LE BISON DES PRAIRIES ENTRE AIRE PROTÉGÉE ET
TERRES AGRICOLES : CAUSES, CONSÉQUENCES ET
PERSPECTIVES DE GESTION
Thèse
Marie Sigaud Doctorat en Biologie Philosophiae doctor (Ph.D.)Québec, Canada
© Marie Sigaud, 2018LE BISON DES PRAIRIES ENTRE AIRE PROTÉGÉE ET
TERRES AGRICOLES : CAUSES, CONSÉQUENCES ET
PERSPECTIVES DE GESTION
Thèse
Marie Sigaud
Sous la direction de :
iii
Résumé
Le travail réalisé au cours de cette thèse aborde la thématique complexe de la place des espèces de la faune sauvage dans des habitats qui sont de plus en plus modifiés par les activités humaines. Nous prenons pour modèle d’étude la population de bisons des prairies (Bison bison bison) établie dans le parc national de Prince Albert (Canada) qui utilisent les terres agricoles exploitées bordant le parc. Dans un premier temps, nous démontrons que ce sont les gains énergétiques qui guident principalement la sélection d’habitat des bisons au détriment d’autres facteurs contribuant à la valeur adaptative, tels que le risque de mortalité lié à la chasse. Les parcelles agricoles constituent alors pour cette population un piège écologique, soit un habitat qui est préféré ou également préféré à d’autres habitats disponibles pourtant de meilleure qualité. L’utilisation des parcelles se diffuse parmi la population grâce aux décisions collectives prises au sein du groupe sur la base de l’expérience passée des membres du groupe. La diminution de la taille de la population est concomitante à la diffusion de ce comportement. Nos résultats nous renseignent sur les effets négatifs que peut avoir l’utilisation des milieux anthropisés et sur comment des mécanismes adaptatifs en milieu naturel, comme l’apprentissage social, peuvent se révéler délétères dans des milieux perturbés par l’homme. Nous étudions ensuite les stratégies comportementales mises en œuvre par les bisons pour échapper aux menaces constituées, à la fois, par leur prédateur naturel (le loup gris, Canis lupus) et par les activités humaines. Les bisons réagissent à la proximité du loup gris quand ils sont dans une prairie naturelle uniquement de nuit en écourtant leur temps de résidence. Alors qu’ils sélectionnent les parcelles agricoles principalement la nuit quand les activités humaines sont au plus bas. Ces stratégies divergentes nous renseignent sur les capacités des bisons à intégrer des informations nouvelles et à ajuster leur comportement en fonction de leur expérience passée. Le déclin de la population nous indique toutefois que ce type de stratégie, certainement efficace pour maximiser le temps passé sur les parcelles agricoles tout en minimisant le risque de perturbations, reste largement insuffisant pour prémunir les bisons contre la mortalité induite par la chasse. Pour mieux comprendre les déplacements des bisons et guider la gestion de la population, nous explorons les
iv
propriétés du réseau de sentiers créés et entretenus par les bisons pour se déplacer entre les prairies naturelles. Le réseau est très redondant avec de nombreux sentiers indépendants connectant la même paire de prairies. Le nombre de sentiers et de connexions diminue en même temps que le nombre d’individus. Ces résultats nous renseignent sur la capacité de résistance du réseau aux perturbations, mais également sur l’influence de la taille de la population sur la connectivité fonctionnelle. L’utilisation des milieux anthropisés par les espèces de la faune peut également avoir des impacts sur les écosystèmes. Nous décrivons une situation souvent négligée dans le domaine de la conservation : le conflit de conservation, situation au cours de laquelle un objectif de conservation entre en conflit avec un ou plusieurs objectifs d’un autre programme. Les bisons des prairies, espèce au statut vulnérable, transportent un grand nombre de graines d’espèces de plantes non indigènes acquises sur les parcelles agricoles et qui sont potentiellement envahissantes. Ils favorisent l’installation de ces espèces en créant des conditions favorables, comme au niveau des « wallows » (zone dépourvue de végétation en son centre créée par les bisons en se roulant au sol) ou des sentiers créés au cours leurs déplacements. Les bisons en tant que vecteurs d’espèces non indigènes représentent un danger pour la restauration des prairies à fétuques, objectif de conservation prioritaire pour Parcs Canada. En conclusion, ce travail apporte des éléments de compréhension sur le comportement des animaux face à des contraintes émergentes dans les milieux anthropisés et sur les conséquences pour les animaux et les écosystèmes. Il soulève la question de la pérennité de certaines populations animales dans ces milieux anthropisés en l’absence de mesures de gestion adaptées.
v
Table des matières
Résumé ... iii
Table des matières ... v
Liste des figures ... viii
Liste des tableaux ... x
Remerciements ... xiv
Avant-propos ... xvi
Introduction ... 1
1. Le choix de l’habitat ... 2
1.1. Les indices de la qualité de l’habitat ... 2
1.2. Décisions collectives : influence du groupe ... 3
1.3. Le piège écologique : émergence et sévérité ... 4
2. Causes et conséquences de l’utilisation des milieux anthropisés ... 6
2.1. Nouveaux paysages, nouvelles ressources, nouvelles stratégies ... 6
2.2. Paysage de la peur et les multiples sources de risque ... 7
2.3. Conflits homme-faune sauvage ...Erreur ! Signet non défini. 2.4. Plasticité comportementale ... 7
3. Les réseaux spatiaux et leur utilisation comme outil de gestion ... 9
3.1. Connectivité et répartition spatiale ... 9
3.2. La théorie des graphes ... 10
3.3. Influencer les patrons de déplacements : un outil de gestion ... 11
4. Les conflits de conservation ... 11
4.1. Définition et émergence... 11
4.2. Le cas de la dissémination de plantes envahissantes dans des milieux sensibles par une espèce animale d’intérêt patrimonial... 12
5. Modèle d’étude : le bison des prairies ... 13
6. Objectif et organisation de la thèse ... 15
Chapitre 1. ... 16
Chapitre 2. ... 16
Chapitre 3. ... 17
Chapitre 4. ... 17
Chapitre 1. Collective decision-making promotes fitness loss in a fusion-fission society ... 18
Résumé ... 19
Abstract ... 20
Introduction ... 21
Methods... 23
Study area and bison data ... 23
Agricultural lands as an ecological trap ... 23
Testing selection for the most profitable plants ... 24
Assessing fitness consequences of selecting agricultural lands ... 25
Social learning promotes selection of an ecological trap ... 26
vi
Results ... 28
Discussion ... 29
Agricultural lands as an ecological trap ... 30
Social learning promotes selection of an ecological trap ... 32
Supporting Information ... 38
Chapitre 2. Coping with multiple patterns of risk: bison develop complementary tactics to avoid humans and wolves ... 54
Résumé ... 55
Abstract ... 56
Introduction ... 57
Methods... 59
Study area and relocation data ... 59
Human activities and broad-scale selection of agricultural lands ... 60
Fine-scale movement responses to predation risk ... 61
Step length ... 62
Results ... 63
Broad-scale selection of agricultural land and human activities ... 63
Bison ... 63
Wolf ... 64
Fine-scale movement response to predation risk ... 64
Distance travelled ... 64
Discussion ... 65
Acknowledgements ... 68
Supporting Information ... 75
Chapitre 3. Spatial network properties and density dependence in functional connectivity for free-ranging bison... 78
Résumé ... 79
Abstract ... 80
Introduction ... 81
Methods... 83
Study Area and bison population ... 83
Network structure and connexion pattern ... 84
Results ... 84
Discussion ... 86
Chapitre 4. Conflicting conservation programmes When a threatened wildlife species jeopardizes the conservation of a rare plant community . 90 Résumé ... 91
Abstract ... 92
Introduction ... 93
Methods... 95
Study area and potential conflicting conservation programmes ... 95
Dispersal of exotic seeds by bison ... 95
Spatial distribution of exotic plant probability of occurrence... 96
Statistical analysis ... 97
Results ... 98
vii
Acknowledgements ... 102
Supporting Information ... 107
Conclusion générale ... 112
Implications pour la conservation ... 115
Perspectives de recherche ... 117
viii
Liste des figures
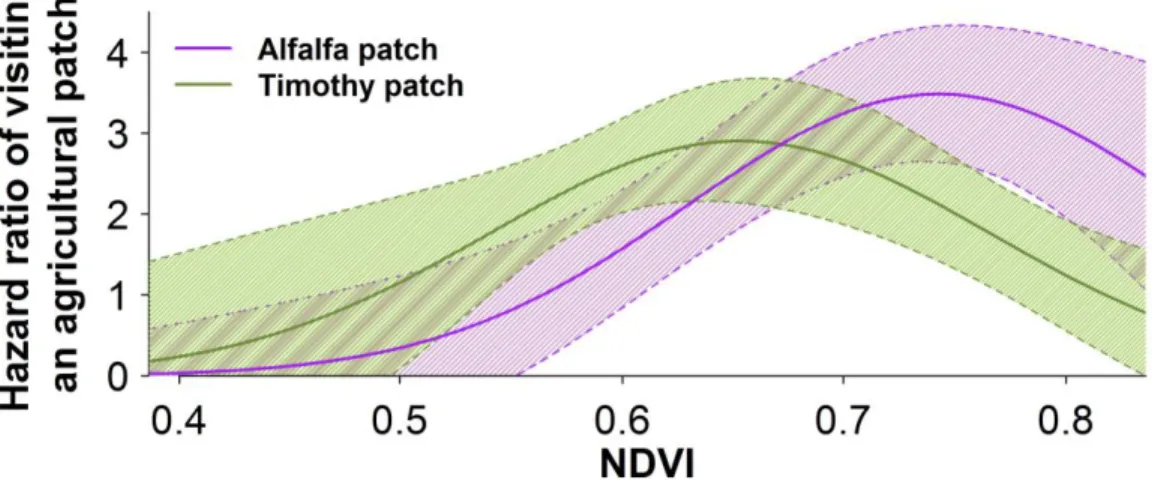
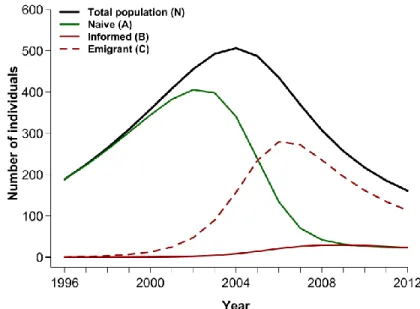
Figure 1.1. The study area in and around Prince Albert National Park, Saskatchewan (Canada). Dashed green line and solid purple line represent 90% contours of kernel distributions of all bison GPS locations from 1997 to 1999 and 2005 to 2013, respectively. Between the 1990s and late 2000s plains bison expanded their range to include 236% more agricultural lands... 35 Figure 1.2. Selection coefficients for meadows and agricultural patches by female plains bison in and around Prince Albert National Park, Saskatchewan (Canada) from July to October 2005-2013 (Table S1.4). Estimates derived from a mixed-effects Resource Selection Function (RSF, see Appendix S1.3). A positive coefficient indicates selection for a given land cover type with respect to forest, the reference cover type. Avoidance of agricultural patches prior to 2006 shifted to selection and increased thereafter (dashed orange line). Radio-collared bison (n = 40) selected meadows (continuous yellow line) consistently across all years. Population size (brown line, with 95% CI) declined from 2005 to 2010, then remained relatively stable (Merkle et al. 2015a). ... 36 Figure 1.3 : Temporal dynamics of knowledge acquisition about an ecological trap and simultaneous changes in population size. (a) Predicted population size (eqn 4) over time based on the 10 best fitting models with (f > 0) and without (f = 0) the social learning component, together with the observed bison population dynamics in and around Prince Albert National Park (PANP), Saskatchewan (Canada). (b) Percentage of naïve individuals in the population, A(t)/N(t), as predicted from the best fitting collective behavioural model, see eqns 1-4, with (f > 0) and without (f = 0) the social learning component, and from observed radio-collared bison (sample size of bison indicated). ... 37 Figure S1.1. Influence of the vegetation cultivated on the recurrence of visiting an agricultural patch. Chance of visiting an agricultural patch (specified as a hazard ratio, exp[β] and its 95% CI in dotted lines) by female plains bison (n = 18) at a given time as a function of available green biomass (indexed using the Normalized Difference Vegetation Index [NDVI]) and type of agricultural land near Prince Albert National Park, Saskatchewan (Canada) during August to October, 2011 to 2012. Types of agricultural lands were either dominated by alfalfa (purple line) or timothy (green line) (Table S1.1). Estimates were derived from a Cox proportional hazards (CPH) regression model with a counting process approach (see Appendix S1.3). ... 46 Figure S1.2. Temporal dynamics of individual behavioural morphs and changes in total population size. Plotted are: number of naïve individuals (A), see eqn 1; informed individuals (B), see eqn 2; emigrants (C), see eqn3; and total population size (N), see eqn 4, as predicted from the best performing collective behavioural model with a social learning parameter (see Appendix S1.4 and Table S1.6). ... 47 Figure 2.1. Average percentage of locations and 95% confidence intervals observed at each hour of the day on meadows and agricultural land for female plains bison (n = 46) in and around Prince Albert Park, Saskatchewan (Canada) from July to October 2005-2015 and percentage of human activities recorded on neighbouring agricultural lands (n = 13) from August to October 2012-2013. ... 72
ix
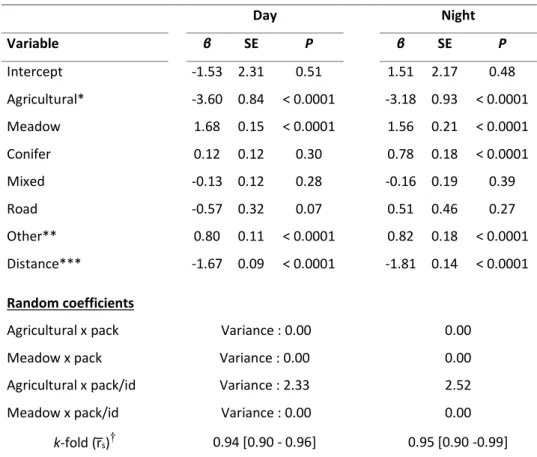
Figure 2.2. Selection coefficients for meadows and agricultural lands at day (a) and night (b) by female plains bison (n = 46) in and around Prince Albert National Park, Saskatchewan (Canada) from July to October 2005-2015. Estimates derived from a mixed-effects Resource Selection Function (Table S2.1). A positive coefficient indicates selection for a given land cover type with respect to deciduous forest, the reference cover type. ... 73 Figure 2.3. Variations of the average hourly distance travelled per year by female plains bison (n = 46) in and around Prince Albert National Park, Saskatchewan, Canada from July to October 2005 to 2015 at dawn, day, dusk and night. ... 74 Figure 3.1. Trails used by plains bison (thick lines), together with links predicted by the Minimum planar graph (thin grey lines) connecting meadow patches (gray polygons, darker polygon representing one of 28 surveyed meadow patches) within the forest matrix of Prince Albert National Park, Canada. Trails A and B, C and D are redundant. Trails C, D and E, are materialising links not predicted by the MPG... 89 Figure 4.1. Overview of the study area, Prince Albert National Park, Saskatchewan, Canada, (dashed black line) and locations of female plains bison (n = 21) during autumns 2011 - 2013. Land cover types displayed are : forest, water, meadows and agricultural land. Identified patrimonial fescue grasslands are delimited as well as potential exotic plant species introduction points (i.e., preferred points used by bison to access agricultural land) on the park boundary. ... 104 Figure 4.2. Probability of occurrence of two exotic plant species along bison trails in deciduous forest and around wallows (based on best performing models of Table 3.1), in Prince Albert National Park, Saskatchewan, Canada, in relation to Euclidian distance to the nearest introduction point from surrounding agricultural areas. ... 105 Figure 4.3. Predicted spatial variation in probability of occurrence of exotic plant species throughout the plains bison range (a-c) and the distributions of the predicted probability of exotic plant species occurrence within fescue grassland patches (d-f) in Prince Albert National Park, Saskatchewan, Canada. Predictions are based on best performing models (Table 3.1) for clover (Trifolium spp.) on bison trails (a-d), timothy (Phleum pratense) on bison trails (b-e) and timothy around bison wallows (c-f). ... 106 Figure S4.1. (a) Number of intact seeds recovered in 238 plains bison feces collected in Prince Albert National Park, Saskatchewan, Canada, during autumn 2012 for the 15 dominant species, on a logarithmic scale. (b) Percentages of fecal samples with at least one seed per species. ... 110 Figure S4.2. Distribution of average distance to the nearest introduction point (bars) for 18 collared female bison relocated every 3 hours from July to October 2010 to 2012 in Prince Albert National Park, Saskatchewan, Canada. Lines represent 95% confidence intervals. ... 111
x
Liste des tableaux
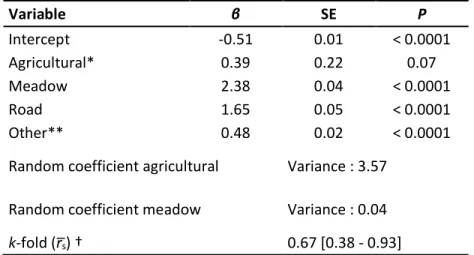
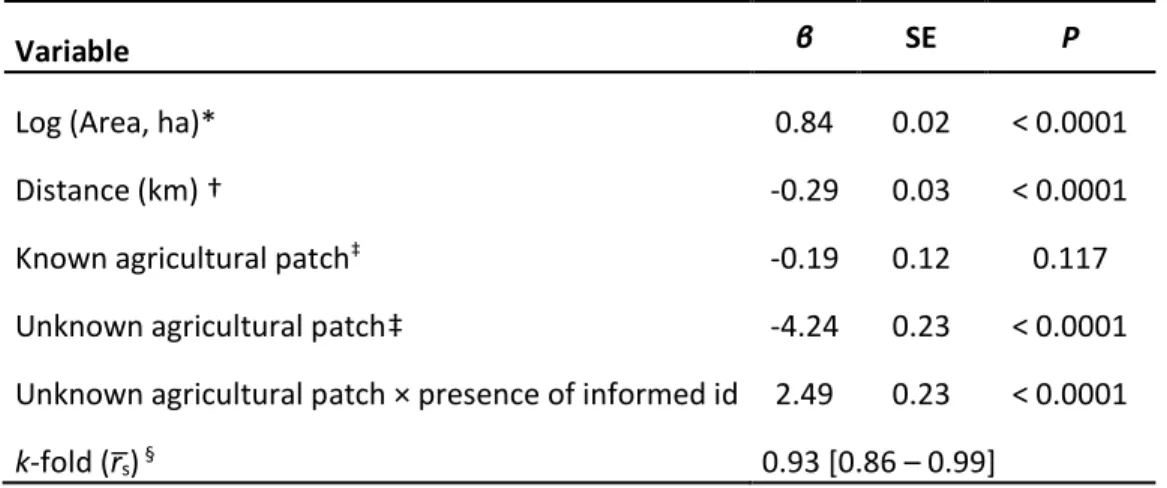
Table S1.1. Parameter estimates of a cox proportional hazards model describing the recurrences of visits by female plains bison to agricultural patches. The analysis was based on 18 GPS-collared female bison from August-October 2011 and 2012 representing 404 visits to agricultural patches near Prince Albert National Park, Saskatchewan (Canada). Model coefficients (β) are presented with hazard ratios, associated robust standard errors (SE), and p-values. A hazard ratio (exp[β]) less than one indicates that the instantaneous chance of recurrence decreases following an increase in the variable. ... 48 Table S1.2. Plant species preferentially consumed on meadows and agricultural lands. Ranking matrix of plant selection by plains bison on agricultural patches near Prince Albert National Park, Saskatchewan (Canada) during autumn (mid-August to early November) 2011 and 2012. Rankings based on comparing consumed and available green vegetation biomass in each agricultural patch (n = 26). A positive sign indicates the plant species in a row is preferentially consumed over the corresponding plant species in a column. A triple sign represents a significant difference (P < 0.05). ... 49 Table S1.3. Profitability of the most common plant species. Average profitability and associated standard errors (SE) for the six most common plant species on agricultural lands near Prince Albert National Park, Saskatchewan (Canada), together with Carex
atherodes, the most profitable plant species for plains bison in meadows within the
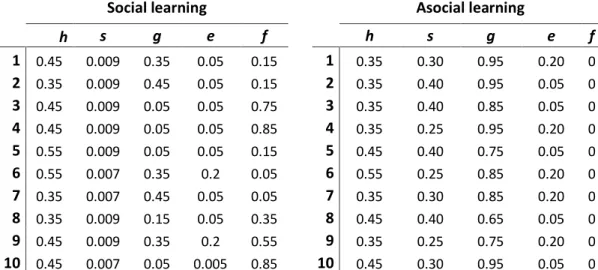
park. ... 50 Table S1.4. Habitat selection by bison. Parameter estimates of a mixed-effect Resource Selection Function (RSF) characterizing habitat selection by female plains bison (n = 40) in and around Prince Albert National Park, Saskatchewan (Canada) between July-October, from 2005 to 2013. Selection coefficients (β) are presented with standard errors (SE) and associated p-values, together with variance estimates of random effects. ... 51 Table S1.5. Factors influencing which patch a bison decides to visit next. Parameter estimates for the patch selection model of female plains bison (n = 22) in and around Prince Albert National Park, Saskatchewan (Canada), 2006 to 2013. Each variable is presented with the estimated coefficient (β), standard errors (SE) and associated p-values... 52 Table S1.6. Parameters values of the top-ranking models of information flow with (f > 0) and without social learning (f = 0). Models were based on the structure of a Susceptible-Infected-Recovered (SIR) model, where h is heightened mortality due to harvesting of emigrants outside Prince Albert National Park, Saskatchewan (Canada),
s represents the proportion of naïve individuals that wander out of the park as a result
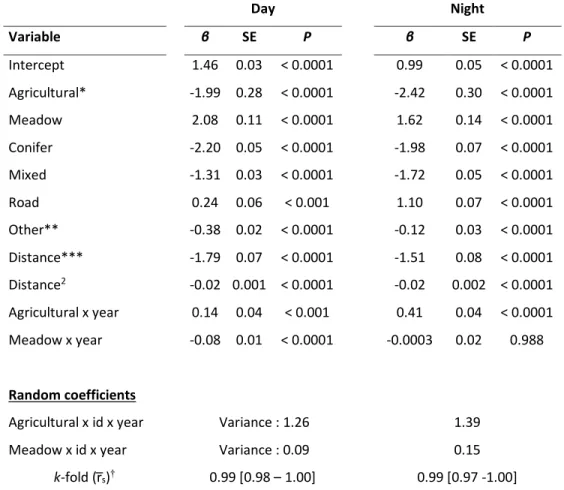
of exploratory movement behavior, g is the rate at which informed individuals leave the park, and e is the rate at which emigrants transfer back into the park. ... 53 Table 2.1. Parameter estimates of a mixed-effect Resource Selection Function (RSF) characterizing habitat selection by female plains bison (n = 46) in and around Prince Albert National Park, Saskatchewan (Canada) between July-October, from 2005 to 2015. Separate models were generated for each time period. Selection coefficients (β) are presented with standard errors (SE) and associated P-values, together with variance estimates of random effects. ... 69
xi
Table 2.2. Parameter estimates of a mixed-effect Resource Selection Function (RSF) characterizing habitat selection by wolves (n = 14) in and around Prince Albert National Park, Saskatchewan (Canada) between July-October, from 2007 to 2015. Separate models were generated for each time period. Selection coefficients (β) are presented with standard errors (SE) and associated P-values, together with variance estimates of random effects. ... 70 Table 2.3. Parameter estimates of a mixed-effect Step Selection Function (SSF) characterizing habitat selection by female plains bison (n = 32) in and around Prince Albert National Park, Saskatchewan (Canada) between July-October, from 2007 to 2015. Separate models were generated for each time period. Selection coefficients (β) are presented with standard errors (SE) and associated p-values, together with variance estimates of random effects. ... 71 Table S2.1. Parameter estimates of a mixed-effect Resource Selection Function (RSF) characterizing habitat selection by female plains bison (n = 46) in and around Prince Albert National Park, Saskatchewan (Canada) between July-October, from 2005 to 2015 including both time periods (day and night). Selection coefficients (β) are presented with standard errors (SE) and associated p-values, together with variance estimates of random effects. ... 75 Table S2.2. Parameter estimates of a mixed-effect linear regression characterizing distance travelled per hour by female plains bison (n = 46) in and around Prince Albert National Park, Saskatchewan (Canada) between July-October, from 2005 to 2015. Selection coefficients (β) are presented with standard errors (SE) and associated 95% confidence intervals (CI). ... 76 Table 4.1. Parameter estimates of the top-ranking logistic regressions characterizing the effects of the land cover type (reference category being deciduous forest), distance to nearest introduction point and utilization of 18 collared bison on the probability of clover and timothy occurrence along bison trails and around wallows in Prince Albert National Park, Saskatchewan, Canada, during autumns 2012 and 2013. Each model covariates is presented with estimated coefficients (β) and 95% confidence intervals (95% CI). ... 103 Table S4.1. Candidate models describing the probability of occurrence of exotic plant species along bison trails (i.e., clover and timothy) in Prince Albert National Park, Saskatchewan, Canada. The number of parameters (K) is presented together with deviance, AIC, and difference in AIC with respect to the model with the most empirical support (∆AIC). ... 107 Table S4.2. Identification of seeds recovered from 238 bison fecal samples collected during autumn 2012 in Prince Albert National Park, Saskatchewan, Canada. ... 108
xii
À mes parents, À Georges,
xiii
« Est-ce que nous ne sommes vraiment plus capables de respecter la nature, la liberté vivante, sans aucun rendement, sans utilité, sans autre objet que de se laisser entrevoir de temps en temps? »
xiv
Remerciements
Je souhaite remercier en premier lieu mon directeur de thèse Daniel Fortin, pour m’avoir offert l’opportunité de faire cette thèse et pour m’avoir encadré tout au long de ce travail. Les membres de mon comité d’encadrement, à savoir Jean-Pierre Tremblay et André Desrochers, ont également contribué à nourrir ma réflexion, et je les ai en remercie.
Je remercie également tous les agents de Parcs Canada qui ont contribué de près ou de loin à la réalisation de ce travail de thèse et ont permis que le terrain se passe dans les meilleures conditions. D’abord, Seth Cherry pour son appui technique mais également pour son travail en tant que co-auteur sur la plupart de mes chapitres ; Joanne Reimer et Glenn Rupert pour leur appui au quotidien pendant mes mois de terrain sur la « West Side » ; mais aussi Dustin Guedo et Jeff Weir pour leur travail sur les prairies à fétuques et les discussions à ce sujet.
Un grand merci à tous les agriculteurs du pourtour du parc pour m’avoir autorisé à collecter des données sur leurs terres mais également pour toutes ces discussions entre 2 portières de pick-up. Je voudrais également saluer le travail des Bison stewards et en particulier celui de Gordon Vaadeland et de Bettie Gillespie.
Une pensée également pour toutes les personnes croisées sur le terrain appartenant aux communautés des premières nations. Même si nos visions divergeaient souvent sur le devenir de la population des bisons du parc, au final, nous cherchons la même chose à savoir la prospérité des bisons.
Bien sûr je ne pouvais pas écrire ces remerciements sans une pensée pour Richard et Cherryl Crashley sans qui mes séjours en Saskatchewan n’auraient pas été les mêmes. Merci pour tous ces moments partagés et votre incroyable bienveillance à mon égard.
Cette thèse repose sur un important travail de terrain que ce soit dans le suivi des bisons, la collecte de données de végétation, les analyses de laboratoires ou encore l’identification de graines. Tout ceci aurait été impossible sans le travail précieux de
xv
mes assistants de recherche. Un grand merci à Nicolas Trudel, qui m’a mis le pied à l’étrier et qui grâce à son aisance en forêt boréale m’a permis de mener mon premier terrain plus sereinement ; à Victorien Valette pour son travail précis et dédié sur les données de végétation, à Chloé Bourson et Mélanie Marteau pour leur aide entre autres sur la récolte et l’identification de graines, et enfin Sylvia Pardonnet pour sa motivation et sa grande force de travail.
Je tiens aussi à remercier tous les membres du laboratoire de Daniel Fortin pour avoir contribué à une atmosphère de travail stimulante. Merci à Marie-Claude Labbé, Orphé Bichet, Chrystel Losier, Sabrina Courant, Léa Harvey, Jerod Merkle, Carolyn Nersesian, Qing Zhao, Mathieu Basille, Olivia Tardy, Julie Martineau, Caroline Gagné, Angélique Dupuch, Hélène le Borgne, Florian Barnier et Tom Mason.
Enfin, tout ceci n’aurait pas eu lieu sans le soutien sans faille de mes précieux parents, merci de toujours croire en moi et de me soutenir dans toutes mes aventures. Et pour terminer, bien sûr, merci Xavier, c’est bien simple sans toi je n’y serai jamais arrivée, cette thèse c’est aussi la tienne.
xvi
Avant-propos
Cette thèse de doctorat est présentée sous la forme de quatre articles scientifiques. Elle inclut également une introduction générale et une conclusion générale qui lient l’ensemble des articles, bien que chacun puisse être lu indépendamment. En tant qu’auteure principale de ces quatre articles, j’ai défini les objectifs de recherche, j’ai planifié, organisé et réalisé la collecte de la plupart des données. J’ai aussi effectué les analyses statistiques, les analyses en laboratoire et rédigé les manuscrits.
Mon chapitre 1 est publié dans Ecology Letters avec Daniel Fortin, Jerod Merkle, Seth Cherry, Andrew Berdahl et John Fryxell comme coauteurs. Le chapitre 2 sera soumis à Journal of Animal Ecology avec Daniel Fortin, Ricardo Simon et Seth Cherry comme coauteurs. Le chapitre 3 sera soumis sous forme de communication courte dans
Landscape Ecology avec Daniel Fortin comme coauteur. Le chapitre 4 est soumis à Animal Conservation avec Tom Mason, Florian Barnier, Seth Cherry et Daniel Fortin
comme coauteurs.
Au cours de mon doctorat, j’ai également contribué à 2 autres articles qui ne sont pas inclus dans la thèse. J’ai participé à la collecte des données, et à la révision du manuscrit de l’article de Fortin et coll. publié en 2014 dans Animal Production Science pour lequel je suis 3e auteure. J’ai participé à l’élaboration de la question de recherche
et à la révision du manuscrit pour l’article de Merkle et coll. publié en 2015 dans
1
Introduction
La pression sur les ressources naturelles est croissante face aux besoins d’une population humaine toujours plus importante. Pour certains chercheurs, nous sommes entrés dans une nouvelle phase de l’histoire, celle de l’anthropocène, pendant laquelle les activités humaines ont déclenché la 6e grande extinction d’espèces (Dirzo et al.
2014, Ceballos et al. 2015). L’anthropocène se caractérise, entre autres, par une forte régression des espaces disponibles pour la faune sauvage et par une importante progression des espèces non indigènes, deux causes majeures de l’érosion de la biodiversité (Vitousek et al. 1997, Fahrig 2003). La conversion rapide des habitats de la faune sauvage en milieu anthropisé pour faire place à de nouvelles activités humaines (p. ex., destruction d’habitats forestiers pour le développement agricole ou l’exploitation minière) crée de plus en plus d’interfaces souvent extrêmes entre milieux naturels et milieux anthropisés. Ainsi, dans les massifs forestiers sujets à l’exploitation forestière, on voit apparaitre de grandes zones de coupes qui présentent des caractéristiques drastiquement différentes de la forêt originelle (Bichet et al. 2016). Les conséquences pour les espèces résidentes de la forêt aussi bien dans ces zones de coupes qu’à leurs abords sont multiples. Les stratégies comportementales normalement valables dans leur milieu naturel peuvent ne plus être rentables dans ces milieux fortement perturbés (Schlaepfer et al. 2002). Ces interfaces entre la faune sauvage et les activités humaines sont également génératrices de conflits. Alors que leurs espaces se réduisent, les animaux utilisent de plus en plus les milieux anthropisés. Nombreux sont les exemples d’animaux qui utilisent des paysages agricoles et consomment des espèces végétales domestiques. Par exemple, les sangliers (Sus scrofa) en Europe se nourrissent très souvent sur des zones agricoles (Schley and Roper 2003). Les conséquences de ces conflits sont souvent dramatiques pour les espèces de la faune. Dans les milieux anthropisés, les animaux de la faune sauvage se retrouvent également en contact avec des nouvelles espèces animales ou végétales, souvent domestiquées. Ces nouvelles interactions interspécifiques peuvent avoir des répercussions sur la transmission d’agents pathogènes (Gogan et al. 2013) ou sur la dissémination d’espèces exotiques (Vellend et al. 2003). Comprendre le comportement des animaux qui vivent à l’interface entre paysages naturels et perturbés, doit permettre d’envisager les
2
conséquences de l’utilisation des milieux anthropisés par la faune sauvage et de mettre en place des mesures de gestion adaptées pour faciliter leur persistance à long terme.
Dans cette introduction, je développe dans un premier temps le cadre théorique pour comprendre les mécanismes qui influencent la sélection de l’habitat, et j’insiste sur un cas particulier : le piège écologique. Deuxièmement, je décris comment la coexistence entre des populations humaines et des espèces de la faune sauvage peut aboutir à des conflits, mais également au développement de stratégies comportementales complexes de la part des animaux. Troisièmement, je m’intéresse à l’utilisation des indices de connectivité et à la théorie des graphes pour étudier les déplacements des animaux, ainsi qu’à leur potentielle utilité dans une perspective de gestion des populations animales. Quatrièmement, je définis un conflit entre différents objectifs de conservation et j’explique en quoi leur prise en compte est négligée bien qu’essentielle dans une démarche de conservation à l’échelle de l’écosystème. Enfin, je termine en décrivant l’organisation de ma thèse et le système étudié.
1. Le choix de l’habitat
1.1. Les indices de la qualité de l’habitat
Choisir un habitat est un processus complexe qui est contraint par de nombreux paramètres liés aux caractéristiques mêmes de l’espèce telles que ses capacités cognitives, de déplacements, sa morphologie et les stratégies biodémographiques de l’espèce (Danchin et al. 2005). Les organismes utilisent alors des indices indirects reflétant la qualité de l’habitat qui sont présents dans leur environnement pour guider leur choix (Schlaepfer et al. 2002). Ces décisions sont adaptatives, car elles se basent sur des indices corrélés de manière fiable avec la survie et le succès reproducteur associés à l’habitat choisi (Williams and Nichols 1984). La sélection d’habitat est donc une conséquence de la sélection naturelle qui a favorisé les individus sélectionnant le meilleur habitat disponible (Fretwell and Calver 1969). Les indices utilisés varient grandement d’une espèce à l’autre. Certaines espèces se basent sur des caractéristiques physiques de l’habitat. Ainsi, le grand tétras (Tetrao urogallus) sélectionne les forêts ouvertes dominées par l’épicéa commun (Picea abies) et le sapin blanc (Abies alba)
3
(Sachot et al. 2003). Le gobemouche à collier (Ficedula albicollis) utilise, quant à lui, l’information publique pour évaluer la qualité des sites de reproduction disponibles en observant les déplacements associés aux soins parentaux de ses conspécifiques (Pärt and Doligez 2003). Le processus de sélection d’habitat devient plus complexe dans le cas d’animaux vivant en groupe qui doivent synchroniser leurs activités et leurs choix.
1.2. Décisions collectives : influence du groupe
La vie en groupe présente de nombreux avantages. Les membres d’un groupe bénéficient par exemple de la vigilance collective (Dehn 1990), de la coopération contre les prédateurs (Bednekoff and Lima 1998) et du partage d’informations (Fernandez-Juricic et al. 2006). Mais cela s’accompagne de certaines contraintes et les membres d’un même groupe doivent synchroniser leurs activités telles que le repos, la recherche de nourriture, et les déplacements à travers le paysage (Conradt and Roper 2000). Les décisions consensuelles favorisent l’option préférée par la majorité, et peuvent avoir un impact significatif sur l’utilisation de l’espace par les individus. Les membres d’un même groupe peuvent avoir une connaissance différente du paysage surtout dans les sociétés fission-fusion. Dans ces sociétés, les individus vont moduler la taille et la composition de leur groupe au travers de fissions temporaires et de la fusion de petites unités indépendantes en accord avec leur besoin et la répartition des ressources (Jacobs 2010). Certains individus peuvent être familiers de parcelles d’alimentation sur lesquelles d’autres membres du groupe ne se sont jamais rendus. Ces individus naïfs qui restent avec le groupe peuvent donc se retrouver sur une nouvelle parcelle simplement parce qu’ils ont suivi le choix de la majorité. Ils découvrent alors une nouvelle parcelle, et si cette parcelle est de bonne qualité ils peuvent l’inclure dans leur panel de parcelles disponibles et ajuster leur utilisation de l’espace et sélection d’habitat en conséquence (Krebs and Inman 1992, Couzin and Krause 2003).
Les travaux sur la valeur adaptative des décisions collectives suggèrent que celles-ci sont en général bénéfiques pour les membres du groupe (Conradt toper 2003, Franks 2003). Par exemple, en mettant en commun leur sens de l’orientation, les
4
membres d’un groupe peuvent identifier une direction de manière plus précise. Cette précision née de la somme des imprécisions individuelles — « The many wrong principle » — offre un bénéfice pour l’ensemble du groupe (Simons 2004). L’impact des décisions collective est généralement étudié soit dans des environnements strictement contrôlés en laboratoire (Franks et al. 2003, Miller et al. 2013) soit en milieu naturel non perturbé (Petit et al. 2009, Strandburg-Peshkin et al. 2015), mais peu de travaux se sont attachés à l’étude des ramifications d’un tel comportement dans des milieux transformés par les activités humaines.
1.3. Le piège écologique : émergence et sévérité
Puisque les animaux doivent évaluer la qualité de leur habitat indirectement en se basant sur des indices présents dans leur environnement, il est possible que la qualité perçue de l’habitat soit en fait déconnectée de sa qualité réelle, à savoir des bénéfices associés en termes de survie et de reproduction. Ceci peut se produire quand le comportement des animaux a été forgé par des conditions particulières, conditions qui changent soudainement (Levins 1968). Un piège écologique est le scénario qui se produit précisément quand des changements environnementaux soudains provoquent le découplage des indices utilisés et de la qualité de l’habitat (Robertson and Hutto 2006). La préférence de l’animal reste inchangée, mais le résultat attendu n’est plus le même et l’animal est contraint par son passé évolutif à faire une erreur alors que de meilleures options sont disponibles (Schlaepfer et al. 2002). Les concepts de pièges écologiques et de dynamique source-puit sont liés (Battin 2004, Gilroy et Sutherland 2007) dans le contexte où un puit qui est préféré à une source est souvent considéré comme un piège écologique (Patten et al. 2010).
Le premier piège écologique a été décrit par Dwernychuck et Boag (1972). Depuis, un certain nombre de pièges écologiques ont été rapportés dans la littérature, principalement chez les oiseaux (Schlaepfer et al. 2002). Mais c’est Roberston (2006) qui posa les bases théoriques permettant de rapporter l’existence d’un piège écologique de manière rigoureuse, et ce à l’aide de trois critères : (1) les individus doivent présenter une préférence égale ou supérieure pour un habitat par rapport à d’autres habitats
5
disponibles ; (2) une mesure valable de la valeur adaptative doit différer entre ces habitats ; (3) la valeur adaptative des individus qui ont choisi l’habitat préféré doit être inférieure à celles des individus dans les autres habitats disponibles.
Un large éventail d’activités humaines est à l’origine de pièges écologiques. L’introduction d’espèces exotiques (Schlaepfer et al. 2005) ou encore l’agriculture et la foresterie (Hallier and Gaertner 2008, Rodewald et al. 2011) sont des causes importantes d’émergence de pièges écologiques. Par exemple, les forêts exploitées de manière sélective peuvent présenter certaines caractéristiques communes avec des forêts ayant récemment brulées et qui sont sélectionnées par certaines espèces comme le passerin indigo (Passerina cyanea) sans pour autant forcément partager les mêmes ressources alimentaires ou risques de prédation équivalents (Weldon and Haddad 2005). Ces habitats peuvent alors se transformer en piège écologique (Robertson and Hutto 2007). De manière plus surprenante, les projets de restauration peuvent aussi avoir des conséquences inattendues et des mesures visant la restauration d’habitats peuvent également engendrer l’émergence de pièges écologiques (Robertson et al. 2013, Hale and Swearer 2017). Par exemple, une espèce rare de papillon (Lycaena
xanthoides) présente dans le sud de l’Oregon, dépose ces œufs plus fréquemment dans
les zones humides restaurées débarrassées des plantes exotiques envahissantes, mais qui sont inondables plutôt que dans les autres zones humides proches, mais non restaurées, provoquant une réduction importante de la survie des individus (Severns 2011).
Une hiérarchie peut être opérée quant au degré de sévérité du découplage entre les indices utilisés et la valeur adaptative, et donc la sévérité du piège écologique (Robertson et al. 2013). Cette distinction est pertinente quand on pense aux conséquences du piège écologique sur les populations animales. Les pièges sévères pouvant conduire au déclin rapide voire à l’extinction locale d’une population (Kokko and Sutherland 2001, Fletcher et al. 2012). On peut classer les pièges en fonction de leur degré d’attractivité. Un piège écologique qui est plus attractif que les autres habitats disponibles peut-être considéré comme un piège sévère en comparaison d’un piège qui est préféré de manière égale à d’autres habitats disponibles (Robertson and
6
Hutto 2006). D’autres facteurs peuvent également contribuer à la sévérité d’un piège écologique. Dans les groupes sociaux, les réponses individuelles sont souvent influencées par les réponses des autres (Galef and Laland 2005). Des individus peuvent alors se retrouver piégés en suivant d’autres individus déjà pris au piège. Or la probabilité d’extinction d’une population augmente avec la fraction des individus de cette population prise dans un piège écologique (Kokko and Sutherland 2001, Fletcher et al. 2012). Pourtant la dimension sociale est rarement prise en cause dans l’étude du développement et de la sévérité des pièges écologiques.
2. Causes et conséquences de l’utilisation des milieux anthropisés
2.1. Nouveaux paysages, nouvelles ressources, nouvelles stratégies
La conversion des habitats de la faune sauvage pour répondre à une demande croissante de la population humaine fait souvent place à des paysages agricoles (Lambin et al. 2003, Tscharntke et al. 2005). L’apparition parfois brutale de ces nouveaux paysages peut aller de pair avec l’apparition de nouvelles ressources. Les paysages agricoles, par exemple, sont caractérisés par l’exploitation d’espèces végétales souvent non-indigènes présentant de bonnes qualités nutritives. Elles peuvent alors être exploitées par des espèces sauvages. C’est le cas notamment de certaines espèces d’oies nichant en Arctique (Anser albifrons flavirostris et Anser
brachyrhynchus) qui ont vu leur population augmenter exponentiellement depuis les
années 50 suite à l’utilisation des parcelles agricoles qui offrent une ressource alimentaire abondante avant leur départ vers leur site de reproduction (Fox et al. 2005). La conservation de la biodiversité est de plus en plus tournée vers la protection des espèces dans des paysages anthropisés hors des zones protégées traditionnelles (Ahlering et al. 2013). Certains auteurs reconnaissent le potentiel croissant des paysages agricoles et agroforestiers pour la préservation de la biodiversité (Estrada et al. 2012). Mais la conservation des espèces hors des espaces qui leur sont dédiés demande une compréhension fine de l’impact des activités humaines sur ces espèces.
7
2.2. Paysage de la peur et les multiples sources de risque
Une des conséquences les plus évidentes de l’utilisation des milieux anthropisés est l’augmentation du risque de mortalité comme les mortalités dues aux collisions avec des véhicules ou à la chasse (Collins and Kays 2011). De nombreux travaux de recherche ont caractérisé l’impact non létal des perturbations humaines sur le comportement et le succès reproducteur des animaux (Frid and Dill 2002). Certains auteurs considèrent que les perturbations anthropiques peuvent affecter indirectement la valeur adaptative et la dynamique de population, au travers des coûts énergétiques supplémentaires (Regel and Pütz 1997), d’opportunités manquées dues au dérangement (Williams et al. 2006) ou encore de coûts physiologiques tels que le stress (Zwijacz-Kozica et al. 2013) de manière similaire au risque de prédation.
Alors que la grande majorité des proies font souvent face à plusieurs espèces de prédateurs (Sih et al. 1998, Lima 2002), la plupart des études empiriques et théoriques se concentrent encore sur des systèmes simplistes composés d’une seule proie et d’un seul prédateur (Lone et al. 2014). Pourtant, différents types de prédateurs du fait de leurs caractéristiques propres peuvent présenter des formes de risques variées pour leurs proies. Ainsi, un prédateur dit mobile comme le loup gris (Canis lupus), qui est constamment en déplacement, produira des indices de présence très variables et provoquera moins certainement une réaction comportementale chronique qu’un prédateur comme le puma (Puma concolor) qui présente un mode de chasse dit à l’affut (sit-and-wait) et des indices de présence plus persistants (Schmitz 2008, Thaker et al. 2011).
2.3. Plasticité comportementale
Les espèces de la faune doivent faire un compromis entre sélectionner des habitats qui offrent des ressources et éviter le risque de prédation ou les activités humaines, là où ces activités sont associées à une mortalité plus importante (Frid and Dill 2002, Darimont et al. 2015). La plasticité comportementale est considérée comme un des mécanismes majeurs grâce auxquels les animaux font face aux défis des nouveaux environnements (Dingemanse and Wolf 2013). Quand les animaux qui
8
cherchent à s’alimenter sont exposés à un prédateur potentiel, ils peuvent ajuster leurs patrons d’approvisionnement au risque perçu basé sur leur expérience passée. Ils peuvent même développer une nouvelle tactique anti-prédation pour continuer à s’approvisionner dans de bonnes conditions (Sol et al. 2013). Un des mécanismes majeurs derrière la plasticité comportementale est l’apprentissage, soit l’acquisition d’une nouvelle information qui influence l’expression d’un comportement (Dukas 2004). Au lieu d’exprimer constamment le même comportement face à un stimulus particulier, l’apprentissage permet aux animaux d’améliorer leur réaction comportementale sur la base de leur expérience passée (Dall et al. 2005). Certains animaux peuvent alors réagir aux dérangements liés aux activités humaines en déplaçant leurs activités dans l’espace (Foster et al. 2010) ou en modifiant leurs patrons d’activités dans le temps (Tigas et al. 2002).
2.4. Conflits homme-faune sauvage
La présence de la faune sauvage à proximité des activités humaines peut créer des conflits, et forcer les gestionnaires à prendre des mesures pour minimiser ces conflits (Ditchkoff et al. 2006). Ces conflits sont en augmentation (Hill 1997, Conover 2002, Naughton-Treves et al. 2003, Campbell-Smith et al. 2011) précisément à cause de la réduction des espaces disponibles. L’exploitation de ressources alimentaires présentes dans les paysages anthropisés, mais absentes des paysages naturels en est souvent à l’origine. Les cultures proches des forêts sont des ressources prévisibles et accessibles pour la faune sauvage. Les dommages sur les cultures causés par la faune sauvage peuvent impacter négativement les moyens de subsistance des fermiers et se traduir par des pertes économiques importantes (Nyhus et al. 2005). Le cerf de Virginie à lui seul provoque plusieurs millions de dollars de pertes par an liées uniquement à la déprédation sur la culture du maïs (Delger et al. 2011). Ces dommages peuvent également réduire la tolérance vis-à-vis de la faune sauvage et ultimement miner les stratégies de gestion et de conservation (Wallace and Hill 2012). Par exemple, plus d’éléphants meurent tués au cours de conflits liés à la destruction des cultures que par n’importe quelle autre source de mortalité une fois adulte, et ce dans tous les pays où le braconnage est maîtrisé (Western and Waithaka 2005). Les efforts de gestion qui
9
visent à minimiser les conflits entre les hommes et les espèces de la faune sauvage ont besoin d’une compréhension détaillée des stratégies comportementales mises en place par les animaux en réaction aux activités humaines. Influencer les déplacements des animaux dans le but de modifier leur utilisation de l’espace est souvent envisagé dans les programmes de gestion.
3. Les réseaux spatiaux et leur utilisation comme outil de gestion
3.1. Connectivité et répartition spatiale
La connectivité est reconnue comme un élément essentiel à prendre en compte dans l’étude de la répartition des espèces et la conservation de la biodiversité (Taylor et al. 1993, Bunn et al. 2000), même si son importance relative par rapport à d’autres attributs de l’habitat est discutée (Doerr et al. 2011, Hodgson et al. 2011). Elle se réfère au degré par lequel un paysage facilite ou empêche le mouvement d’un organisme entre des parcelles d’habitat (Taylor et al. 1993). En effet, la capacité d’une espèce à se déplacer entre les parcelles d’habitat dans un milieu hétérogène peut influencer directement le succès d’approvisionnement ou de dispersion de cette espèce ou de celles qui lui sont inféodées, et ainsi influencer leur répartition spatiale et la dynamique des populations (Goodwin 2003).
La connectivité entre des parcelles peut être définie sur des bases structurelles ou fonctionnelles (Tischendorf and Fahrig 2000a, Crooks and Sanjayan 2006, Taylor et al. 2006). La connectivité structurelle considère l’agrégation spatiale des parcelles et inclut donc la grandeur des parcelles et la distance qui les sépare. Elle est mesurée en analysant la structure du paysage sans prendre en compte les mouvements de l’organisme à travers le paysage (Crooks and Sanjayan 2006). La connectivité fonctionnelle implique la manière dont le paysage facilite ou gêne les déplacements des espèces entre les parcelles de ressources (Taylor et al. 1993, Crooks and Sanjayan 2006). Elle est propre à chaque espèce et dépend de la façon dont l’organisme perçoit la structure du paysage, de ses capacités cognitives, de déplacement et de ses motivations (Bunn et al. 2000, Tischendorf and Fahrig 2000b, Bélisle 2005). Les travaux récents menés chez le bison sur la connectivité fonctionnelle se sont intéressés,
10
entre autres, aux caractéristiques du paysage influençant le choix de la prochaine parcelle visitée (Dancose et al. 2011). Mais le déplacement des bisons à l’échelle du paysage au travers d’un ensemble de parcelles connectées les unes aux autres reste encore à clarifier.
3.2. La théorie des graphes
Un graphe est constitué de nœuds et de liens, où les nœuds, dans le cas d’un réseau d’habitats, représentent les parcelles de ressources pour un animal, et les liens, les routes qui relient ces parcelles (Urban et al. 2009). L’utilisation spatiale des graphes, souvent nommés réseaux, permet l’étude d’un modèle simplifié du paysage réalisant un compromis entre les données biologiques et la complexité qu’il est possible de prendre en compte (Theobald 2006, Urban et al. 2009). La définition des liens et la manière dont le réseau est assemblé (c.-à-d., quels nœuds sont connectés) influencent fortement la structure du réseau (Galpern et al. 2011).
Les études qui utilisent les réseaux pour l’étude des déplacements des animaux ont souvent recourt pour des questions pratiques à une structure de réseau définie a
priori basée sur la connectivité structurelle (Galpern et al. 2011). Le graphe planaire
minimum (GPM), par exemple, est un type de réseau qui considère que les déplacements entre les parcelles d’intérêt sont restreints entre des nœuds adjacents et se font par le biais de « stepping stones », mais également qu’aucun lien ne se croise (Theobald 2006). L’utilisation du GPM est proposée comme présupposé dans certaines études de connectivité (Fall et al. 2007) bien que peu d’études empiriques aient montré l’existence d’une telle structure dans les réseaux utilisés par les animaux (Courbin et al. 2014). Pourtant, la structure d’un réseau a des implications importantes sur son fonctionnement (Estrada and Bodin 2008) et ses propriétés telle que la résistance (Fall et al. 2007) – soit sa capacité à se maintenir après une perturbation (Holling 1973) et à garantir la circulation des individus.
11
3.3. Influencer les patrons de déplacements : un outil de gestion
L’utilisation d’une représentation pertinente du réseau est nécessaire pour la mise en place de mesures de gestion efficaces et pour prédire comment les déplacements des individus vont être influencés suite à une perturbation. Si un réseau est particulièrement redondant, c’est-à-dire qu’il existe de nombreuses routes alternatives pour relier la même paire de nœuds (Rayfield et al. 2011), il va être plus difficile à perturber qu’un réseau dont les nœuds sont faiblement connectés. Ainsi une amélioration de la compréhension des conséquences écologiques de la connectivité des réseaux est nécessaire pour une meilleure prédiction des conséquences liées aux perturbations naturelles et anthropiques (Urban et al. 2009). Elle peut également être mise à profit pour cibler certaines interventions visant à réduire l’utilisation par une espèce animale d’une ressource d’intérêt pour l’homme (Fortin et al. 2010). Les grandes espèces de mammifères telles que les éléphants ou les bisons font souvent l’objet de mesures de gestion plus ou moins efficaces visant à leur interdire l’accès à certaines zones exploitées par l’homme (Meagher 1989a, King et al. 2009). Dans ces zones, les animaux sont susceptibles de rentrer en contact avec des espèces domestiques et peuvent contribuer à la propagation d’agents pathogènes ou d’espèces exotiques (Daszak et al. 2000).
4. Les conflits de conservation
4.1. Définition et émergence
Alors que de plus en plus d’espèces sont menacées, le nombre de programmes de conservation se multiplie pour essayer d’enrayer la crise de la biodiversité. Mais certaines mesures de gestion et de conservation qui visent à favoriser une espèce en particulier peuvent avoir des impacts négatifs sur d’autres efforts de conservation orientés vers d’autres espèces également menacées. Ces situations peuvent alors aboutir à un dilemme de conservation inextricable (Williams et al. 2011). Ce type de phénomène est plus susceptible de se produire dans des zones où les activités humaines ont fortement altéré les interactions entre les espèces (Sinclair and Byrom 2006). Ainsi, certaines actions de conservation, comme par exemple, l’éradication d’une espèce
12
envahissante ou la réintroduction d’espèces localement éteintes, peuvent avoir des conséquences inattendues et négatives sur d’autres taxa.
4.2. Le cas de la dissémination de plantes envahissantes dans des milieux sensibles par une espèce animale d’intérêt patrimonial
On considère les processus d’envahissement par des espèces animales et végétales introduites par l’homme comme une des causes majeures du déclin de la biodiversité (Clavero and García-Berthou 2005). Le caractère envahissant des espèces végétales non-indigènes dans de nouveaux habitats est fortement dépendant des patrons et des mécanismes de dispersion de graines. Les événements de dispersion à longue distance et les multiples points d’introduction peuvent augmenter le succès d’envahissement et contribuer à une large répartition spatiale des populations de plantes non-indigènes (Lockwood et al. 2007). La participation d’un vecteur animal est souvent nécessaire, notamment dans le cas particulier des espèces envahissantes (Vellend et al. 2003, Myers et al. 2004). Les animaux de grande taille favorisent la dispersion à longue distance du fait de leur grand domaine vital et de leur vitesse de déplacement, ainsi que du temps de rétention important lié à leur capacité digestive (Nathan et al. 2008). Les grands animaux agissent comme des vecteurs généralistes et transportent une grande variété de graines (Myers et al. 2004, Rosas et al. 2008). Les perturbations au niveau du microsite où les graines sont déposées peuvent favoriser la croissance de ces espèces aux dépens des autres espèces naturellement présentes (Seabloom et al. 2003, Torn et al. 2010). Un vecteur qui dépose des propagules d’une espèce donnée dans des zones perturbées peut être un agent de dispersion de grande valeur pour cette espèce (Schupp 1993, Wenny 2001).
Les grandes espèces de mammifères qui vivent à l’interface de milieux naturels et anthropisés peuvent être d’importants vecteurs de dissémination de plantes non-indigènes. Par exemple, le cerf de Virginie (Odocoileus virginianus) en Amérique du Nord contribue à la dispersion à longue distance de centaines d’espèces de plantes différentes (Myers et al. 2004). Leurs déplacements sur des distances importantes peuvent les mener à fréquenter à la fois des milieux naturels préservés et des zones
13
anthropisées riches en espèces végétales introduites, comme les zones cultivées. Un conflit de conservation peut alors émerger quand une espèce animale vulnérable à forte valeur patrimoniale ou en danger d’extinction se révèle être un vecteur préférentiel d’introduction d’espèces de plantes exotiques dans des milieux fragiles ou en cours de restauration.
5. Modèle d’étude : le bison des prairies
Dans le cadre de ce travail de doctorat, j’ai étudié une population de bisons des prairies (Bison bison bison) établie dans et autour du parc national de Prince Albert (PNPA, Canada). Les bisons sont parmi les plus grands herbivores existants en Amérique du Nord (Meagher 1989b). Avant l’arrivée des Européens, le nombre de bisons avoisinait les 60 000 millions, aujourd’hui ils sont classés comme espèce menacée selon la loi sur les espèces en péril du Canada (COSEWIC 2013). En Amérique du Nord, on dénombre actuellement plus de 500 000 bisons, mais seulement 5 % d’entre eux vivent au sein de populations vouées à la conservation de l’espèce à l’état naturel. La grande majorité étant entretenue par des entreprises privées à des fins commerciales, principalement pour l’élevage (Freese et al. 2007, Gates et al. 2010). La population de bisons des prairies du PNPA est l’une des rares populations en liberté (c.-à-d., ne se trouvant pas dans une zone clôturée) en Amérique du Nord et la seule qui vit dans son aire de répartition historique au Canada.
La population de bisons du PNPA n’a jamais dépassé les 500 individus, et a connu un déclin important depuis 2006 (Merkle et al. 2015a). En 2014, la population était composée d’environ 222 individus. Les bisons de PNPA sont une priorité de conservation pour Parcs Canada qui cherche à maintenir une population en liberté, autosuffisante (c.-à-d., sans apport de nourriture exogène) et qui se régule naturellement (Parks Canada 2013). Elle occupe une aire de répartition d’environ 1 000 km2 principalement composée de forêts. Les autres grands herbivores présents sur
le même territoire sont le wapiti (Cervus canadensis), le cerf de Virginie (Odocoileus
14
americanus), mais c’est surtout la présence sur ce même territoire du loup gris (Canis lupus) qui peut représenter une menace pour les bisons.
Les bisons sont grégaires et vivent au sein de groupes dont la dynamique est de type fusion-fission, c.-à-d. caractérisée par des changements fréquents dans leur composition (Lott and Minta 1983). Ainsi, la taille des groupes observés chez les bisons de PNPA varie de 3 à 150 individus et les événements de fission se produisent environ toutes les 21 h (Fortin et al. 2009). Cette dynamique de groupe a une influence notable sur le comportement d’approvisionnement et l’utilisation de l’espace par le bison (Courant and Fortin 2010, Merkle et al. 2015b).
Chez le bison des prairies, la connectivité fonctionnelle a été étudiée à l’échelle du déplacement d’une parcelle vers une autre (Dancose et al. 2011). Mais l’intégration à l’échelle du paysage et l’étude de la connectivité au niveau de l’ensemble du réseau de parcelles disponibles reste à explorer. Pourtant nous nous trouvons dans un système particulièrement adapté étant donné que les bisons se déplacent naturellement au sein d’un réseau de parcelles enchâssées dans une matrice de forêt. Les bisons se déplacent au sein d’un large réseau de sentiers qui sont créés par le passage répété de groupe de bisons. Les sentiers de bisons sont faciles à identifier, car ce sont des zones bien visibles, larges et dénuées de végétation (McHugh 1958).
Les études menées chez le bison montrent qu’il peut jouer un rôle dans la dispersion d’espèces végétales par endozoochorie et épizoochorie (Constible et al. 2005, Rosas et al. 2008). De plus, les bisons créent des dépressions dans le sol (« wallow ») suite à l’utilisation répétée de certains sites pour se rouler dans la terre (Coppedge and Shaw 2000), ce qui crée des microsites au sein desquels la végétation indigène est perturbée. Les caractéristiques physiologiques [temps moyen de rétention dans le système digestif de 78-79 h (Schaefer et al. 1978)] et comportementales (déplacements entre milieu naturel et anthropisé, créations de « wallows ») du bison des prairies en font un vecteur potentiellement important pour la dissémination d’espèces envahissantes au sein du PNPA.
15
Cette population de bisons offre l’occasion d’étudier les causes et les conséquences de l’utilisation des milieux anthropisés dans un paysage partagé en aire protégée et terres agricoles. En effet, les grands mammifères sont souvent au centre de problématiques conflictuelles liées au partage des espaces du fait de la grande taille de leurs domaines vitaux et des dégâts qu’ils peuvent engendrer sur les terres privées (Kilpatrick et al. 2009, Webber et al. 2011). L’utilisation de l’espace par les bisons des prairies est marquée par une forte saisonnalité qui les mène souvent à l’extérieur des territoires qui leur sont alloués (Meagher 1989b, Shaw and Carter 1990). Ce phénomène également observé au sein du PNPA est en augmentation depuis les années 90 et aboutit au développement de conflits d’usage dans la mesure où le PNPA est bordé par des terres agricoles exploitées (Falardeau Alain 2007, Duchesne et al. 2010). Quand ils se trouvent à l’extérieur du parc, les bisons peuvent être chassés par les populations autochtones dans le cadre de la chasse de subsistance. Les chasseurs doivent au préalable obtenir l’autorisation des propriétaires terriens pour chasser sur leur terre mais n’ont aucune obligation de rapporter le nombre de bisons abattus. Il n’y pas de quota de chasse et le nombre de bisons abattus chaque année est laissé à la discrétion des chasseurs.
6. Objectif et organisation de la thèse
Ce travail de doctorat vise une meilleure compréhension des mécanismes poussant les espèces sauvages à fréquenter les milieux anthropisés, et à leurs conséquences à la fois sur le comportement et la valeur adaptative de ces espèces, mais aussi sur l’intégrité des écosystèmes dans des paysages de plus en plus perturbés par les activités humaines. Elle s’intéresse également aux déplacements des bisons par le prisme du réseau de sentiers connectant les prairies naturelles. Elle interroge alors la pertinence de gêner les bisons dans leurs déplacements pour réduire leur utilisation des parcelles agricoles. Cette thèse comprend 4 chapitres, pour lesquels je propose ci-dessous un résumé de leur importance scientifique et de leurs objectifs.
16
Chapitre 1.
Les bénéfices du partage d’informations entre individus ont largement contribué à l’évolution de la vie en groupe. Pour que les animaux tirent avantage de l’information sociale, leurs congénères doivent adopter des comportements appropriés. Or, avec les modifications de l’environnement par les activités humaines, les animaux peuvent être attirés par un habitat anthropisé qui leur semble de haute qualité, mais qui ne l’est pas. Sous ces nouvelles contraintes, les décisions consensuelles au sein d’un groupe, généralement considérées comme bénéfiques, peuvent se révéler délétères et réduire la valeur adaptative des individus. Les objectifs de ce chapitre sont 1) d’évaluer si les parcelles agricoles constituent un piège écologique et 2) de déterminer si le transfert d’information entre les individus peut expliquer la diffusion rapide du comportement de sélection des parcelles agricoles au sein de la population de bisons.
Chapitre 2.
En milieu naturel, les proies font rarement face à un seul type de prédateurs, mais plutôt à une communauté de prédateurs. Face à ces menaces multiples, les proies doivent mettre en place différentes stratégies qui correspondent à différents types de prédateurs. Dans les milieux anthropisés, le risque de prédation peut englober à la fois les prédateurs naturels, mais également la menace assimilée aux activités humaines. Comprendre comment ces espèces ajustent leur comportement spatial et temporel pour minimiser le risque lié aux activités humaines tout en continuant de prêter attention à leurs prédateurs naturels, nous permet de mieux anticiper comment les espèces proies échappent au risque de prédation dans les milieux anthropisés. Les objectifs de ce chapitre sont 1) d’évaluer la stratégie comportementale mise en place par les bisons pour minimiser le risque de prédation représenté par le loup gris ; 2) d’étudier le comportement de sélection des prairies naturelles par le loup gris et 3) de mesurer l’ajustement du comportement de sélection des parcelles agricoles pour éviter les activités humaines.
17
Chapitre 3.
L’utilisation des réseaux spatiaux est un outil puissant pour étudier les déplacements des animaux. La résilience aux perturbations et la connectivité d’un réseau sont des propriétés qui dépendent directement de la structure du réseau. Les objectifs de ce chapitre sont 1) d’étudier la structure du réseau utilisé par les bisons pour se déplacer entre des parcelles d’alimentation (c.-à-d., prairies naturelles) au sein d’une matrice forestière ; 2) de comparer le réseau observé à un modèle théorique couramment utilisé, le graphe planaire minimum (GPM) et 3) de comparer le réseau à deux périodes caractérisées par une diminution importante de la taille de la population de bisons.
Chapitre 4.
L’émergence de conflits entre différents objectifs de conservation est une problématique grandissante et souvent sous-estimée par les gestionnaires. Pourtant avec la multiplication des programmes de conservation pour faire face au déclin de la biodiversité, il est attendu que certains enjeux de conservation visant une espèce bien particulière puissent entrer en conflit avec d’autres programmes de conservations centrés sur d’autres espèces d’intérêt. Identifier les mécanismes d’émergence de ces conflits et quantifier leurs impacts potentiels apparaît nécessaire pour améliorer la conservation de l’écosystème. Les objectifs de ce chapitre sont d’étudier 1) la dispersion des graines d’espèces de plantes non-indigènes par le bison et 2) l’impact potentiel de la dispersion de ces graines par le bison pour les prairies de fétuques (Festuca hallii) en cours de restauration dans le parc national de Prince Albert.
18
Chapitre 1. Collective decision-making promotes
fitness loss in a fusion-fission society
Marie Sigaud1, Jerod A. Merkle1,2, Seth G. Cherry3, John M. Fryxell4, Andrew Berdahl5, and Daniel Fortin1
1 Département de Biologie et Centre d’Étude de la Forêt, Université Laval, Québec,
Canada.
2 Wyoming Cooperative Fish and Wildlife Research Unit, University of Wyoming,
Laramie, USA.
3 Parks Canada Agency, Radium Hot Springs, British Columbia, Canada.
4Department of Integrative Biology, University of Guelph, Guelph, Ontario, Canada. 5Santa Fe Institute, Santa Fe, New Mexico, USA
© Reproduction autorisée de l'article : Sigaud, M., Merkle, J.A., Cherry, S.G., Fryxell, J.M., Berdahl, A. & Fortin, D. (2017) Collective decision-making promotes fitness loss in a fusion-fission society. Ecology Letters, 20, 33-40.
19
Résumé
Alors que les décisions collectives sont reconnues comme contribuant de manière significative à la valeur adaptative des espèces sociales, le résultat opposé est logiquement également possible. Nous montrons que les décisions collectives de déplacements qui sont guidées par des individus qui partagent une information erronée sur la qualité de l’habitat promeuvent l’utilisation d’un piège écologique. Les associations courtes, mais fréquentes entre bisons qui possèdent des connaissances spatiales différentes mènent à un basculement de leur comportement au départ d’évitement puis vers une sélection des parcelles agricoles au cours d’une période de 9 ans, dans et autour du Parc National de Prince Albert, Canada. Les bisons sont plus susceptibles de se rendre sur une parcelle agricole pour la première fois en suivant un de leurs congénères déjà familiers avec les parcelles agricoles. La mortalité annuelle a augmenté de 12% du fait de la chasse sur les parcelles agricoles. Ce comportement social mésadapté a été une force majeure dans le déclin de la population de 50% en moins d’une décennie. Dans les paysages altérés par les activités humaines, l’apprentissage social par les espèces qui vivent en groupes peut mener à une diminution de la valeur adaptative, en particulier dans les sociétés fission-fusion.
20
Abstract
While collective decision-making is recognized as a significant contributor to fitness in social species, the opposite outcome is also logically possible. We show that collective movement decisions guided by individual bison sharing faulty information about habitat quality promoted the use of ecological traps. The frequent, but short-lived, associations of bison with different spatial knowledge led to a population-wide shift from avoidance to selection of agricultural patches over nine years in and around Prince Albert National Park, Canada. Bison were more likely to travel to an agricultural patch for the first time by following conspecifics already familiar with agricultural patches. Annual adult mortality increased by 12% due to hunting of bison on agricultural lands. Maladaptive social behavior accordingly was a major force that contributed to a 50% population decline in less than a decade. In human-altered landscapes, social learning by group-living species can lead to fitness losses, particularly in fusion-fission societies.