man might have expanded eastward from Africa and colonized South Asia before 60 ka (after Toba), and have subsequently spread across Southeast Asia around 50 ka, possibly through the past coastline, as exemplified by the stone artifact and genetic evidence (Appenzeller, 2012). This hypothesis confirms that identifications of Homo sapiens remains recovered from the late Middle Pleistocene of Southeast Asian and South Chinese sites are not consistent. Interestingly, a human cranium from the cave of Tam Pa Ling (Fig. I3), where the age has been dated between 63 and 46 ka by OSL and TL dating of sediments and U-series dating of bones, provides the first evidence of early modern humans in mainland
Southeast Asia, supporting an early dispersal out of Africa and into Southeast Asia by the Late Pleistocene (Demeter et al., 2012, 2015). The presence of extinct arvicolines (Rodentia) in this cave might have indicated cooler environments than today (Demeter et al., 2015). I.8.3) Cambodia
A few data documenting Cambodian Pleistocene mammal faunas are known, only some sites having been previously described (e.g., Phnom Loang in Kampot). Beden and Guérin (1973) reported a Middle Pleistocene fauna, known as “Phnom Loang” (Fig. I5), which corresponds to both the modern (e.g., Panthera tigris, Rusa unicolor, Bubalus cf. arnee) and the extinct (Crocuta crocuta ultima, a spotted hyaena) taxa. A Middle Pleistocene age has been attributed to that site (Beden and Guérin, 1973), probably slightly older than that of Thum Wiman Nakin (>169 ka) based on the evolutionary stage of the fauna by Bacon et al. (2011). Louys and Meijaard (2010) performed a discriminant function analysis of the community structure of the mammalian fauna and suggested a mixed habitat for this locality. In addition, thick laterites in Cambodia occurred commonly during the Middle Pleistocene (Takaya, 1967), suggesting some seasonality of the humid tropical climate (Whittow, 1984). Another late Middle Pleistocene cave, known as “Boh Dambang”, in Kampot has been reported by Demeter et al. (2013) (Fig. I5). The mammalian assemblage, as well as the age, is considered to be similar to that of Phnom Loang, consisting of several modern taxa: Prionailurus bengalensis, Cuon alpinus, Ursus thibetanus, Axis porcinus, Rusa unicolor, Muntiacus muntjak, Bos sp., Bubalus arnee, Capricornis sumatraensis, and Macaca sp. and locally extinct species: Crocuta crocuta ultima and Pongo pygmaeus (Demeter et al., 2013). The presence of the spotted hyaena and large bovids in the locality is supposed to have represented a humid climate with relatively open environments (Demeter et al., 2013). I.8.4) Vietnam
Three Middle Pleistocene sites including Tham Khuyen (~475 ka, based on the U-series (performed on speleothems) and ESR (operated with tooth enamel and sediments) analyses, Ciochon et al., 1996), Tham Hai ((~475 ka based on the biochronogical correlation, Olsen and Ciochon, 1990), and Tham Om (250-140 ka based on the biochronological correlation, Olsen and Ciochon, 1990) have been known since the late 20th century
excavation (Fig. I5). Two preceding Vietnamese faunas, Tham Khuyen and Tham Hai, have displayed several extinct taxa (e.g., Gigantopithecus, Pongo, Stegodon orientalis, Megatapirus augustus, and a mystery ape (previously identified as belonging to Homo
erectus, Ciochon (2009)), but modern species (e.g., Sus scrofa, Rusa unicolor, Muntiacus muntjak, and Bos gaurus) were also documented (Kha and Bao, 1967; Cuong, 1971; Olsen and Ciochon, 1990). The Tham Om fauna shows relatively similar taxa with those of Tham Khuyen and Tham Hai, but lacks Gigantopithecus blacki. This species became possibly extinct during the late Middle Pleistocene in Vietnam (Louys et al., 2007). Louys and Meijaard (2010) used a discriminant function analysis of the community guild structure to interpret the paleoenvironments of these three caves. The results suggest that the paleoenvironments corresponded to mixed habitats for all of these localities.
Based on the faunal correlations, Hang Hum and Lang Trang seem to be representatives of the late Middle to early Late Pleistocene sites in Vietnam (Fig. I3), which is confirmed by absolute ages tentatively estimated between 140 and 80 ka (Kha, 1976; Olsen and Ciochon, 1990) and between 100 and 80 ka (de Vos and Long, 1993; Long et al., 1996), respectively. These two faunas exhibit relatively modern elements, especially carnivores, with some extinct taxa (e.g., Pongo pygmaeus, Stegodon orientalis, Palaeoloxodon namadicus, and Dicerorhinus sumatrensis). Archaic Homo sp. and Chinese suids (Sus cf. officinalis and Sus cf. lydekkeri) were additionally recovered from Hang Hum (Kha and Bao, 1967; Cuong, 1985; Olsen and Ciochon, 1990), whereas a giant panda (Ailuropoda melanoleuca) was further present in Lang Trang (de Vos and Long, 1993). The presence of Pongo in Hang Hum and Lang Trang suggests a tropical to sub-tropical vegetation cover, whereas the temperate bamboo species are presumably present according to the occurrence of Ailuropoda melanoleuca in the latter site (Tougard et al., 1996; de Vos, 1983; Louys et al., 2007). The synecological analysis using discriminant function methods indicates also mixed habitats for these two localities (Louys and Meijaard, 2010). For the late Middle to Late Pleistocene site of Ma U’Oi in northern Vietnam (Fig. I5), the in situ mammalian fossils (dated to the Late Pleistocene, >49 ka based on the U/Th dating performed on the fossiliferous braccia) have yielded a relatively modern species, which are still living today in Vietnam, and extinct Palaeoloxodon namadicus (Bacon et al., 2004, 2006). Two teeth and skull fragments assigned to archaic Homo were extracted from the ground floor in rooms A2 and A3 and a corridor A (Demeter et al., 2004, 2005). Numerous remains of microvertebrates (including primates, rodents, insectivores, chiropterans, and small reptiles and amphibians) were also recovered from the roof of corridor “A” in the same cave (Bacon et al., 2006; fig. 4). The in situ Ma U’Oi fauna suggests open woodlands close to the present-day environments in Vietnam, whereas the microvertebrate fauna (dated to the late Middle
Pleistocene, 193 ± 17 ka) indicates a more closed canopy (Bacon et al., 2006). With regards to the Late Pleistocene cave of Duoi U’Oi (66 ± 3 ka for the U/Th geochronology of the calcitic floor) (Fig. I5), the large mammalian fauna is slightly less diversified than those of Tam Hang South and Nam Lot, but a species of the carnivore guild (a leopard, Panthera pardus) is additionally documented (Bacon et al., 2008b). Two isolated teeth assigned to Homo sp. were also recovered from this site (Bacon et al., 2008b).This fauna is suggested to have been typical of those living in open seasonal forests (Bacon et al., 2015). In addition, between 62 and 19 ka, the reworking of aeolian sands along the southeastern Vietnam coast has indicated a reduction of vegetation covers and landscape instability, in relation to the climate change, for this area (Murray-Wallace et al., 2002). The possibly more recent site of Keo Leng (30-20 ka based on the faunal correlation with other Vietnamese localities, Cuong (1985)) (Fig. I5) yields most of modern faunas (including Homo sapiens) with some extinct taxa: Ailuropoda melanoleuca, Pongo pygmaeus, and Stegodon orientalis (Kha, 1976; Long and Du, 1981; Olsen and Ciochon, 1990). The paleoenvironments of this site might have corresponded to a closed habitat due to the presence of both Pongo and Ailuropoda. The Late Pleistocene sites from Vietnam seem to have therefore represented a variety of habitat types (Louys and Meijaard, 2010).
I.8.5) Peninsular Malaysia
The Kinta valley (Tambun) fauna constitutes possibly one of the few Middle Pleistocene sites in the Malay Peninsula (Fig. I5). This fauna has yielded some archaic species including Hexaprotodon sp., Duboisia santeng, and Palaeoloxodon namadicus, but a modern Javan rhinoceros, Rhinoceros sondaicus, was also present (Hooijer, 1962). Other modern species were probably present, but the more detailed taxonomic identification needs to be further addressed. According to the faunal correlations, the age of the site has been tentatively established as being around the Middle Pleistocene (Hooijer, 1962; Medway, 1972). However, the dating of this site is not well-constrained, a Late Pleistocene age being possible according to the geological evidence (Kamaludin et al., 1993; Thorp and Thomas, 1993; Kamaludin and Azmi, 1997; Teeuw et al., 1999). The paleoenvironment of this site is supposed to have corresponded to a savannah condition with some parts of rather swampy areas according to the presence of ecological mammal indicators (Duboisia santeng and an extinct hippopotamus (Hooijer, 1962), in agreement with Heaney (1991) who suggested a savanna corridor for the Sundaland during the Middle Pleistocene. The late Middle Pleistocene site of Badak Cave (dated between 278 and 208 ka, based on OSL and TL datings
of sediments) was described by Ibrahim et al. (2013) (Fig. I5). Its fauna contains obviously modern elements of large mammals (such as Ursus thibetanus, Helarctos malayanus, Sus scrofa, Rusa unicolor, Muntiacus muntjak, and Capricornis sumatraensis) and a locally extinct orangutan (Pongo sp.). Peat with detritus remains of Pinus, Gramineae, and ferns were recorded from the possible Middle Pleistocene of Sunung, an area of about 230 km south of Badak Cave C, indicating the existence of a savanna-type habitat with more seasonal climates (Batchelor, 1979; Morley and Flenley, 1987). Moreover, Morley (1998) mentioned that Pinus savanna was probably widespread on the Malay Peninsula at about 660, 480, 200, 22 ka. During the interglacial period, the climate in the lowlands was probably similar to that of prevailing today, as suggested by the palynological records deposited at about 80 and 55 ka (Kamaludin and Azmi, 1997).
The other younger caves are formed within the metamorphosed Kuala Lumpur limestone and known as “Batu Caves” (Fig. I5), being of Late Pleistocene in age (between 66 and 33 ka) according to the OSL and TL datings of sediments (Ibrahim et al., 2013). These caves have yielded a similar large mammal fauna to that of Badak Cave, but some modern taxa (e.g., Panthera tigris, Tapirus indicus, Dicerorhinus sumatrensis, and Sus cf. barbatus) were further documented. The presence of Pongo suggests a prevailing (evergreen) forest habitat for both Badak Cave and Batu Caves, implying that sufficient forest covers persisted in the western coast of Peninsular Malaysia through the late Middle to middle Late Pleistocene (Ibrahim et al., 2013). Other late Late Pleistocene to Holocene sites, where large mammal remains have been recovered in Peninsular Malaysia, were reported (e.g., Gua Gunung Runtuh (Davidson, 1994) and Gua Cha (Groves, 1985)) (Fig. I5). In the Quaternary deposits of Sungei Besi, western Malaysia, carbon-dated peat and wood and pollens suggest perhumid vegetation that occurred at around 41.2 and 36.4 ka (Ayob, 1970). Overall, the closed environements (evergreen vegetation) were dominating and existed in Peninsular Malaysia before the Late Glacial Maximum (LGM).
I.8.6) Thailand
In Thailand, large mammalian fossils have been never or poorly recorded from the Early Pleistocene, but having been better documented during the Middle to Late Pleistocene. The occurrence of Pliocene to Early Pleistocene Formation of thick laterites in the lower Central Plain of Thailand (Thiramongkol, 1986) could suggest high seasonality under humid tropical climate (Whittow, 1984). Several Middle fossiliferous localities include Had Pu Dai
(Pramankij and Subhavan, 2001a), Kao Pah Nam (Pope et al., 1981), Thum Wiman Nakin (Ginsburg et al., 1982; Chaimanee and Jaeger, 1993; Tougard, 1998, 2001), Thum Prakai Phet (Tougard, 1998; Filoux et al., 2015), Thum Phedan (Yamee and Chaimanee, 2005), and the Cave of the Monk (Zeitoun et al., 2005, 2010), all of which are fissure-filling deposits (Fig. I5). The Had Pu Dai fauna has not been studied in details yet, but fundamentally consists of a giant panda, a hyaena, cervids, suids, and possibly a large extinct ape (Gigantopithecus sp.) and an orangutan (Pongo sp.) (Tobias, 2002). An age of 500 ka has been proposed by Pramankij and Subhavan (2001a; 2001b) on the basis of the presence of archaic taxa. However, the age of Had Pu Dai is still doubtful due to the inadequate taxonomic descriptions of the fauna. The Kao Pah Nam site yields modern (e.g., Panolia eldii and Bos cf. gaurus) and extinct (Crocuta sp., Hippotamus?, and Pongo?) taxa. The age of the site has been dated to around 690 ka based on the geological and faunistic data by Pope et al. (1981). The paleoenvironments of this site have been interpreted as corresponding to relatively open and dry Dipterocarp woodland based on the presence of Hippopotamus, hyaenids, and large cervids and bovids as well as the absence of gibbons (Pope et al., 1981). During the late Middle Pleistocene, Thum Wiman Nakin represents one of the most abundant and diversified mammal fossils in mainland Southeast Asia. It contains numerous modern (e.g., Ursus thibetanus, Rhinoceros sondaicus, Sus scrofa, Axis porcinus, Rusa unicolor, Bos javanicus, Bubalus arnee, Capricornis sumatraensis) and extinct (e.g., Crocuta crocuta ultima, Ailuropoda melanoleuca, Pongo pygmaeus, and Rhinoceros unicornis) taxa that are representatives of the late Middle Pleistocene fauna (Tougard, 1998, 2001), dated to 169 ka for the minimum age based on the U-series dating of the stalagmitic floor above the fossiliferous layer (Esposito et al., 1998, 2002). A tooth of Homo sp. was also recovered in this locality (Tougard et al., 1998). Chaimanee (1998) suggested a wetter and cooler climate than today on the basis of the rodent assemblage and vegetation. Based on an analysis of the cenogram (Legendre, 1986, 1989), the paleoenvironments of Thum Wiman Nakin are characterized by a slightly open forest landscape with relatively humid conditions (Tougard and Montuire, 2006). According to stable carbon isotope analysis of tooth enamel of Thum Wiman Nakin mammals, the paleovegetation was dominated by a mosaic of C3 and C4 plants and the paleoenvironments corresponded to open and semi-wooded savannah with forest patches and deep forest cover (Pushkina et al., 2010). The cave of Thum Prakai Phet is suggested to be similar in faunal composition and age to that of Thum Wiman Nakin (Tougard, 1998; Filoux et al., 2015). Accordingly, the paleoenvironments of Thum Prakai Phet are proposed to have corresponded to a slightly open forest (Tougard, 1998; Filoux et