DOCTEUR DE L'UNIVERSITÉ DE POITIERS UFR des sciences fondamentales et appliquées
Laboratoire de neurosciences expérimentales et cliniques - LNEC (Diplôme National - Arrêté du 7 août 2006)
École doctorale : Biologie-santé - Bio-santé
Secteur de recherche : Aspects moléculaires et cellulaires de la biologie
Présentée par : Joëlle Nader
Nouvelles stratégies pour prévenir les effets néfastes des psychostimulants : l'exposition à l'environnement enrichi
et la stimulation du système cannabinoïde endogène
Directeur(s) de Thèse : Marcello Solinas, Nathalie Thiriet Soutenue le 16 novembre 2012 devant le jury
Jury :
Président Mohamed Jaber Professeur des Universités, Université de Poitiers Rapporteur Catherine Belzung Professeur des Universités, Université de Tours
Rapporteur Catherine Le Moine Directeur de recherche, CNRS, Université Victor Segalen Bordeaux 2 Membre Marcello Solinas Chargé de recherche, CNRS, Université de Poitiers
Membre Nathalie Thiriet Maître de conférences, Université de Poitiers Membre Emmanuel Valjent Chargé de recherche, INSERM de Montpellier
Pour citer cette thèse :
Joëlle Nader. Nouvelles stratégies pour prévenir les effets néfastes des psychostimulants : l'exposition à
l'environnement enrichi et la stimulation du système cannabinoïde endogène [En ligne]. Thèse Aspects moléculaires et cellulaires de la biologie. Poitiers : Université de Poitiers, 2012. Disponible sur Internet
1
Pou l’o te tio du G ade de
DOCTEUR DE l’UNIVER“ITE DE POITIER“
(Faculté des Sciences Fondamentales et Appliquées)
(Diplôme National
– Arrêté du 7 Août 2006)
Ecole Doctorale : BioSanté N°524
Secteur de Recherche : Aspects moléculaires et cellulaires de la
Biologie
Présentée par
Joëlle NADER
Nouvelles stratégies pour prévenir les effets néfastes des
psychostimulants
: l’e positio à l’e vi o e e t e i hi et
la stimulation du système cannabinoïde endogène
Directeurs de Thèse
Nathalie THIRIET et Marcello SOLINAS
Soutenue le 16 Novembre 2012
de a t la Co
issio d’E a e :
Madame le Professeur Catherine BELZUNG
Rapporteur
Madame le Professeur Catherine LE MOINE
Rapporteur
Monsieur le Docteur Emmanuel VALJENT
Examinateur
Monsieur le Professeur Mohamed JABER
Examinateur
Madame le Docteur Nathalie THIRIET
Examinateur
2
A mes parents … Et A Dany …
3
remercie Madame le Professeur Catherine BELZUNG et Madame le Docteur Catherine LEMOINE d’avoir accepté d’être les rapporteurs de cette thèse.
Merci également à Monsieur le Docteur Emmanuel VALJENT, pour
m’avoir fait l’honneur de faire partie de mon jury.
Je tiens à remercier Monsieur le Professeur Mohamed Jaber qui m’a accueillie dans son unité. Merci pour m’avoir permis de vivre cette expérience dans les meilleures conditions, humaines et scientifiques, et de m’avoir permis de faire partie de l’agréable famille du « LNEC ». Je ne pouvais rêver d’un milieu plus enrichissant pour faire mes premiers pas dans le monde de la recherche !
Je tiens à exprimer ma sincère gratitude à Madame le Docteur Nathalie THIRIET, ma directrice de thèse, qui a dirigé l’ensemble de ce travail. Je te suis très reconnaissante, pour avoir cru en mes capacités et pour la confiance que tu m’as accordée dès mon premier jour au labo. Merci pour ton encadrement et tes compétences scientifiques. Merci pour ton implication, ta générosité, ta disponibilité et ta patience. Merci pour tes conseils, toujours pertinents, et ton amitié. J’ai énormément appris à travers nos conversations scientifiques et variées. Ces années auront été très enrichissantes, aussi bien sur le plan scientifique que sur le plan humain.
Je remercie également Monsieur le Docteur Marcello SOLINAS qui a codirigé cette thèse. Merci pour la richesse de tes idées, ta rigueur scientifique, ton enthousiasme pour la Science et ta disponibilité. Merci de m’avoir donné la possibilité de participer à plusieurs congrès, nationaux et
internationaux, et de découvrir le monde de la recherche.
Un très grand merci à tous les membres du LNEC, chercheurs, enseignants chercheurs, techniciens et personnel administratif, qui m’ont très bien accueillie. Merci pour votre gentillesse, votre disponibilité et votre bonne humeur.
4
souris pendant ces trois ans.
A toutes les « filles » du labo. Pour les citer : Sophie, Céline, Claire, Claudia, Marine, Maureen, Virginie, Laetitia et Nathalie V.. Merci pour les pauses déjeuners, les « restals », les goûters, et toutes les discussions et les fous rires que nous avons eus ensemble. Ces moments vont certainement me manquer.
Merci Maureen pour toutes les heures de défoulement en danse africaine et au volley (aussi bien sur le terrain que dans les gradins !).
Merci à tous mes amis, libanais et français, tourangeaux, parisiens et pictaviens, qui ont rendu inoubliables ces huit ans passés en France.
A Pascale M. « Marha », il y a un an, tu étais à ma place ! Tu m’as dit « à chacun son tour ». Aujourd’hui c’est à moi de te remercier pour toutes les sorties shopping, les cafés, les discussions, les pleurs, les délires, les chants, les conseils... Merci pour les inoubliables semaines passées à Rome, à Barcelone, et à Poitiers. Les aventures de « Farha w Marha » ne s’arrêteront pas là!!
A Hania A. Merci pour toutes ces journées et nuits passées à rédiger, à discuter, et à rigoler. J’ai fini !! Maintenant, c’est à ton tour de clôturer cette expérience et de publier ton travail … Tiens bon, tu y es presque !!
Un grand merci à ma deuxième famille, les SABEK. Douha, merci pour tous les conseils que tu m’as donnés, pour les WE remplis de conversations, de rires et de confidences. Merci de m’avoir donné la chance de faire partie de ta merveilleuse famille, et de m’avoir confié tes adorables
enfants, Roxane, Phillipe et Louise.
Toute ma reconnaissance va à la famille Chamieh, parents et enfants. Merci pour tous les moments passés en famille. Une pensée particulière à Michel C., pour tous les bons moments que nous avons passés ensemble, aussi
5
Un grand merci à Houssam H. Les quelques phrases que je me permets de t’adresser ici ne sont pas suffisantes pour exprimer tout le respect, l’amitié et la reconnaissance que j’éprouve envers toi. Merci d’avoir été mon confident, de m’avoir supportée et soutenue, pendant toutes ces années. On a tous besoin d’une personne sur qui compter. Merci d’avoir été cette personne. Merci d’avoir répondu présent aux moments où j’en avais le plus besoin. Merci pour nos longs débats et nos chamailleries. Merci pour tes supers recettes et les supers restos que nous avons découvert ensemble, à Tours, à
Poitiers, à Besançon, à Paris, à Langres, à Dijon, à Bordeaux…
Enfin, à ma famille adorée, sans qui je ne serai pas là aujourd’hui. Papa, mon ange gardien, ta présence m’a manqué pendant ces années et me manquera toujours. Qu’est ce que je n’aurai donné pour t’avoir à mes côtés en ce grand jour. Merci de m’avoir appris à aimer la vie. J’espère que, depuis le paradis, tu es fier de ce que je suis devenue et de tout ce que j’ai accompli jusqu’à présent.
Maman, je t’adresse ici toute mon affection et ma plus profonde estime. Merci pour tout l’amour et la confiance que tu m’as témoigné, malgré les milliers de kilomètres qui nous séparent. Merci d’avoir été assez forte pour me soutenir et supporter mes sauts d’humeur. Merci pour ton « c’est pas grave !» qui m’a souvent aidé à franchir des obstacles de la vie.
A Joseph, mon grand frère sur qui j’ai pu compter pendant toutes ces années. Merci pour tes conseils, tes encouragements et ton soutien dans toutes les décisions que j’ai prises.
A mes sœurs, Jessy et Joyce. Pour tous nos souvenirs d’enfance que je garde au fond du cœur. Pour toutes les confidences auxquelles j’ai eu droit pendant les vacances d’été. Je suis désolée d’avoir choisi de grandir loin de vous. Mais je me rattraperai bientôt, c’est promis.
A mon petit frère, Joudy, que j’ai vu grandir par épisodes de quelques semaines. J’aurai voulu être présente plus souvent à tes côtés. J’espère que tu es fier de ce que ta grande sœur a pu accomplir, parce que moi, je suis très
6
Huit ans en France, dont trois à Poitiers. Une expérience durant laquelle j’ai eu la chance de croiser un grand nombre de personnes enthousiastes, optimistes et sympathiques.
Huit ans de bonheur, de rencontres, et de découvertes, personnelles et professionnelles.
Huit ans qui ont fait de moi la personne que je suis aujourd’hui.
Huit ans au bout desquels j’ai rencontré mon âme sœur, l’homme avec qui j’écrirai la nouvelle histoire de ma vie, mon Dany !!
« Toute histoire a une fin. Mais dans la vie, chaque fin annonce une
7
RESUME ... 11
SUMMARY ... 12
Liste des travaux ... 13
Liste des illustrations ... 14
Liste des abréviations... 16
Structure générale du manuscrit ... 18
Revue de la littérature ... 19
Chapitre 1 - Les psychostimulants ... 20
A- Généralités ... 20
1. La Cocaïne ... 20
2. La Méthamphétamine ... 22
B-Effets psychologiques et somatiques des psychostimulants... 24
1.Effets psychologiques ... 24 i.Administration aigue ... 24 ii.Administration chronique ... 24 2.Effets somatiques ... 24 C- Effets neurobiologiques ... 25 1.Cibles moléculaires ... 25
i.Mode d’a tio de la o aï e ... 25
ii.Mode d’a tio de la METH ... 26
2.Circuits neuronaux impliqués dans les effets récompensants des psychostimulants ... 27
i.Description du système dopaminergique mésocorticolimbique... 28
ii.Des iptio du i uit de l’a gdale te due ... 29
iii.Interactions des psychostimulants avec le circuit de la récompense ... 30
3.Interactions des psychostimulants avec les systèmes neuronaux impliqués dans la réponse au stress... 31
i.L’a e HPA : principal système physiologique de réponse au stress... 31
ii.Le système du CRF : un neuropeptide impliqué dans la réponse au stress ... 34
D- Dépendance aux psychostimulants ... 35
1.Description de la dépendance ... 35
i.Diagnostic ... 35
ii.Vulnérabilité ... 36
2. Mécanismes impliqués dans la dépendance ... 38
i.Les récepteurs dopaminergiques ... 40
ii.Les kinases ... 40
iii.Les facteurs de transcription : exemple de CREB ... 43
iv.Les gènes à réponse précoce ... 46
E-Toxicité induite par la METH ... 47
1.Atteintes neurologiques induites par la METH ... 47
i.Attei tes eu ologi ues hez l’Ho e ... 48
ii.Atteintes neurologiques chez les primates non humains ... 49
iii.Atteintes neurologiques chez les rongeurs ... 50
2.Mécanismes de toxicité induits par la METH ... 50
i.METH et stress oxydatif ... 51
ii.METH et Hyperthermie ... 57
8
3.Exposition à divers stress et consommation de drogues ... 62
i.Le stress ... 62
ii.Interactions neurobiologiques entre facteurs de stress et addiction ... 66
4.Impact des expériences positives sur la dépendance et mod le de l’e i o e e t enrichi ... 68
i.L’e i o e e t e i hi ... 68
ii.I pa t de l’EE da s di e s od les de d pe da e au d ogues ... 71
F-Impact environnemental sur les processus neurotoxiques ... 74
1.Impact des facteurs de stress sur les processus neurotoxiques ... 74
i.Effets toxiques du stress dans les régions corticolimbiques ... 74
ii.Impact du Stress sur les processus neurotoxiques ... 75
.I pa t de l’E i o e e t E i hi su les p o essus eu otoxiques ... 77
i.I pa t de l’EE su les aladies eu od g ati es ... 77
ii.I pa t de l’EE su les p o essus eu oto i ues i duits pa les ps hosti ula ts ... 78
Chapitre 3
– Le Système Cannabinoïde Endogène ... 79
A-Généralités ... 80
G-Les composants du système cannabinoïde endogène ... 80
1.Les Récepteurs Cannabinoïdes ... 80
i.Le récepteur CB1 ... 80
ii.Le récepteur CB2 ... 81
iii.Signalisation des récepteurs CB1 et CB2 ... 82
iv.Autres récepteurs fixant les cannabinoïdes ... 83
2.Les Endocannabinoïdes ... 84
i.L’A a da ide ... 85
ii.Le 2-arachidonoylglycérol ... 86
iii.Autres ECs ... 88
iv.Les ECs sont synthétisés à la demande ... 89
v.Les ECs : des messagers rétrogrades... 89
vi.La métabolisation des ECs ... 90
H-SCE et système de récompense ... 92
1.Interaction du SCE avec le système de récompense ... 92
i.Implication des récepteurs CB1 ... 92
ii.Implication des récepteurs CB2 ... 94
iii.Effets des ECs sur le système de récompense ... 95
.Effets des d ogues d’a us su le “CE ... 96
I-SCE et systèmes de stress ... 97
1.Régulation des émotions par le SCE ... 97
i.Les récepteurs cannabinoïdes ... 97
ii.Les ECs ... 98
2. Régulation du SCE par le stress ... 99
J-Rôle neuroprotecteur du SCE ... 99
1.SCE et atteintes cérébrales ... 100
2.Mécanismes potentiellement impliqués dans la neuroprotection induite par le SCE ... 101
i.SCE et modulation des neurotransmissions ... 101
ii.SCE et modulation du système immunitaire ... 103
iii.Effets opposés sur la survie/mort neuronale ... 104
9
4.La modulation du niveau de stress : un mécanisme sous-te da t l’i pa t des
manipulations environnementales sur la dépendance aux drogues ? ... 163
i.L’e positio à u EE di i ue le i eau d’a i t ... 164
ii.Le « switch » environnemental induit un état émotionnel négatif ... 164
iii.Mécanismes régulés par les manipulations environnementales et potentiellement impliqués dans la modulation du niveau de stress des individus ... 165
5.Perspectives ... 169
i.Etude de l’i pli atio de la eu ot a s issio dopa i e gi ue da s les effets p e tifs de l’EE su la se si ilisatio à la o aï e ... 169
ii.Existe-t-il une ségrégation neuronale de la phosphorylation de CREB induite par le « switch » EE>SE ? ... 172
K-Stratégies pour prévenir la neurotoxicité de la METH ... 173
1.Rappel : L’e positio à l’EE e duit pas la toxicité de la METH ... 174
2.La stimulation pharmacologique du SCE : une nouvelle approche pour prévenir la toxicité induite par la METH ... 174
3.Perspectives : caractériser les mécanismes sous tendant la neuroprotection induite par le SCE ... 176
i.Quel récepteur est impliqué dans la protection induite par le JZL184 ? ... 176
ii.R gulatio de l’a ti it i ogliale ... 176
Conclusion ... 178
10
psychostimulants a montré que des facteurs négatifs, comme le stress, augmentent le risque de d eloppe u e addi tio , alo s ue des fa teu s positifs, o e l’e positio à des conditions stimulantes, le réduisent. Une partie de cette thèse a consisté à rechercher les mécanismes neurobiologiques et cellulaires qui sous tendent cette influence e i o e e tale. Ai si, l’e positio d’a i au à u e i o e e t e i hi EE , ui procure des conditions stimulantes, diminue leur iveau d’a i t , un effet qui serait en partie lié à la régulation de gènes appartenant au système cannabinoïde endogène (SCE) dans des régions impliquées dans la réactivité au stress (Article 1). Par ailleurs, nos travaux ont mis en évidence des limites de l’e positio à l’EE : quand celle-ci est interrompue, ses effets bénéfiques sont perdus et la vulnérabilité à la cocaïne est même augmentée. Ceci s’e pli ue ait pa l’appa itio d’u tat otio el gatif, asso i à une activation du facteur CREB da s l’a gdale te due, u e gio a efou e t e la o pe se et le stress (Article 2). Nous nous sommes également intéressés à la toxicité de la méthamphétamine (METH) et à sa modulation par le SCE, pour lequel des propriétés neuroprotectives avaient déjà été suggérées. Ainsi, une stimulation pharmacologique du SCE permet de prévenir la neurotoxicité dopaminergique induite par la METH (Article 3). Nos sultats soulig e t la o ple it d’utilisatio des a ipulatio s e i o e e tales et mettent en lumière les capacités protectrices du SCE contre la dépendance et la neurotoxicité engendrées par les psychostimulants.
Mots clés : addiction ; psychostimulants ; environnement enrichi ; stress ; système cannabinoïde endogène ; neurotoxicité
11
have shown that negative factors, such as stress, increase the risk of developing drug addiction, while positive factors, such as exposure to stimulating conditions, reduce it. The first aim of this work was to look for the neurobiological and cellular mechanisms that underlie this environmental influence. We found that exposure of animals to stimulating enriched environments (EE) reduces anxiety levels, an effect that may be partly related to the regulation of genes belonging to the endogenous cannabinoid system (ECS) in regions involved in stress reactivity (Article 1). In addition, our work has highlighted some limitations of the exposure to EE since discontinuation of enrichment results not only in the loss of its beneficial effects but also in increased vulnerability to cocaine. This effect is associated with emotional distress and changes in the activity of the transcription factor CREB in the extended amygdala, an interface region between reward and stress processes (Article 2). We also investigated whether ECS, for which neuroprotective properties have already been suggested, could reduce the brain toxicity induced by methamphetamine (METH). We found that pharmacological stimulation of ECS provides protection against the METH-induced dopaminergic neurotoxicity (Article 3). Our results highlight the complex consequences of environmental conditions on brain and behavior, and highlight the protective role of ECS against both addiction and neurotoxicity induced by psychostimulants.
Keywords: addiction; psychostimulants; enriched environment; stress; endogenous cannabinoid system; neurotoxicity.
12
Publications
Nader J, Chauvet C, El Rawas R, Favot L, Jaber M, Thiriet N, Solinas M. Loss of environmental enrichment increases vulnerability to cocaine addcition. Neuropsychopharmacology. Neuropsychopharmacology (2012) 37:1579-87
El Rawas R, Thiriet N, Nader J, Lardeux V, Jaber M, Solinas M. Early exposure to environmental enrichment alters the expression of genes of the endocannabinoid system. Brain Research (2011) 1390:80-9
Nader J, Rapino C, Gennequin B, François A, Francheteau M, Makryannis A, Macarrone M, Solinas M, Thiriet N. Stimulation of the Endocannabinoid System prevents Methamphetamine-induced neurotoxicity in mice (en cours de préparation)
Communications
L’auteu soulig est elui ui a p se t le t availThiriet N, El Rawas R, Chauvet C, Nader J, Jaber M, Solinas M. Switch from enriched to standard living environment increases the rewarding effects of cocaine. 7th FENS meeting, Amsterdam – Pays Bas, Juillet 2010.
Nader J, Thiriet N, El Rawas R, Chauvet C, Jaber M, Solinas M. Switch from enriched to standard living environment increases the rewarding effects of cocaine. EBPS Workshop
D ug, Ps hiat i Diso de s a d Neu oge esis , Tou s – France, Septembre 2010.
Nader J, Chauvet C, El Rawas R, Favot L, Jaber M, Thiriet N, Solinas M. Switch from positive to neutral living environments increases vulnerability to cocaine. 10th Colloque de la Société des Neurosciences, Marseille – France, Mai 2011.
Thiriet N, Nader J, El Rawas R, Chauvet C, Jaber M, Solinas M. Switch from enriched to standard living environment increases the rewarding effects of cocaine. 8th IBRO world congress, Florence – Italie, Juillet 2011.
Nader J, Gennequin B, François A, Francheteau M, Solinas M, Thiriet N. Stimulation of the Endocannabinoid System prevents Methamphetamine-induced neurotoxicity in mice. 8th FENS meeting, Barcelone – Espagne, Juillet 2012.
13
Figure 1. Structure du chlorhydrate de cocaïne. ... 21
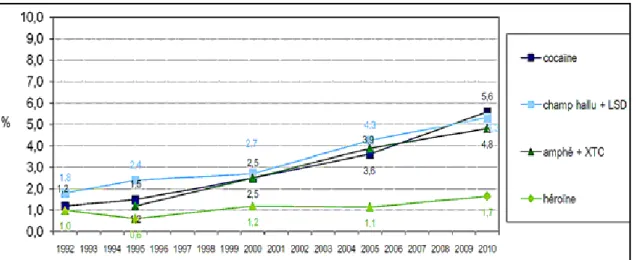
Figure 2. Usage au cours de la vie de substances psychoactives (hors alcool, tabac et cannabis) parmi les 18-44 ans. Evolution en France, entre 1992 et 2010. Amphé : amphétamines ; XTC : extasy. Source : Baromètre Santé 1992, 1995, 2000, 2005, 2010, INPES exploitation OFDT www.ofdt.fr ... 22
Figure 3.Structure de la méthamphétamine. ... 23
Figure 4. Schéma représentatif des circuits neuronaux impliqués dans les effets renforçants des psychostimulants chez les rongeurs. ... 27
Figure 5. Schéma simplifié de la voie mésocorticolimbique chez les rongeurs. ... 29
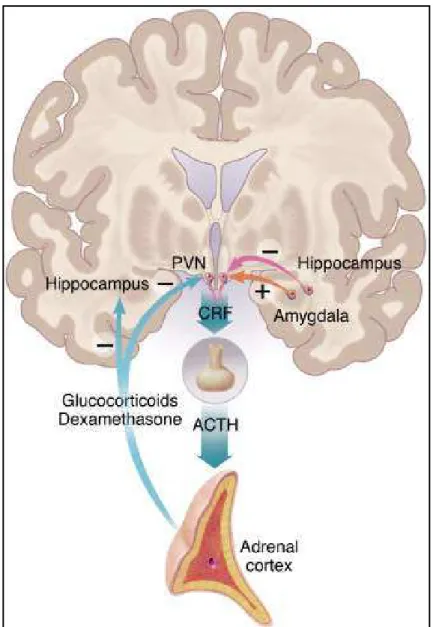
Figu e . L’a e h pothala o-hypophyso-surrénalien (axe HPA) chez l’Ho e. ... 33
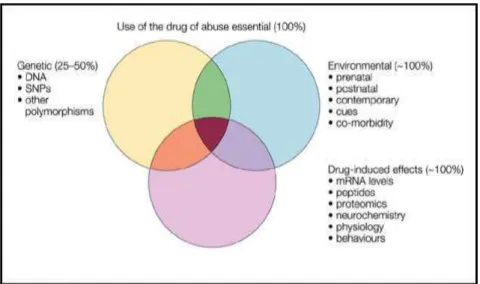
Figu e . L’i te a tio e t e les fa teu s g ti ues, e i o e e tau et asso i s à la d ogue pe et d’ alue la ul a ilit au d eloppe e t de l’addi tio . ... 38
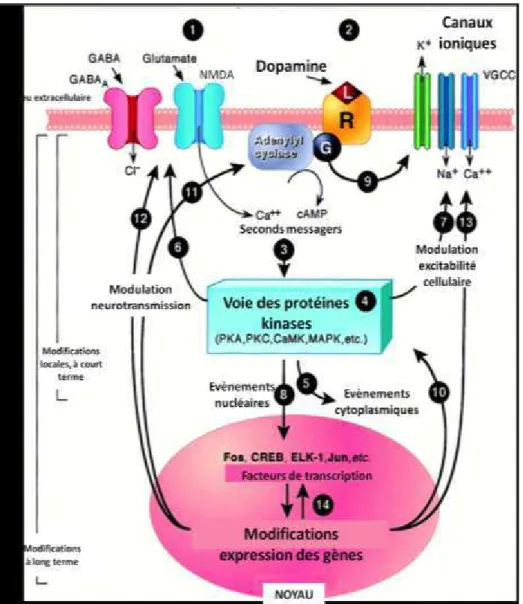
Figure 8. Boucle striato-thalamocorticale impliquée dans la régulation du système glutamatergique. 39 Figure 9. Mécanismes moléculaires intervenants dans les neuroadaptations induites par les drogues d’a us. ... 42
Figure 10. Représentation de la régulation de la stimulation de ERK par les kinases et les phosphatases suite à une administration aigue de psychostimulants. PP1 : protéine phosphatase 1 ; STEP : st iatal e i hed phosphatase d’ap s Valje t et al., ... 43
Figu e . R t o o t ôle de l’a ti it des eu o es dopa i e gi ues pa la d o phi e. ... 45
Figure 12. Régulation des protéines de la fa ille FO“ pa les d ogues d’a us. ... 47
Figu e . M a is es d’i te a tio dopa i e-glutamate impliqués dans la dégénérescence du système dopaminergique nigrostriatale. (Cadet et Brannock 1998) ... 52
Figure 14. Schéma représentatif des causes et conséquences de la neurotoxicité induite par la METH. 54 Figu e . L’a ti atio i ogliale i duite pa la METH. ... 61
Figu e . Rep se tatio s h ati ue de l’i te a tio eu o iologi ue e t e st ess et p ise de drogues. ... 67
Figu e . Illust atio s de l’e i o e ent enrichi... 69
Figure 18. Interactions entre METH et stress. ... 77
14
Figure 21. Deux principales voies de synthèse du 2-AG ont été identifiées (Piomelli, 2003) ... 87 Figure 22. Mécanismes impliqués dans la synthèse du 2-AG ia la PLC. d’ap s Dohe t et
Dingledine, 2003) ... 87 Figure 23.Structure chimique de la Virodhamine et la Noladine. (Piomelli, 2003) ... 88 Figure 24. Les ECs sont des messagers rétrogrades. (Venance et al, 2004) ... 90 Figu e . Lo alisatio des a is es de s th se et de d g adatio des ECs. d’ap s Di Ma zo et al., 2004)) ... 92 Figu e . R gulatio de l’a ti it des a au io iques présynaptiques par activation des récepteurs CB1. (Piomelli, 2003) ... 102 Figure 27.Schéma représentant le rétrocontrôle négatif exercé par les endocannabinoïdes (eCB) sur les eu o es CRF du PVN. “uite à l’a ti atio de l’a e HPA, la o ti ost o e CORT) libérée par les glandes surrénales induit, par activation des récepteurs aux glucocorticoïdes (mGR) (1), la synthèse
et la li atio d’eCB pa les eu o es CRF. Ces eCB a ti e t les epteu s CB e p i s pa les neurones présynaptiques glutamatergiques (4), inhibent la libération de glutamate (5), diminuant ai si la li atio de CRF pa les eu o es du PVN . d’ap s Hill et al, ... 167 Figure 28.Schéma représentant les mécanismes potentiellement impliqués dans la mise en place des conséquences négatives induites par la perte des conditions stimulantes. ... 169 Figure 29. Schéma représentatif des voies de signalisation intracellulaires activées en réponse aux d ogues d’a us. d’ap s Lu et al, ... 171
15
METH : méthamphétamine AMPH : amphétamine DA : dopamine
CPF : cortex préfrontal CPu : noyau caudé putamen NAc : noyau accumbens ATV : aire tegmentale ventrale
BLA : amygdale basolatérale (basolateral amygdala)
DAT : transporteur dopaminergique (dopamine transporter)
VMAT : transporteur vésiculaire de monoamine (vesicular monoamine transporter) GABA : acide gamma-aminobutyrique (gamma aminobutyric acid)
BNST : noyau du lit de la strie terminale (bed nucleus of the stria terminalis) CeA : amygdale centrale (central amygdala)
Axe HPA : axe hypothalamo-hypophyso-surrénalien (hypothalamic-pituitary-adrenal axis) CRF : corticotropin releasing factor
6-OHDA : 6-hydroxydopamine
PVN : noyau paraventriculaire (paraventricular nucleus) ACTH : hormone corticotrope (adrenocorticotropic hormone) SN : substance noire
AMPA : récepteur α-amino-3-hydroxy-5-methyl-4-isoxazolepropionate NMDA : récepteur N-methyl-D-aspartate
AMPc : adénosine monophosphate cyclique
CREB : protéine se fixant au site CRE (cAMP response element binding protein) PKA : protéine kinase A
DARPP32 : phosphoprotéine r gul e pa la dopa i e et l’AMP dopa i e a d AMP responsive phosphoprotein 32)
ERK : extracellular signal-regulated kinase
MAP kinase : protéine kinase activée par des agents mitogènes (mitogen activated protein kinase)
16
SNC : système nerveux central TH : tyrosine hydroxylase
CPP : préférence de place conditionnée (Conditioned Place Preference)
BDNF : facteur neurotrophique dérivé du cerveau (brain derived neurotrophic factor) ROS : espèce réactive oxygénée (reactive oxygen species)
CuZnSOD : cuivre/zinc superoxyde dismutase NO : o o de d’azote it i o de
NOS : NO synthase
ATP : adenosine triphosphate THC: delta-9-hydrocannabinol
SCE : système cannabinoïde endogène
EC : endocannabinoïde ou cannabinoïde endogène JNK : c-Jun-N-terminal kinase
2-AG : 2-arachidonylglycérol PLD : phospholipase D PLA1 : phospholipase A-1 PLC : phospholipase C
FAAH : fatty acid amide hydrolase MGL : monoacylglycerol lipase EE : environnement enrichi SE : environnement standard
MPTP : 1-méthyl-4-phényl-1,2,3,6-tétrahydropyridine SOD : superoxyde dismutase
17
Ce travail de thèse a consisté à rechercher des outils de protection contre les effets addictifs et toxiques des psychostimulants. Dans la première partie de ce manuscrit, une analyse bibliographique décrit le contexte scientifique de ces travaux de thèse. Cette partie est divisée en trois chapitres. Dans le premier, je présenterai les deux psychostimulants utilisés lors de mes travaux de thèse, la cocaïne et la métha ph ta i e. J’e pli ue ai leu s a is es d’a tio et je d i ai, plus sp ifi ue e t, des a is es sous-tendant la neurotoxicité induite par la méthamphétamine qui, en comparaison avec les autres ps hosti ula ts, a la pa ti ula it d’i dui e u e fo te toxicité neuronale. Le deuxième chapitre traitera des facteurs environnementaux supposés avoir un impact sur les effets à long terme induits par les psychostimulants, comme le démontrent les résultats récemment obtenus dans notre équipe. Enfin, le troisième chapitre sera consacré à la présentation du système cannabinoïde endogène que nous avons ciblé, en vue de réduire la toxicité de la méthamphétamine. Dans la deuxième partie du manuscrit seront présentés les principaux résultats obtenus et qui ont fait l’o jet de deu a ti les s ie tifi ues pu li s et d’u troisième, en cours de préparation. Une discussion générale des résultats obtenus et les perspectives envisagées sont présentées à la fin de ce document.
18
19
Chapitre 1 - Les psychostimulants
Dans ce chapitre, je présenterai (1) les deux psychostimulants, la cocaïne et la méthamphétamine (METH), utilisés pendant ma thèse ; (2) les adaptations cellulaires et moléculaires induites par ces deux psychostimulants, ainsi que (3) les mécanismes de neurotoxicité induits dans le cadre de la consommation de METH.
A-
Généralités
1.
La Cocaïne
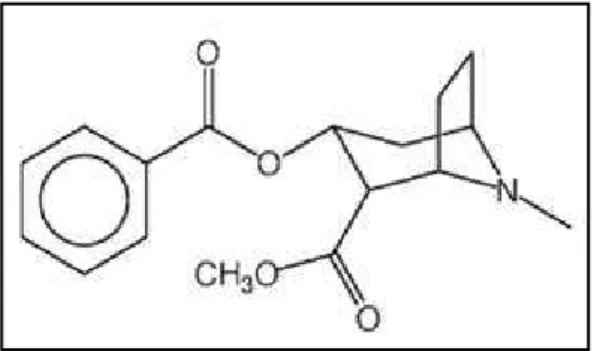
La cocaïne est un alcaloïde extrait à partir des feuilles de deux cocaïers : Erythroxylum coca et Erythroxylum novogranatense. Ces a ustes pousse t à l’ tat sau age e A ique du “ud, da s la o dill e des A des. L’usage de ette pla te e o te à plus de a s, où les peuples A i die s la o sid aie t o e pla te sa e et l’utilisaie t lo s de cérémonies religieuses. Pendant longtemps, la coca fut utilisée, sous forme de tisane ou de pâte masticatoire, pour bloquer la faim et la fatigue, et pour se protéger contre le froid en altitude. Les p e i es feuilles de o a ’a i e t e Eu ope u’e , g â e au ota iste f a çais Joseph de Jussieu, ais l’i t t s ientifique pour cette plante ne débuta u’au XIXèmesi le a e l’e t a tio de la o aï e et la d te i atio de sa fo ule hi i ue
C17H21NO4 (Figure 1). Elle fut notamment utilisée comme anesthésique locale lors des
chirurgies, comme substitut pour traite les addi tio s à l’al ool, à l’opiu et à la o phi e, et dans le traitement des maladies respiratoires et des troubles gastriques. Vers la fin du XIXème siècle la cocaïne fut incorporée dans différentes boissons telles que le Coca-Cola. A la même époque se répandit un nouveau procédé de consommation, plus simple et beaucoup plus efficace : l’i halatio de la poud e de o aï e. Et au d ut du XXème siècle, plusieurs cas de dépendance furent décrits. Commença alors la lutte contre la consommation de la co aï e. L’adoptio du « Harrison Narcotic Act » aux Etats-Unis en 1914 a permis de gle e te l’usage et la dist i utio de la o aï e et à i te di e, p og essi e e t, so usage non-médical. En 1916, la France publia une liste de stupéfiants et une loi interdisant la o so atio des d ogues. E , l’ONU sig a la « convention unique sur les
20
stupéfiants » visant à limiter la production et le commerce de ces substances. Dans les années 70- , u e ou elle pid ie de o so atio de o aï e s’i stalla, a ec l’appa itio d’u e fo e de o aï e i solu le da s l’eau, le « crack », moins chère et plus efficace que la cocaïne en poudre. Depuis, le trafic, la consommation et les décès liés à la
o aï e e esse t d’aug e te OEDT, .
Figure 1. Structure du chlorhydrate de cocaïne.
La cocaïne est la deuxième drogue la plus consommée dans le monde, après le a a is et u e hausse glo ale de sa o so atio a t o se e au sei de l’U io Européenne lors de la dernière décennie (OEDT, 2006) (Figure 2). Cette augmentation est notamment associée à la baisse de son prix, la hausse de sa disponibilité sur le marché, la aisse de l’âge de p e i e p ise et la di e sifi atio des odes de o so atio OEDT, 2006). Sous forme de poudre, elle peut t e i hal e di e te e t ou hauff e afi d’i hale ses apeu s. L’effet se fait se ti au out de i utes et du e e i o heu e. Elle peut t e aussi fu e, i g e ou i je t e e i t a ei euse. Ce de ie ode d’ad i ist atio pe et de sentir rapidement les effets de la drogue (environ 10 secondes).
E fo tio du ode d’ad i ist atio , de la dose o so e et du sujet lui-même, la demi- ie de la o aï e peut s’ le e jus u’à i utes. U e g a de pa tie de la d ogue est métabolisée dans le foie dans les 2 heures qui suivent sa prise, et ceci selon 3 voies de dégradation (Fleming et al., 1990) :
(1) Celle des est ases h pati ues p oduisa t l’este de th le go i e ; (2) L’h d ol se spo ta e, sa s i te e tio e z ati ue, p oduisa t la
21
(3) La N-déméthylation de la cocaïne produisant un produit actif, la norcocaïne.
Figure 2. Usage au cours de la vie de substances psychoactives (hors alcool, tabac et cannabis) parmi les 18-44 ans.Evolution en France, entre 1992 et 2010. Amphé : amphétamines ;
XTC : extasy. Source : Baromètre Santé 1992, 1995, 2000, 2005, 2010, INPES exploitation OFDT www.ofdt.fr
“elo la oie d’ad i ist atio utilis e, l’ li i atio des ta olites de la o aï e se fait en proportions varia les A e et al., . Ils peu e t t e d te t s da s l’u i e jus u’à h ap s la p ise de o aï e, da s le lait ate el jus u’à h ap s la p ise de la d ogue, ai si ue da s les he eu et les o gles ap s de lo gues p iodes, puis u’ils gardent une trace des produits consommés au moment où ils poussent.
2.
La Méthamphétamine
La METH est un psychostimulant appartenant à la famille des phénéthylamines su stitu s. “a st u tu e est appa e t e à l’a ph ta i e AMPH et, e se le, fo t pa tie d’u g oupe de composés couramment désigné comme amphétamines (Fleckenstein et al., 2007) (Figure 3).
22
Figure 3.Structure de la méthamphétamine.
Largement distribuée pendant la deuxième guerre mondiale, la METH fut prescrite aux Etats-Unis o t e di e ses aladies, o e la d p essio et l’o sit , a a t d’ t e classée comme stupéfiant en 1970 (pour revue (Chiadmi and Schlatter, 2009)). Au début des a es , ette d ogue t a e se l’Atla ti ue pou appa aît e da s le a h des d ogues britanniques (Schifano et al., 2007), puis en France. La majeure partie de la production eu op e e est lo alis e e Eu ope de l’Est, ota e t e R pu li ue T h ue, e Slovaquie et en Bulgarie (Chiadmi and Schlatter, 2009). Connues sous diverses appellations a k, speed, pilule thaï, stal, i e, stal eth… , la o so atio de la METH a rapidement augmenté au cours de la dernière décennie puisque, sur le marché illicite, la METH, avec une pureté moyenne de 43 à 50%, fait partie des drogues les moins chères : un g a e de METH oûte e o e e € o t e € pou le g a e de o aï e OEDT, 2006).
La synthèse de la METH est facile et peut être réalisée à partir de produits chimiques et de médicaments relativement courants mais de plus en plus contrôlés. Les principaux précurseurs sont des médicaments délivrés en pharmacie sans ordonnance comme décongestionnants nasaux : la pseudo ph d i e, l’ ph d i e et la o ph d i e ou phénylpropanolamine (Barr et al., 2006). La synthèse de la METH nécessite aussi des p oduits hi i ues se o dai es tels ue le phospho e ouge, l’iode, le lithiu , l’a o ia a h d e, le tolu e et aut es sol a ts ases et a ides. L’a s fa ile à es p oduits a pe is le développement de nombreux laboratoires artisanaux de production de METH (Barr et al., 2006).
La METH se présente essentiellement sous forme de poudre blanche et cristalline, i odo e, pou a t t e dissoute fa ile e t da s l’eau ou l’al ool. Elle peut t e fu e,
23
aspirée par voie nasale ou injectée par voie intraveineuse. Elle peut même être compressée sous forme de comprimés et ingérée. Sa dégradation a lieu dans le foie et produit différents ta olites, do t l’AMPH et le -hydroxyméthamphétamine, tous les deux éliminés par voie rénale (Caldwell et al., 1972).
B-
Effets psychologiques et somatiques des psychostimulants
1. Effets psychologiques
i.
Administration aigue
L’ad i ist atio aigue de o aï e est g ale e t a o pag e d’u se ti e t de puissance et de maîtrise de soi. Le sujet se sent confiant, très énergique et euphorique. Cette phase est appelée « rush » ou « flash » et est de t pe o gas i ue. L’effet de la d ogue dure quelques heures et son intensité varie en fonction de la dose administrée, du mode de consommation ainsi que de la fréquence de la prise. Une fois les premiers effets disparus, le sujet se et ou e fa e à u e fatigue i te se et l’eupho ie est e pla e pa u e d p essio . C’est la phase dite de « crash ».
ii.
Administration chronique
L’effet plaisa t esse ti lo s des p e i es p ises de d ogue pousse certains individus à en reprendre, de façon contrôlée au départ puis de manière excessive et compulsive. On parle alors de dépendance aux drogues. Cette consommation excessive induit des dommages physiologiques et psychoaffectifs, aussi bien pour le sujet que pour son environnement. La consommation chronique de psychostimulants est notamment associée à des t ou les de la oi e, de l’hu eu ai si u’à des hallu i atio s et des d li es de t pe paranoïde (Koob et le Moal, 2006).
24
2.
Effets somatiques
Chez l’Ho e, uel ue soit la oie d’ad i ist atio , les ps hosti ula ts i duise t plusieurs manifestations somatiques. Ils stimulent le système nerveux sympathique provoquant une hausse du rythme cardiaque et respiratoire, une augmentation de la pression sanguine, une augmentation de la température et une dilatation des pupilles (pour revue : METH (Chiadmi and Schlatter, 2009) ; Cocaïne (Johanson and Fischman, 1989)). La o aï e est o ue pou so a ti it a esth si ue lo ale puis u’elle est apa le d’i hi e les canaux sodiques dépendants du voltage, de façon rapide et réversible. Elle produit ainsi une insensibilisation temporaire des terminaisons nerveuses. A plus long terme, la METH peut provoquer la mort en induisant des hémorragies cérébrales, des infarctus du myocarde, des uptu es h pati ues et ales ai si u’u e fo te h pe the ie pou e ue Al e tso et al., 1999)). Enfin, La METH et la cocaïne sont aussi des substances anorexigènes et provoquent des troubles du sommeil, ainsi que des sudations et des douleurs musculaires.
C-
Effets neurobiologiques
1.
Cibles moléculaires
En absence de toutes drogues, les transporteurs membranaires permettent aux monoamines extracellulaires de réintégrer la terminaison présynaptique où elles seront soit dégradées par les monoamines oxydases A et B, soit à nouveau stockées dans des vésicules de s tio da s le ut d’u e p o hai e utilisatio . Da s les deu as, le eu o e se et ou e p ot g o t e les effets to i ues i duits pa l’o datio des o oa i es présentes en g a de ua tit da s l’espa e s apti ue. Cet uili e est pe tu pa les psychostimulants.
i.
Mode d’a tio de la o aï e
La cocaïne agit principalement en se fixant sur les transporteurs membranaires de monoamines, inhibant leur fonctionnement et empêchant ainsi la recapture de la dopamine
25
(DA), de la sérotonine et de la noradrénaline par le neurone présynaptique. Par conséquent, il y a une augmentation des neurotransmissions dopaminergique, noradrénergique et sérotoninergique (Ritz et al, 1987 ; Giros et al, 1996 ; Broderick et al, 2003). Cette augmentation a notamment été observée, par microdialyse chez le rat, dans le cortex p f o tal CPF , le o au aud puta e CPu , le o au a u e s NA et l’ai e tegmentale ventrale (ATV) (Di Chiara and Imperato, 1988; Maisonneuve et al., 1990; Reith et al., . De plus, il a t o t ue l’ad i ist atio aigue de o aï e hez le at di i uait la synthèse de la DA et de la sérotonine dans plusieurs régions cérébrales telles que le CPF, le NAc, le CPu et l’a gdale asolat ale BLA Bau a et al., .
ii.
Mode d’a tio de la METH
Comme la cocaïne, la METH induit une augmentation des concentrations extracellulaires de la DA et de la sérotonine (Seiden et al., 1993). Mais, contrairement à la cocaïne qui agit principalement en bloquant les transporteurs membranaires de recapture de monoamines, la METH peut pénétrer dans la terminaison présynaptique soit par diffusion passive à travers la membrane plasmique grâce à son caractère lipophile, soit par transport actif en se liant aux transporteurs membranaires notamment les transporteurs de DA (DAT) (Giros et al, 1996). Elle induit alors une augmentation de la libération de monoamines dans la fente s apti ue. Plusieu s od les pe ette t d’e pli ue le a is e d’i e sio du transport de la DA par le DAT. En 1979, et grâce à des études visant à identifier les a is es d’a tio de l’AMPH, Fis he et olla o ateu s o t ta li u od le « d’ ha ge par diffusion » selo le uel le t a spo t de l’AMPH à l’i t ieur de la terminaison présynaptique par le DAT favorise la liaison de la DA intracellulaire à ces transporteurs et, en conséquence, la sortie extracellulaire de DA (Fischer and Cho, 1979). Selon un autre modèle, dit modèle « de déplétion vésiculaire », la concentration intracellulaire de DA serait à l’o igi e du t a spo t i e se assu pa les DATs “ulze et al., . U e fois da s le tosol, et g â e à sa si ila it st u tu ale a e la DA, l’AMPH p t e da s les si ules synaptiques, où elle modifie le gradient de concentration. Aussi la METH modifie la distribution des transporteurs vésiculaires de monoamines de type 2 (VMAT-2) de façon à diminuer leur disponibilité pour stocker les monoamines dans les vésicules de stockage
26
(Fumagalli et al., 1999; Hogan et al., 2000; Riddle et al., 2002). Ces phénomènes induisent une diminution de la recapture vésiculaire et favorisent une accumulation cytoplasmique de DA. Fonctionnant selon un gradient de concentration, les DATs assurent alors la libération de la DA da s la fe te s apti ue. C’est ai si ue la METH i duit u e aug e tatio di e te des o e t atio s e t a ellulai es de DA. Cet effet est espo sa le de l’i po ta te neurotoxicité provoquée par la METH (cf. chapitre 1-II. Toxicité de la METH).
2.
Circuits neuronaux impliqués dans les effets récompensants des
psychostimulants
Bie u’elles varient en fonction des effets comportementaux et pharmacologiques u’elles e ge d e t, les d ogues d’a us o t e o u la apa it d’i dui e u e se satio de plaisir (Robinson et Berridge, 2003). Ces effets sont aussi observés dans le cadre de renforçateurs naturels tels que la nourriture. En partant de cette observation, il a été sugg ue les d ogues d’a us affe te t u s st e de o pe se, o ale e t impliqué dans le contrôle de la motivation pour les récompenses naturelles telles que la ut itio et l’a ti it se uelle Ro i so et Be idge, . Ce s st e p se te des eu oadaptatio s i duites pa u e p ise h o i ue de d ogues ui pou aie t fou i l’ tat moti atio el essai e pou la ise e pla e de l’addi tio . Il eg oupe deu i uits majeurs : le s st e dopa i e gi ue so o ti oli i ue et le i uit de l’a gdale étendue (Figure 4).
27
Figure 4. Schéma représentatif des circuits neuronaux impliqués dans les effets renforçants des psychostimulants chez les rongeurs.
La o aï e et la METH i duise t la li atio de DA da s le o au a u e s, le o te et l’a gdale via une action directe sur les terminaisons dopaminergiques. En rouge est représenté le système dopa i e gi ue so o ti oli i ue. E leu est ep se t le i uit de l’a gdale te due.
odifi e d’ap s Koo a d Le Moal,
FC : cortex frontal ; C-P : noyau caudé putamen ; N Acc. : noyau accumbens ; VP : Pallidum ventral ; AC : commissure antérieure ; AMG : amygdale ; Hippo : hippocampe ; VTA : aire tegmentale ventrale ; SNr : substance noire pars reticulata ; LC : locus coeruleus ; Cer : cervelet.
i.
Description
du
système
dopaminergique
mésocorticolimbique
Le système dopaminergique mésocorticolimbique est constitué des neurones issus de l’ATV et ui p ojette t e s i des st u tu es t le phali ues ota e t le NA , ii des régions corticales comme le CPF, et (iii) des structures du système limbique telles que l’a gdale Figu e . Du fait de es p oje tio s, ette oie est i pli u e da s la gulatio des processus moteurs, affectifs, cognitifs, attentionnels et mnésiques (pour revue (Feltenstein and See, 2008)). Elle est aussi impliquée dans la motivation pour les récompenses naturelles comme la nutrition, la vie socio-affe ti e et l’a ti it se uelle (Robinson and Berridge, 2003).
28
L’ATV sous-te d l’effet des e fo çateu s atu els ai si ue les p op i t s e fo ça tes des d ogues d’a us Ro i s and Everitt, 1996; Koob and Le Moal, 1997). Elle est constituée p i ipale e t des eu o es dopa i e gi ues ui p ojette t e s le NA et le CPF. L’a ti it de es eu o es est gul e pa les eu o es GABAe gi ues p se ts da s l’ATV, les afférences excitatrices glutamatergiques en provenance du CPF, et (3) les afférences inhibitrices GABAergiques en provenance du NAc (pour revue : (White, 1996)).
Figure 5. Schéma simplifié de la voie mésocorticolimbique chez les rongeurs.
Les eu o es dopa i e gi ues issus de l’ai e teg e tale e t ale ATV p ojette t e s le o au accumbens (Nac) et le cortex préfrontal (CPF) (rouge). Ces neurones sont contrôlés par les systèmes glutamatergiques (bleu) et GABAergiques (orange). Les neurones glutamatergiques proviennent du CPF, de l’a gdale, de l’hippo a pe et du Bed Nu leus of the “t ia Te i alis BN“T , et p ojette t ota e t e s le Na et l’ATV. Les eu o es GABAe gi ues so t issus du NA et p ojette t e s l’ATV. (Kauer and Malenka, 2007)).
CPF : cortex préfrontal ; NAc : noyau accumbens ; VP : pallidum ventral ; BNST : bed nucleus of the stria terminalis ; LH : hypothalamus latéral ; VTA : aire tegmentale ventrale.
Le NAc est une structure cérébrale située dans la partie ventrale du striatum. Sur les plans morphologique et fonctionnel, il peut être divisé en deux sous-régions : le core et le shell (Di Chiara, 2002). Il reçoit majoritairement des afférences dopaminergiques provenant de l’ATV, des aff e es e itat i es gluta atergiques provenant du CPF, du cortex i gulai e a t ieu , de l’a gdale et de l’hippo a pe et e oie des p oje tio s i hi it i es GABAe gi ues e s la su sta e oi e et l’ATV pou e ue : (White, 1996)). Ces connexions
29
lui o f e t le ôle d’i t g ateur des informations sensorielles, affectives et cognitives asso i es à la p ise de d ogue, lui pe etta t d’ alue les effets e fo ça ts et de déclencher les réponses locomotrices (Koob and Le Moal, 1997).
ii.
Des iptio du i uit de l’a
gdale te due
Il existe un substrat anatomique commun aux effets récompensants induits par les d ogues et les affe ts gatifs i pli u s da s la ise e pla e d’u e o so atio o pulsi e de d ogues. Cette e tit eu oa ato i ue est l’« amygdale étendue ». Elle est constitu e d’u o ti uu de st u tu es t le phali ues eg oupa t le “hell du NA , le noyau du lit de la strie terminale (BNST : Bed Nucleus of the Stria Terminalis) et le noyau e t al de l’a gdale CeA pou e ue Alheid, . Ces t ois gio s pa tage t des si ila it s au i eau a hite tu e et o e ti it . L’a gdale te due eçoit des aff e es p o e a t des st u tu es li i ues telles ue l’hippo a pe et la BLA, et e oie des eff e es e s la pa tie dia e du pallidu e t al, l’ATV et l’h pothalamus latéral (pour e ue Alheid, . G â e à es o e tio s, l’a gdale te due o stitue u e i te fa e entre les structures limbiques impliquées dans la régulation des émotions et le système oteu e t ap a idal. Pa ailleu s, l’a gdale te due ontient plusieurs systèmes eu o hi i ues asso i s au effets e fo ça ts des d ogues d’a us e . dopa i e et peptides opioïdes , ai si ue des o posa ts de l’a e HPA et des s st es du st ess e . GABA, CRF et dynorphine) (Koob and Le Moal, 2005). Ainsi, elle représente un centre d’i t g atio des p o essus h do i ues et des p o essus du st ess, p oduisa t u tat émotionnel sous-te da t le e fo e e t gatif i duit pa les d ogues d’a us Koo a d Volkow, 2010).
iii.
Interactions des psychostimulants avec le circuit de la
récompense
Les eu o es dopa i e gi ues situ s da s l’ATV et p ojeta t e s le NA , e pa ti ulie le “hell, so t a ti s pa l’ad i ist atio de d ogues d’a us et leu a ti atio sous-tend les effets récompensants de ces drogues. Premièrement, du a t l’auto-administration de cocaïne, une augmentation de la libération de DA est détectée dans le NAc
30
grâce à la microdialyse in vivo (Di Chiara and Imperato, 1988). Deuxièmement, une destruction sélective des neurones dopaminergiques mésocorticolimbiques ou une lésion spécifique des fibres dopaminergiques dans le NAc, induites par la toxine 6-hydroxydopamine (6-OHDA), li i e t le o po te e t d’auto-administration de cocaïne (Roberts et al., 1980). Pa ailleu s, il a t o t ue les ats s’autoad i ist e t l’AMPH (Ikemoto et al., 2005) et la cocaïne (Ikemoto, 2003) dans le Shell, et non dans le core, du NAc. L’ad i ist atio aigue de la plupa t des d ogues d’a us i duit gale e t u e augmentation des concentrations extracellulaires de DA dans le Shell du NAc (Pontieri et al., 1995). Le Shell semble donc être plus important que le core dans la récompense aux drogues. Des études pharmacologiques ont permis de démontrer que les effets récompensants induits par la cocaïne font intervenir les récepteurs dopaminergiques D1 et D , puis ue l’ad i ist atio d’a tago istes du epteu D di e te e t da s le “hell du NA di i ue le o po te e t d’auto-administration de cocaïne (Caine et al., 1995) et (2) l’ad i ist atio de l’eti lop ide, u a tago iste du epteu D , di i ue l’auto-administration de cocaïne chez le singe (Bergman et al., 1990).
3.
Interactions des psychostimulants avec les systèmes neuronaux
impliqués dans la réponse au stress
Pou u g a d o e de pe so es d pe da tes, l’addi tion peut ne pas être la seule pathologie engendrée par la consommation de drogues. En effet, des syndromes de d p essio et d’a i t so t le plus f ue e t asso i s. “elo les tudes pid iologi ues, à % des pe so es d pe da tes à l’al ool, la ocaïne et les autres psychostimulants présentent aussi des symptômes dépressifs et/ou anxieux (Kreek et al., . E pa ta t de e o stat, de o euses tudes se so t i t ess es à l’i te a tio e t e la o so atio de o aï e et l’ tat de st ess des individus, et ont permis de mettre en évidence que la cocaïne et le stress font intervenir les mêmes systèmes hormonaux et eu o hi i ues. Je p se te ai, da s ette pa tie, l’effet des ps hosti ula ts su deu systèmes majeurs impliqués dans la réponse au stress : l’a e h pothala
o-hypophyso-31
su alie , plus ou a e t o u sous le o d’a e HPA, et le eu opeptide corticotropin releasing factor (CRF).
i. L’a e HPA : principal système physiologique de réponse au
stress.
L’a e HPA est le p i ipal système physiologique de réponse au stress et de régulation de l’ho ostasie de l’o ga is e. Co e so o l’i di ue, il est o stitu de t ois niveaux Figu e : le o au pa a e t i ulai e de l’h pothala us PVN : paraventricular nucleus), qui contient des neurones exprimant le CRF, et forme un lien entre le système e eu et le s st e e do i ie g â e au p oje tio s u’il e oie e s l’h poph se ; (2) l’h poph se, ui est situ e à la ase du e eau et est di is e e deu lo es : l’ad oh poph se (hypophyse antérieure) et la neurohypophyse (hypophyse postérieure) ; (3) les glandes surrénales, situées au-dessus des reins, et divisées en médullosurrénale et corticosurrénale. Quand un élément stressant survient, le PVN libère le CRF dans le système porte hypothalamo-h poph sai e. Cette eu oho o e attei t l’h poph se, où elle i duit la s th se et la s tio de l’ho o e o ti ot ope ACTH, Adreno CorticoTropic Hormone). L’ACTH est hi ul e pa le sa g jus u’au gla des o ti osu ales où elle induit la libération de glucocorticoïdes (cortisol chez les primates et corticostérone chez les o geu s . A leu tou , es ho o es o t la apa it de gule l’a ti it de l’a e HPA , pa rétrocontrôle négatif, au niveau de différentes structures cérébrales, telles que l’hippo a pe et l’a gdale pou e ue Nestle et al., .
32
Figure 6. L’a e h pothala o-hypophyso-surrénalien (axe HPA) chez l’Ho e.
Les glu o o ti oïdes o t ôle t l’a ti it de l’a e HPA g â e au afférences neuronales excitatrices, provenant de l'amygdale, et inhibitrices, provenant de l'hippocampe. (d’ap s Nestle et al., 2002)
Dans le cadre de la recherche sur la dépendance aux drogues, il a été montré que l’ad i ist atio aigue de ps hosti ula ts, hez l’Ho e et les o geu s, p oduit u e a ti atio de l’a e HPA se la le à elle i duite pa le st ess pou e ue “a ai et al., . E effet, hez l’Ho e, l’ad i ist atio aigue de o aï e pa oie i t a ei euse induit une augmentation du taux sanguin de cortisol chez des sujets ayant déjà consommé la drogue (Baumann et al., 1995), ainsi que chez des sujets naïfs (Heesch et al., 1995). Chez les
33
o geu s, u e i je tio lo ale de o aï e au i eau de l’h pothala us “aphie et al., ou du striatu Ike oto a d Goede s, i duit u e a ti atio de l’a e HPA, esu e pa une augmentation des niveaux sanguins de corticostérone. Cette activation dépend des epteu s du CRF, puis ue l’ad i ist atio e t ale d’a tago iste du epteu CRF lo ue l’aug e tatio de o ti ost o e hez des ats a a t eçu de la o aï e “a ai et al., . Pa ailleu s, l’ad i ist atio h o i ue de ps hosti ula ts i duit u e h pe a ti it soute ue de l’a e HPA e ap s l’a t des p ises de d ogue : comparés à des rats naïfs, des rats ayant subi 3 à 6 semaines de « binge » (c'est-à-dire des épisodes de prise, répétés et rapprochés) de cocaïne présentent une augmentation du poids des glandes surrénales et du taux basal de corticostérone, 12h après la dernière injection reçue (Sarnyai et al., 1998). De a i e i t essa te, l’i e se est o se da s u p oto ole d’auto-administration de cocaïne : h ap s l’a t de la p ise de d ogues, les ats p se te t u e di i utio du tau basal de corticostérone associée à une augmentation de la densité des récepteurs aux glu o o ti oïdes da s l’hippo a pe Ma ts h a d Goede s, .
ii. Le système du CRF : un neuropeptide impliqué dans la
réponse au stress
Le neuropeptide CRF est largement exprimé dans le cerveau, avec une densité importante dans le PVN, le BNST, le CeA, le locus coeruleus et le noyau du raphé. Du fait de cette distribution, le CRF est impliqué à la fois dans la réponse au stress et dans la d pe da e au d ogues d’a us. Il peut se lie à deu t pes de écepteurs : le récepteur CRF de type 1 (CRF1) et de type 2 (CRF2). Le récepteur CRF1 a une expression centrale et est très e p i au i eau de l’h poph se, ta dis ue le epteu CRF p se te u e de sit plus importante au niveau périphérique, et est exprimé dans certaines structures cérébrales telles ue le septu , l’h pothala us et le o au du Raph pou e ue A o elius et al., 1999)).
Co e l’ad i ist atio , aigue ou h o i ue, de o aï e a ti e l’a e HPA de a i e dépendante du système CRF (Sarnyai et al., 1992), les chercheurs se sont intéressés à savoir
34
si la cocaïne peut moduler le système du CRF. Il a ainsi été démontré que l’ad i ist atio aigue de cocaïne à des rats induit une diminution de la concentration de CRF dans l’h pothala us, le NA , l’hippo a pe et le o te f o tal, ais pas da s l’a gdale, où u e augmentation de CRF a été observée (Sarnyai et al., 1993). De même, dans une étude de microdialyse in vivo, Ri hte et olla o ateu s o t t ou ue l’ad i ist atio p t e de cocaïne induit une augmentation de la synthèse et de la libération de CRF dans le CeA (Richter et al., 1995). Par ailleurs, le CRF est exprimé dans des régions appartenant au s st e dopa i e gi ue so o ti oli i ue NA , o te f o tal et à l’a gdale te due (BNST, CeA, NAc). De ce fait, outre son action de régulateur des réponses hormonales induites par les facteurs stressants, le système CRF peut être engagé dans différents aspects de la d pe da e au d ogues d’a us. L’a t de o so atio de e tai es d ogues, telles ue l’al ool, i duit u e aug e tatio de la li atio de CRF da s la CeA et le BN“T Fu k et al., 2006). De plus, l’i je tio d’a tago iste du CRF au i eau de l’a gdale lo ue les effets anxiogènes induits par le sevrage à la cocaïne (pour revue (Sarnyai et al., 1998)), et l’ad i ist atio d’a tago istes du CRF da s le BN“T pe et de lo ue la e hute i duite pa le st ess pou la o aï e et l’h oï e “haha et al., .
D-
Dépendance aux psychostimulants
1. Description de la dépendance
i.
Diagnostic
La dépendance aux drogues est un désordre chronique caractérisé par (1) une e he he o pulsi e de la d ogue, u e pe te de o t ôle de la p ise, et l’ e ge e d’u tat otio el gatif ua d la d ogue ’est pas a essi le Koo a d Le Moal, 1997). De os jou s, le diag osti de d pe da e à u e d ogue d’a us se ase su les it es du DSM-IV (Diagnostic and Statistical manual of Mental disorders - 4ème édition ; American Psychiatric Association, 1994) (Tableau 1) et du CIM-10 (Classification statistique Internationale des Maladies et des problèmes de santé connexes – 10ème édition ; Organisation Mondiale de la Santé, 1992) (Tableau 2).
35
Tableau 1. Critères de la dépendance selon le DSM-IV
La dépendance est une utilisation inappropriée d’u e d ogue e t aî a t u e alt atio du fo tio e e t ou u e souff a e, li i ue e t sig ifi ati e, et a a t is e pa l’appa itio d’au moins trois des symptômes cités ci-dessous et ela su u e p iode d’u a :
1- Une tolérance caractérisée soit par une aug e tatio des doses o so es afi d’o te i u effet si ilai e, soit pa u e di i utio de l’effet i duit e as d’utilisatio p t e d’u e même dose.
2- “e age a a t is soit pa l’appa itio d’u s d o e de se age suite à l’a t de consommatio de la d ogue, soit pa la p ise de la e su sta e ou d’u e su sta e appa e t e da s le ut de soulage ou d’ ite les s ptô es de se age.
3- Substance souvent prise en quantité supérieure ou pendant un laps de temps plus long que ce que la personne a envisagé.
4- Un désir persistant pour diminuer ou arrêter la prise de drogue
5- Un temps de plus en plus important est consacré à la recherche de la drogue
6- A a do des a ti it s p ofessio elles et so iales à ause de l’utilisatio de la d ogue 7- Une poursuite de la consommation de la drogue, malgré la conscience des problèmes
u’elle e ge d e
DSM-IV : Diagnostic and Statistical manual of Mental disorders - 4ème édition ; American Psychiatric Association, 1994
Tableau 2. Critères de la dépendance selon le CIM-10
Le diag osti d’u e d pe da e essite l’appa itio d’au moins trois des manifestations suivantes, en même temps, au cours de la dernière année :
1- D si puissa t ou o pulsif d’utilise u e su sta e ps hoa ti e
2- Diffi ult s à o t ôle l’utilisation de la substance (début ou interruption de la o so atio au i eau de l’utilisatio
3- E e ge e d’u s d o e de se age ph siologi ue ua d le sujet di i ue ou a te la o so atio d’u e su sta e ps hoa ti e, o e e t oig e t la su e ue d’u sy d o e de se age a a t isti ue de la su sta e, ou l’utilisatio de la e su sta e
ou d’u e su sta e appa e t e pou soulage ou ite les s ptô es de se age
4- Mise e ide e d’u e tol a e au effets de la su sta e ps hoa ti e : le sujet a besoin d’u e ua tit plus i po ta te de la su sta e pou o te i l’effet d si
5- D si t t p og essif pou toute aut e a ti it au p ofit de l’utilisatio de la su sta e psychoactive, et augmentation du temps passé à se la procurer, la consommer ou récupérer ses effets
6- Poursuite de la consommation de la substance malgré ses conséquences nocives.
CIM-10 : Classification statistique Internationale des Maladies et des problèmes de santé connexes – 10ème édition ; Organisation Mondiale de la Santé, 1992
36
ii.
Vulnérabilité
Nombreuses sont les personnes qui expérimentent les drogues mais 15 à 20% seulement en deviennent dépendantes (Piazza and Le Moal, 1996). Cette dépendance est caractérisée par une prise de drogues de façon compulsive et non contrôlée. Le passage d’u e p ise de d ogue o t ôl e à u e p ise o pulsi e est le sultat d’i te a tio s complexes entre trois principaux facteurs rendant un individu « vulnérable » aux effets addi tifs d’u e d ogue K eek et al., Figu e .
(1) les déterminants génétiques : l’h ita ilit de e tai es addi tio s a t ise e évidence grâce à des études épidémiologiques réalisées auprès de jumeaux et de personnes adoptées (Johnson et al, 1996). Aussi, des études de génome ont montré que les personnes a a t l’all le (A1) du gène codant pour le récepteur dopaminergique D2 aurait un risque le de p dispositio à l’addi tio à l’al ool Blu et al., . Pa ailleu s, le passage au modèles animaux a permis de sélectionner des souches de rongeurs en fonction de s’ils s’autoad i ist e t fa ile e t ou pas les d ogues Koste et al., ; Brodkin et al., 1998). De plus, des études réalisées chez des souris knock-out pour le transporteur dopaminergique (DAT) ont montré que, contrairement à des souris wild type, les souris KO ne présentent pas d’aug e tatio de l’a ti it lo o ot i e e po se au ps hosti ula ts Gi os et al., 1996).
(2) les effets induits par les drogues : Quand elles sont consommées de façon chronique, les drogues induisent des neuroadaptations moléculaires et cellulaires (pour revue (Vanderschuren and Kalivas, 2000)) qui joueraient un rôle primordial dans la transition d’u usage o asio el à u e p ise o pulsi e de d ogues d’a us. Ces eu oadaptatio s o t pour cible principale le système de récompense qui est impliqué dans la motivation pour les récompenses naturelles (Robinson and Berridge, 2003). Les comportements dirigés vers la prise de drogue sont alors renforcés au détriment des comportements naturels, entraînant ainsi un état pathologique chronique (Kalivas and O'Brien, 2008). Ces mécanismes adaptatifs sont développés dans la partie suivante de ce chapitre.
37
(3) les facteurs environnementaux : De nombreuses études ont permis de mettre en ide e l’i flue e des e p ie es de ie su la sensibilité des individus aux drogues d’a us. Ai si, des o ditio s de ie gati es a oi de pau es elatio s fa iliales et sociales, avoir un faible statut socio-économique, avoir déjà été exposé à un stress i po ta t… o stitue aie t des fa teu s de isque de développement de dépendance (McEwen, 2000; Goeders, 2002; McEwen, 2007), tandis que des expériences de vie positives
i eau d’ tudes le s, t s o es elatio s fa iliales, o statut so io- o o i ue… seraient des facteurs de protection contre l’appa itio d’u e d pe da e au d ogues (Jessor and Jessor, 1980). Tous ces facteurs vont pouvoir moduler le cerveau et rendre ainsi l’i di idu plus ou oi s ul a le au d ogues d’a us. Le hapit e de e a us it de thèse est consacré à ces facteurs environnementaux.
Figure 7. L’i te a tio e t e les fa teu s g ti ues, e vi o e e tau et asso i s à la d ogue pe et d’ value la vul a ilit au d veloppe e t de l’addi tio .
Les facteurs associés à la drogue regroupent les changements induits par la drogue sur le taux d’ARN RNA le els et le p ot o e e g al ; s’ajoute t à ela les fa teu s e i o e e tau , o e le st ess et les e p ie es pass es, et le pat i oi e g ti ue de l’i di idu “NP : Single Nucleotide Polymorphism). d’ap s K eek et al,
2.
Mécanismes impliqués dans la dépendance
Co e ous l’a o s d it p de e t, l’ad i ist atio de ps hosti ula ts active la neurotransmission dopaminergique (cf. partie 1.C- Effets neurobiologiques), et
38
gluta ate gi ue da s le NA , l’hippo a pe, l’ATV et la su sta e oi e “N Kali as a d Duffy, 1995; Smith et al., 1995). Cette libération de glutamate serait dépendante des récepteurs dopaminergiques D1 (Kalivas and Duffy, 1995) et est régulée par une boucle de rétrocontrôle striatothalamocortical (Kalivas and Duffy, 1995; Mark et al., 2004). En effet, u e aug e tatio de l’a ti it des epteu s dopa i e gi ues D p se ts au i eau des terminaisons striatales présynaptiques provoquerait une stimulation du système GABA nigrostriatal. Il y a alors augmentation de la libération de GABA dans la SN pars reticulata, induisant une inhibition du signal GABA dans le système nigrothalamique, et donc d si hi itio des aff e es thala o o ti ales. D’où l’aug e tation de la libération de glutamate dans le striatum (Mark et al., 2004) (Figure 8).
Figure 8. Boucle striato-thalamocorticale impliquée dans la régulation du système glutamatergique.
U e aug e tatio de l’a ti it des epteu s dopaminergiques D1 (1), présents au niveau des terminaisons striatales présynaptiques, provoquerait une stimulation du système GABA nigrostriatal. Il y a alors augmentation de la libération de GABA dans la substance noire pars reticulata (2), induisant une inhibition du signal GABA dans le système nigrothalamique (3), et, par conséquent, une d si hi itio des aff e es thala o o ti ales . D’où l’aug e tatio de la li atio de gluta ate dans le striatum (5). d’ap s Ma k et al.,