RELATIONS ALLOMETRIQUES DE L'ÉPINETTE NOIRE (PICEA
MARIANA(MILL.) B.S.P.) ET DE L'ÉPINETTE BLANCHE (PJCEA GLAUCA
(MOENCH)
VOSS)
THÈSE
PRÉSENTÉE
COMME EXIGENCE PARTIELLE
DU DOCTORAT EN BIOLOGIE
PAR
HUGUES POWER
UNIVERSITÉ DU QUÉBEC À MONTRÉAL Service des bibliothèques
Avertissement
La diffusion de cette thèse se fait dans le respect des droits de son auteur, qui a signé le formulaire Autorisation de reproduire et de diffuser un travail de recherche de cycles supérieurs (SDU-522 - Rév.01-2006). Cette autorisation stipule que «conformément à l'article 11 du Règlement no 8 des études de cycles supérieurs, [l'auteur] concède
à
l'Université du Québecà
Montréal une licence non exclusive d'utilisation et de publication de la totalité ou d'une partie importante de [son] travail de recherche pour des fins pédagogiques et non commerciales. Plus précisément, [l'auteur] autorise l'Université du Québec à Montréal à reproduire, diffuser, prêter, distribuer ou vendre des copies de [son] travail de rechercheà
des fins non commerciales sur quelque support que ce soit, y compris l'lntern~t. Cette licence et cette autorisation n'entraînent pas une renonciation de [la] part [de l'auteur]à
[ses] droits moraux nià
[ses] droits de propriété intellectuelle. Sauf entente contraire, [l'auteur] conserve la liberté de diffuser et de commercialiser ou non ce travail dont [il] possède un exemplaire.»Le travail de la thèse a été réalisé sous la supervision et avec la collaboration de mon directeur et de mes co-directeurs. J'ai planifié, supervisé et exécuté la prise de donnée sur les sites du Lac-St-Jean et du Témiscamingue. J'ai participé à la prise de donnée sur le site de la forêt de recherche de Petawawa. La planification et le choix des peuplements et des arbres à échantillonner à la forêt de recherche de Petawawa a été effectué par l'étudiant à la maîtrise Adam Kuprivicius sous la supervision du Dr. John Caspersen de l'Université de Toronto. Les données provenant de l'Alberta proviennent des travaux de doctorat de Derek F. Sattler, travaux effectués sous la supervision du Dr. Phil Carneau à l'Université de l'Alberta.
La thèse, qui compte trois chapitres, est présentée sous forme d'articles scientifiques rédigés en anglais. Le premier chapitre qui porte sur la comparaison des dimensions de la cime vivante de l'épinette noire et de l'épinette blanche a été publié dans la Revue canadienne de la recherche forestière sous le titre: Differences in crown characteristics between black (Picea mariana) and white spruce (Picea glauca). Les deuxièmes et troisièmes chapitres portant respectivement sur la comparaison de la biomasse foliaire des deux espèces et sur le modèle tubulaire de l'épinette blanche et de l'épinette noire seront soumis à des journaux avec révision par les pairs.
Cette thèse est le fruit de la collaboration avec différents chercheurs. Je tiens à remercier '
tout particulièrement mes deux co-directeurs les Drs. Frank Berninger et Valerie LeMay ainsi que mon directeur le Dr. Daniel Kneeshaw qui m'ont guidé tout au long de la réalisation de ce projet et qui ont fait preuve d'une grande disponibilité. Je voudrais également remercier le Dr. Robert Schneider pour sa fructueuse collaboration, Derek F. Sattler, la forêt de recherche de Petawawa et Adam Kuprivicius pour avoir rendu disponible leurs données permettant ainsi d'améliorer la pmtée du projet. Cette thèse n'aurait pu être réalisée sans le travail dévoué des étudiants chercheurs et des assistants de recherche qui ont participé au travail de terrain et à celui de laboratoire à un moment ou l'autre du projet.
l1l
Je tiens également à remercter, le Réseau stratégique CRSNG sur l'aménagement forestier pour les produits à valeur ajouté ainsi que le Fond québécois de recherche nature et technologie (FQRNT). Ces deux organismes ayant financé le projet.
Je voudrais finalement remercier ma famille mes amis et Marie-Eve pour le support qu'ils ont manifesté à un moment ou l'autre du projet.
AVANT-PROPOS ... ii
LISTE DES FIGURES ... ix
LISTE DES TABLEAUX ... x
LISTE DES ABRÉVIATIONS SIGLES ET ACRONYMES ... xi
RÉSUMÉ ... xiii
INTRODUCTION GÉNÉRALE ... 1
0.1 Relations allométriques et traits fonctionnels ... 1
0.2 Autoécologie de l'épinette noire et de l'épinette blanche ... 3
0.3 Implications pratiques de l'étude ... 6
0.4 Objectif général de la thèse ... 6
0.5 Objectifs spécifiques de la thèse ... 7
CHAPITRE I Differences in crown characteristics between black (Picea mariana)and white spruce (Picea glauca) ... 8
1.1 Résumé ... 9
1.2 Abstract ., ... 10
1.3 Introduction ... 11
1.4 Materials and methods ... 13
1.4.1 Data ... 13
1.4.2 Calculations of plot- and tree-level variables including imputations of rnissing heights ... 15
v
1.4.3 Crown reconstruction ... 17
1.4.4 Crown length ... 19
1.4.5 Crown profile ... 20
1.4.6 Crown surface area ... 21
1.5 Results ... 22
1.5.1 Crown reconstruction ... 22
1.5.2 Crown length models ... 22
1.5.3 Crown profile models ... 23
1.5.4 Crown surface area ... 24
1.6 Discussion ... 24
1.6.1 Crown length ... 24
1.6.2 Crown profile ... 25
1.6.3 Crown shape ... 26
1.6.4 Crown surface area ... 27
1.6.5 Constraints of the study ... 28
1.7 Conclusions ... 28
CHAPITRE II Comparison of three different foliage biomass characteristics of black (Picea mariana) and white spruce (Picea glauca) ... 38
2.1 Résumé ... 39
2.3 Introduction ... 41
2.4 Materials and methods ... : ... 4 3 2.4.1 Data ... 43
2.4.2 Foliage biomass analysis models ... 48
2.5 Results ... 52
2.5.1 Total foliage biomass ... 52
2.5.2 Foliage biomass density ... 53
2.5 .3 Cumulative relative foliage biomass vertical distribution ... 53
2.6 Discussion ... 54
2.6.1 Variation in foliage biomass and foliage biomass density ... 54
2.6.2 Relative cumulative foliage biomass vertical distribution ... 56
2.7 Conclusion ... 58
2.8 Appendix ... 65
2.8.1 Total height imputation ... 65
2.8.2 Branch foliage biomass estimation ... 65
2.8.3 Crown radii estimation ... 66
2.8.4 Crown surface area estimation ... 67
CHAPITRE III Differences in pipe model ratios between two spruce species, within individuals and across different growing conditions ... 69
Vll
3.2 Abstract ... 71
3.3 Introduction ... 72
3 .4 Materials and methods ... 7 4 3.4.1 Data ... 74
3.4.2 Calculations ofplot-level variables ... 77
3.4.3 Branch-level foliage biomass modelling and imputation ... 77
3.4.4 Tree-level pipe model ratio models ... 79
3 .4.5 Disk-level pipe model ratio models ... 80
3.5 Results ... 81
3.5 .1 Branch lev el foliage biomass ... 81
3.5.2 Tree-level pipe model ratio ... 82
3.5.3 Disk-level pipe model ratio ... 83
3.6 Discussion ... 85
3.6.1 Shade tolerance differences ... 85
3.6.2 Hydraulic support requirements ... 86
3.6.3 Mechanical support requirements ... 88
3.6.4 Originality and limitations of the study ... 89
3.7 Conclusions ... 89
CONCLUSION GÉNÉRALE ... 99
4.2 Limites de la thèse ... 102 4.3 Applications pratiques ... 103 RÉFÉRENCES ... 1 04
LISTE DES FIGURES
Figure 1.1 Relationship between horizontallength, branch length and branch angle ... 33
Figure 1.2 Crown length models ... 34
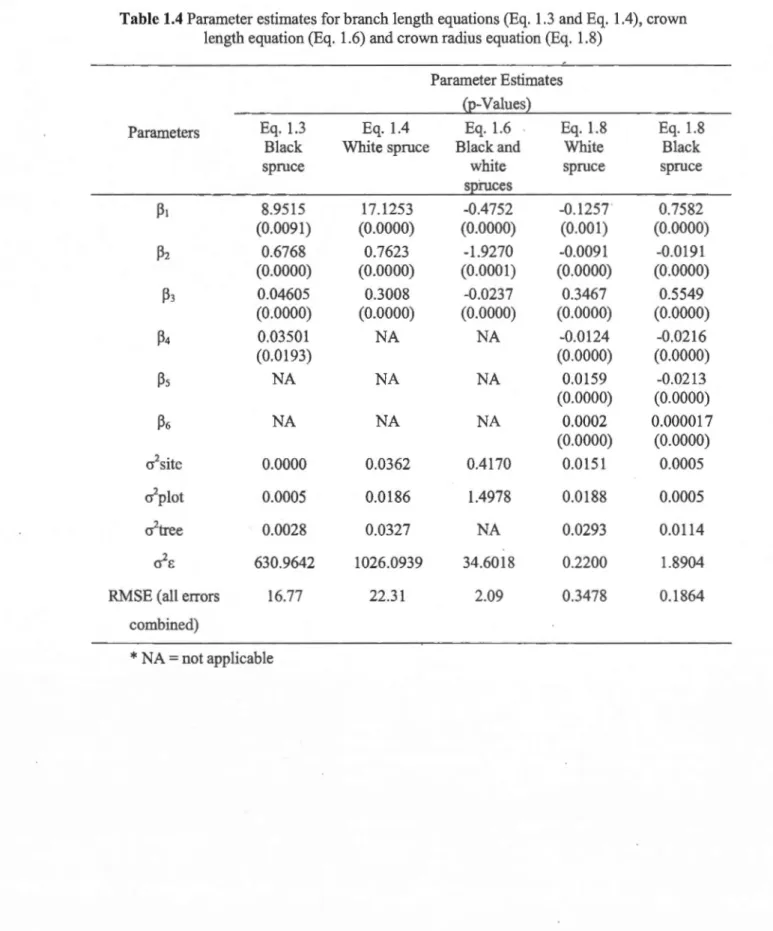
Figure 1.3 Crown radius over depth ... 35
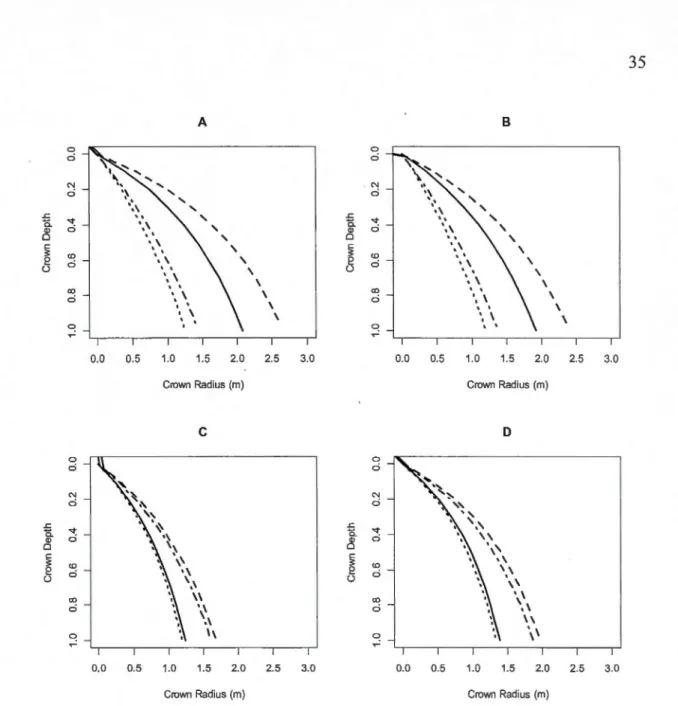
Figure 1.4 Relative crown radius over crown depth ... 36
Figure 1.5 Crown surface area ... : ... 37
Figure 2.1 Total foliage biomass ... · ... 62
Figure 2.2 Foliage biomass density ... 63
Figure 2.3 Relative cumulative foliage biomass ... 64
Figure 3.1 Sample tree representation with stem disks, and total versus cumulative foliage biomass ... 94
Figure 3.2 Black and white spruce foliage biomass branch basal area relationship ... 95
Figure 3.3 Tree leve! pipe model ratio ... 96
Figure 3.4 Pipe model ratio below crown base ... 97
Tableau 1.1 Characteristics and locations of
s
ample sites
...
..
.
.
...
....
...
.
....
.
...
.
....
.
.
...
..
....
.
29
Tableau 1.2 Summary statistics for plot and tree level variables ...
.
...
.
.
.
.
.
.
.
. 30
Tableau 1.3 Bran
ch and crown characteristics for sample trees
.
...
.
...
.
...
..
.
..
...
31
Tableau 1.4
Parameter estima tes for bran ch length equations ...
.
...
.
..
.
... 32
Tableau 2.1 Characteristics and locations of sample sites
...
...
..
..
.
..
.
....
.
.
.
...
.
.. 59
Tableau 2.2 Summary statistics for plot- and tree-level
v
ariables
..
..
.
...
.
.
.
.
.
...
.
.
..
.
.
60
Tableau 2.3
Parameter estimates and their associated P-values and error term for
equations 2.3
,
2.6 and 2.9 .
.
...
.
...
.
...
...
...
.
...
.
...
.
..
...
..
.
...
...
.
....
.
.
. 61
Tableau 3.1 Characteristics and locations of
the sample sites
.
...
.
...
.
...
.
...
.
..
.
.
91
Tableau 3.2 Summary statistics for plot, tree and branch level variables .
.
..
.
....
.
...
.
... 92
Tableau 3
.
3 Parameter
estimates for branch foliage
biomass
(Eq
.
3
.
1 and 3
.
3)
,
tree-level pipe model ratio (Eq
.
3.4 and 3
.
5)
,
and disk-le
v
el pipe
model ratio (Eq. 3
.
6 to 3.8 including 3.8a)
...
.
...
.
....
.
...
.
....
.
...
.
... 93
Bd Bl Cçi Cl Cr Crwf Cs D/H DBH Df DH Dm Dmp Dmq Fd G Gb Ht Hl Hrel PDBH PHt PSP
LISTE DES ABRÉVIATIONS, SIGLES ET ACRONYMES
Branch basal diameter (mm) Branch length (rn)
Relative crown depth Crown length (rn) Crown radius (rn)
Cumulative relative foliage biomass Crown surface area (m2
)
Diameter at breast height over total height ratio Diameter at Breast Height (cm)
Degree of freedom Dominant height (rn)
Dumrny variable for species (0 black spruce; 1 white spruce) Dumrny variable for p parame ter of beta function
Dumrny variable for q parameter ofbeta function Foliage density (g/m2
) Stand basal area (m2/ha) branch basal area (mm2
) Total tree height (rn)
Horizontal branch length (rn) Relative height on tree stem
Plot mean diameter at breast height (cm) Plot mean tree height (rn)
QDBH Rd RSSE Rt Sd SI SPH Wf Wfb
Quadratic mean diameter at breast height (cm) Pipe model ratio at disk level (g/cm2
)
Residual sum of square
Pipe model ratio at tree level (g/cm2 )
Relative height between the base of the tree and crown base Site index, domiant height at 50 years old (rn)
Stand density (stems/ha) Foliage biomass (Kg) Branch foliage biomass (g)
RÉSUMÉ
L'étude des relations allométriques est depuis longtemps un élément impotiant de la biologie, et ce, pour différentes raisons allant de la prédiction de la taille et du poids d'un individu à l'explication des processus physiologiques qui gouvement sa croissance. Cette étude compare les relations allométriques qui unissent les différentes parties de la biomasse aérienne de deux espèces phylogénétiquement proches, l'épinette noire et l'épinette blanche. L'étude des relations allométriques des deux espèces d'épinettes a permis de mieux comprendre comment les caractéristiques écologiques du milieu dans lequel croissent les individus affectent leur développement morphologique et comment les traits fonctionnels propres à chaque espèce se reflètent dans leurs relations allométriques. Les résultats permettront de mieux caractériser la distribution de la biomasse aérienne de 1' épinette blanche et de l'épinette noire et éventuellement d'utiliser ces relations dans des modèles de croissance à bases fonctionnelles. Ceci permettra de mieux comprendre la croissance et le développement de ces deux espèces écologiquement et économiquement importantes.
La thèse porte spécifiquement sur les relations allométriques entre les différentes parties de la cime vivante de l'épinette noire et de l'épinette blanche et sur la biomasse foliaire et sa distribution dans la cime vivante. La thèse porte également sur la relation entre la biomasse foliaire et la superficie d'aubier de l'arbre. L'hypothèse principale soutenant ce travail de recherche est que des différences entre les traits fonctionnels des espèces permettront d'expliquer les différences de relations allométriques et l'influence des conditions de croissance sur ces relations.
Afin de répondre à ces questions, un échantillonnage destructif d'épinettes noires et d'épinettes blanches a été effectué sur quatre sites en Alberta, en Ontario et au Québec. La mesur.e du diamètre des branches vivantes de même que leur positionnement à l'intérieur de la cime vivante a permis de reconstruire la cime vivante des arbres et d'en estimer la longueur et le profile. Un échantillonnage de feuillage a également été réalisé afin d'estimer la biomasse foliaire de l'arbre et sa distribution verticale dans la cime vivante. Finalement, des sections de tiges ont été échantillonnées le long du tronc afin d'estimer la superficie d'aubier à plusieurs endroits de l'arbre. Des régressions linéaires et non linéaires mixtes ont été utilisées afin de paramétrer les relations allométriques.
L'étude a permis de constater que la longueur de la cime vivante de l'épinette noire et de l'épinette blanche ne différait pas lorsque les dimensions des arbres et les conditions de croissance étaient prises en compte. Par contre, le profil des deux espèces s'est avéré différent. L'épinette blanche montre une cime plus large dont le profil est plus sensible à la compétition exercée par les autres arbres du peuplement. De plus, l'indice de qualité de station a un effet différent chez les deux espèces laissant supposer une différence en termes d'allocation des ressources. Des différences ont également été remarquées quant à la
biomasse foliaire des deux espèces. En présence de conditions de croissance similaires, l'épinette noire supporte une plus grande quantité de feuillage que l'épinette blanche. De plus, l'épinette noire montre une plus grande densité de feuillage. Cette différence pourrait être reliée à la plus grande tolérance à l'ombre de l'épinette noire. Pour les deux espèces, la distribution verticale du feuillage a varié de façon similaire en fonction de l'âge et de la vitesse de croissance en hauteur. La relation allométrique entre la biomasse foliaire et la superficie d'aubier s'est avérée différente entre les deux espèces. L'épinette noire maintient une plus grande quantité de feuillage par surface d'aubier que l'épinette blanche. Cette différence pourrait être reliée à un besoin en eau diminué pour le feuillage de l'épinette noire compte tenu de sa plus faible productivité.
La thèse permet de révéler des différences entre les relations allométriques de 1' épinette noire et de l'épinette blanche et des différences concernant l'influence des conditions de croissance sur celles-ci. Ces différences laissent supposer que les stratégies d'investissement des ressources et que la relation face à la compétition diffère entre les deux espèces.
Mots Clefs : Cime vivante, Biomasse foliaire, Modèle tubulaire Keywords: Live crown, Foliage biomass, Pipe model
INTRODUCTION GÉNÉRALE
0.1
Relations allométriques et traits fonctionnels
Depuis longtemps, les relations allométriques ont fait l'objet de travaux, et ce, dans plusieurs domaines de la biologie (Beuchat et al., 1997 ; Niklas, 1994 ; Schmidt-Nielsen, 1984 ; West, Brown et Enquist, 1997). L'allométrie peut être définie comme l'étude des conélations entre différentes mesures reliées à la taille, à la forme ou au métabolisme des êtres vivants (Niklas, 1994). Les relations allométriques peuvent être abordées empiriquement ou physiologiquement. L'étude empirique s'intéresse exclusivement à la relation mathématique entre deux variables alors que l'étude physiologique se préoccupe des origines de la relation (Niklas, 1994). Les relations allométriques peuvent donc être utiles pour explorer les liens entre les différentes parties ou caractéristiques d'un organisme. Elles peuvent également être utiles pour comprendre comment le changement de dimension d'une partie d'un organisme induit le changement d'une autre partie et comprendre les processus physiologiques qui gouvernent ces relations (Niklas, 1994 ; Schrnidt-Nielsen, 1984). Les traits fonctionnels d'une plante, qui lui permettent de tirer avantage du milieu et de la position qu'elle occupe dans ce milieu (Wright et al., 2004), se reflètent dans les relations allométriques qui unissent ses différentes parties. On pourrait donc penser que les relations allométriques peuvent être différentes selon les milieux de croissance et selon les espèces, permettant à la plante d'utiliser les ressources de manière optimale et aux espèces de se distinguer quant à leur comportement dans un milieu donné.
L'épinette noire et l'épinette blanche sont deux des espèces arborescentes les plus répandues de la forêt boréale nord-américaine (Cauboue et Malenfant, 1988). Leurs
caractéristiques morphologiques sont généralement considérées comme étant différentes (Marie-Victorin, 1995). Bien que très peu d'études, à notre connaissance, comparent directement l'allométrie des deux espèces, il semble probable que les différences morphologiques se traduisent par des différences de relationsallométriques. Étant donné que les exigences du milieu de croissance influencent les traits physiologiques et par le fait même l'allométrie des arbres (Beminger et Nikinmaa, 1997 ; Vanninen et Makela, 2005), la variété des sites colonisés par ces deux espèces pourrait expliquer des différences de relations allométriques entre elles.
L'influence du milieu sur les relations allométriques s'observe entre autres pour le modèle tubulaire (Pipe mode! theory; Shinozaki et al., 1964b ). Cette relation qui unit la biomasse foliaire à la superficie d'aubier change avec le stress hydrique imposé à la plante. Les arbres poussant sur des sites sujets au stress hydrique maintiennent une plus petite quantité de feuillage par unité de surface d'aubier, prévenant ainsi la rupture de la colonne d'eau (Mencuccini et Bonosi, 2001). Cependant, l'influence de l'environnement ne saurait expliquer à elle seule les différences de relations allométriques et des traits fonctionnels spécifiques à l'espèce demeurent. En reprenant l'exemple du modèle tubulaire, on constate que les arbres d'essences intolérantes à l'ombre montrent des rapports entre la masse de feuillage et la surface d'aubier plus petits que les arbres d'essences tolérantes, permettant ainsi d'acheminer une plus grande quantité de sève par unité de feuillage (Waring, Schroeder et Oren, 1982). Il est ainsi plausible qu'une portion des différences entre les relations allométriques de l'épinette noire et de l'épinette blanche provienne directement de différences associées à leurs traits fonctionnels et au rapport que chaque espèce entretient avec son environnement.
Plusieurs équations allométriques ont mis en relation différentes parties de l'arbre. Parmi les relations ayant des bases physiologiques, Shinozaki et al. (1964a), avec la théorie du modèle tubulaire, mettent en relation la superficie du tronc à la base de la cime vivante et la biomasse foliaire de l'arbre, établissant ainsi un lien de conduction hydraulique entre ces deux parties de l'arbre. Enquist et al. (1999) établissent quant à eux une relation liant la masse des arbres à la photosynthèse brute, Famsworth et Van Gardingen (1995) étudient la
3
relation entre la longueur et le diamètre des branches en apportant des explications de nature hydraulique et mécanique à la relation. Le bois de la branche a deux principales utilités, soit servir de milieu de conduction de la sève et de support mécanique pennettant au feuillage de se déployer dans un environnement lumineux adéquat. La relation allométrique entre le diamètre de l'arbre mesuré à 1,3 rn (DHP) et la hauteur totale de l'arbre a également été analysée sous l'angle des exigences hydrauliques et des exigences de support mécanique (e.g. Alves et Santos, 2002; King, 2005; McMahon et Kronauer, 1976; O'Brien et al., 1995). En plus de subir des tensions hydraulique plus forte avec l'augmentation de la hauteur (Tyree et Zimmermann, 2002) l'arbre doit également accroitre son diamètre afin d'assurer sa stabilité mécanique (McMahon, 1973).
Les espèces montrant des différences au point de vue de leur autoécologie (e.g. espèces sciaphiles versus héliophile) alloueront différemment leurs ressources afin de satisfaire les exigences de leur milieu de croissance (Cannell, 1985 ; King, 2005 ; Wiemann et Williamson, 1989). Le schéma de distribution de la biomasse variera donc entre les espèces et entre les milieux de croissance. Il est alors essentiel d'analyser l'ensemble de l'arbre et tenir compte des conditions de croissance lorsque l'on compare la productivité de deux espèces différentes.
0.2 Autoécologie de l'épinette noire et de l'épinette blanche
L'épinette noire et l'épinette blanche sont deux espèces arborescentes possédant une distribution générale dans la portion nord-américaine de la forêt boréale, un biome qui occupe près de 310 millions d'hectares et qui représente 10% de la superficie forestière mondiale (Canadian Forest Service, 2005). La distribution presque identique des deux espèces couvre le continent d'est en ouest et s'étend du nord des États-Unis d'Amérique jusqu'à 69° de latitude nord au Canada et en Alaska. Par contre, la distribution commerciale des deux espèces est considérablement plus réduite (Burns et Honkala, 1990 ; Vincent, 1965). L'importance économique des deux essences est manifeste, particulièrement celle de
l'épinette noire qui est l'essence la plus employée en Amérique du nord dans la fabrication des pâtes et papiers (Burns et Honkala, 1990), qui est également employée dans le bois de construction et qui est une espèce de choix pour les bois structuraux en dans l'est du Canada (Butos et a/2008).
L'épinette noire et l'épinette blanche forment toutes deux des peuplements purs et mélangés. L'épinette noire se retrouve très souvent en peuplements purs dans le domaine bioclimatique de la pessière à mousses ou sur les sols organiques dans le reste de sa distribution (Cauboue et Malenfant, 1988). L'épinette noire s'associe également à l'épinette blanche, au pin gris (Pinus banksiana Lamb.), au sapin baumier (Abies balsamea (L.) Mill.), au mélèze laricin (Larix laricina (Du Roi) K. Koch), au peuplier faux-tremble (Populus tremuloides Michx.) et au bouleau à papier (Betula papyrifera Marshall) pour former des peuplements mélangés (Burns et Honkala, 1990 ; Cauboue et Malenfant, 1988 ; Vincent, 1965). L'épinette blanche quant à elle forme plus rarement des peuplements purs et est généralement associée à l'épinette noire, au sapin baumier, au peuplier faux-tremble, au bouleau à papier, au bouleau jaune (Betula alleghaniensis Britton) et au pin tordu (Pinus conforta Douglas ex Loudon) au sein de peuplements mélangés (Burns et Honkala, 1990 ; Cauboue et Malenfant, 1988; Sutton, 1969).
L'épinette noire et l'épinette blanche peuvent toutes deux se retrouver sur plusieurs types de stations, allant des sols minces sur les sommets de collines aux sols organiques localisés dans les dépressions (Cauboue et Malenfant, 1988 ; Sutton, 1969 ; Vincent, 1965). Malgré leur présence sur une variété de stations, les deux espèces se développent à leur plein potentiel sur des sols profonds, de texture moyenne et bien drainés (Burns et Honkala, 1990 ; Cauboue et Malenfant, 1988 ; Vincent, 1965). Cependant, l'épinette noire, grâce à son système racinaire superficiel, réussit à 1nieux croître sur les stations où la nappe phréatique se situe à proximité de la surface du sol (Burns et Honkala, 1990 ; Larsen, 1980 ; Vincent, 1965).
5
L'épinette noire atteint des hauteurs de 12 à 25 rn alors que l'épinette blanche peut atteindreplus de 30 rn (Burns et Honkala, 1990). La forme de la cime de l'épinette noire estgénéralement considérée étroite, avec les branches tombantes. On note également la présence d'un houppier au sommet de l'arbre sur les sites de mauvaise qualité (Marie-Victorin, 1995 ;
Vincent, 1965). À la différence de l'épinette noire, la forme de la cime de l'épinette blanche est généralement obtuse et arrondie (Sutton, 1969). La forme de la cime vivante des deux
espèces varie en fonction de la qualité du site sur lequel elles poussent. Le volume de bois à maturité atteint de 100 à 300m3
/ha pour l'épinette noire alors qu'il peut être beaucoup plus grand et atteindre 400m3/ha pour l'épinette blanche (Burns et Honkala, 1990). Sur des sites
de même fertilité, l'épinette blanche produit plus de bois que l'épinette noire (Pothier et Savard, 1998; Thiffault et al., 2003) bien que le bois de l'épinette noire soit plus dense et ait une meilleure résistance mécanique (Jessome, 1977). Ces différences de caractéristique
pourraient donc représenter des stratégies d'investissement différentes entre les deux espèces, l'épinette noire favorisant un tronc moins volumineux mais plus résistant à la rupture assurant
ainsi sa fonction de soutient mécanique.
L'épinette noire et l'épinette blanche sont considérées comme des espèces qm sont modérément à tolérantes à l'ombre (Burns et Honkala, 1990). L'épinette blanche atteint son
plein potentiel de croissance en hauteur sous 50% de pleine lumière mais cette tolérance diminue avec l'âge et avec la taille (Cauboue et Malenfant, 1988 ; Kneeshaw et al., 2006). L'épinette noire est considérée comme étant plus tolérante à l'ombre que l'épinette blanche et
son feuillage atteint un point de saturation lumineuse en situations plus ombragées que
l'épinette blanche (Grossnickle, 2000; Lamhamedi et Bernier, 1994; Man et Lieffers, 1997). De plus, l'épinette noire produit une réaction positive de plus grande envergure que l'épinette blanche à l'augmentation de C02 en conditions ombragées (Marfo et Dang, 2009),
confirmant sa plus grande tolérance à l'ombre.
Le feuillage de l'épinette noire et de l'épinette blanche se ressemblent à plusieurs points de vue. Les deux espèces possèdent des aiguilles quadrangulaires persistantes. Cependant, les feuilles de l'épinette blanches (1.5 à 3 cm) sont plus grandes que celles de l'épinette noire
(0.5 à 1.5 cm). (Marie-Victorin, 1995) mais elles ont un ratio surface poids plus petit que celles de l'épinette noire (Dang et Cheng, 2004). Ces résultats sont conséquents avec les résultats de Messier et al. (1999) selon lesquels le ratio surface/poids de feuillage est plus grand pour les espèces tolérantes à l'ombre que pour les espèces intolérantes.
0.3 Implications pratiques de l'étude
Les implications pratiques de l'étude des relations allométriques sont nombreuses. En foresterie, la relation entre le diamètre et la hauteur de 1 'arbre est utilisée afin d'estimer le volume des arbres à l'aide de l'unique mesure du diamètre (e.g. Perron, 1985). De nos jours, avec l'utilisation du LIDAR aéroporté, la relation inverse (prédire le diamètre à l'aide de la hauteur totale de l'arbre) peut également être utilisée (Vepakomma et al., en préparation). De nombreuses relations allométriques sont également utilisées par les modèles de croissance intégrant des éléments physiologiques (Makela, 1997). L'utilisation de ces modèles devient de plus en plus pertinent dans le contexte des changements climatiques et de l'estimation des effets de nouveaux traitements sylvicoles (Weiskittel et al., 2011). Les relations allométriques présentées dans la thèse pourront être utilisées afin de mieux comprendre comment chacune des deux espèces investit ses ressources et quel impact cette stratégie d'investissement a sur la productivité de l'arbre.
0.4 Objectif général de la thèse
L'objectif principal de la thèse est de modéliser et comparer les dimensions et la biomasse de différentes parties de 1' épinette noire et de 1' épinette blanche afin de mieux comprendre comment des espèces phylogénétiquement proches se distinguent quant· à leurs relations allométriques lorsque les conditions de croissance sont prises en considération. Les différences entre les deux espèces peuvent indiquer des différences de traits fonctionnels influençant leur autoécologie. Les relations allométriques pourront par la suite être utilisées
7 par des modèles de crOissance afin de mieux prédire la croissance de l'arbre et de ses différentes parties.
0.5 Objectifs spécifiques de la thèse
Spécifiquement, parce que les caractétistiques morphologiques de la cime vivante sont souvent mises en relation avec la biomasse foliaire et avec la croissance de l'arbre, la thèse vise à modéliser et à comparer la longueur, le profil et la superficie latérale de la cime vivante de l'épinette noire et de l'épinette blanche.
Parce que le feuillage est le moteur de croissance de 1' arbre, la thèse a également pour objectif de modéliser et comparer la biomasse foliaire à l'échelle de l'arbre, sa distribution verticale et son lien avec la surface de la cime vivante pour l'épinette noire et l'épinette blanche.
En dernier lieu, parce que cette relation est importante afin de caractériser la distribution de la biomasse de l'arbre, la thèse vise à établir et comparer la relation entre la surface transversale d'aubier et la biomasse foliaire à l'échelle de l'arbre et à plusieurs endroits dans la tige de l'arbre pour l'épinette noire et l'épinette blanche. Pour l'ensemble des objectifs précédemment mentionnés, la thèse vise également à caractériser l'effet des conditions de croissance sur ces relations. L'analyse de l'effet des conditions de croissance permettra de mieux comprendre comment les traits fonctionnels propres à une espèce lui permettent de croître de façon optimale dans un milieu donné.
DIFFERENCES IN CROWN CHARACTERISTICS BETWEEN
BLACK (PICEA MARIANA) AND WHITE SPRUCE (PICEA GLAUCA)
Hugues Power, Valerie LeMay, Frank Beminger, Derek F. Sattler et Daniel Kneeshaw
9
1.1
Résumé
L'épinette noire (Picea mariana (Mill.) B.S·.P.) et l'épinette blanche (Picea glauca (Moench)) sont deux espèces phylogénétiquement proches l'une de l'autre mais dont la productivité diffère. Malgré l'importance de ces deux espèces au sein de la forêt boréale canadienne et malgré l'importance de la cime des arbres afin de mieux comprendre et modéliser leur croissance, les connaissances sur les caractéristiques des cimes vivantes des épinettes noires et des épinettes blanches demeurent partielles. Dans cet article, nous avons caractérisé et comparé la longueur, le profil, la forme et la surface latérale de la cime vivante des deux espèces d'épinettes. Ce travail a été effectué grâce à l'échantillonnage destructif de 57 épinettes noires et 65 épinettes blanches provenant des provinces canadiennes de 1' Alberta de l'Ontario et du Québec. La longueur de cime vivante a été mesurée sur chaque arbre échantillon et le profile de cime a été obtenu à la suite de la reconstruction de la cime vivante à l'aide de mesures de longueur de branches. Notre étude montre que la longueur de cime ne diffère pas entre les deux espèces lorsque les dimensions des arbres ainsi que les conditions dans lesquelles ils croissent sont prises en compte. Cependant, 1' épinette noire et 1' épinette blanche croissent généralement dans des milieux différents entrainant des différences en termes de longueur de cime vivante. Nous avons cependant trouvé des différences entre les espèces quant au profil de la cime vivante. L'épinette blanche a montré une cime plus large qui passe d'une forme parabolique à une forme conique en présence d'une forte compétition. Ces différences se reflètent dans la surface latérale de la cime vivante des deux espèces.
Black (Picea mariana (Mill.) B.S.P.) and white spruce (Picea glauca (Moench) Voss) are phylogenetically proximal species that differ in productivity. Crown characteristics of these two species have not been extensively studied, in spi te of the importance of these two species to the Canaôian boreal forest and the importance of tree crowns for understanding and modelling tree growth. In this paper, we characterize and compare crown lengths, crown profiles (i.e, radii), shapes and surface areas of these two species using 65 white spruce and 57 black spruce trees destructively sampled in the provinces of Alberta, Ontario and Québec, Canada. Crown length was measured on every sample tree while crown profile was obtained by reconstructing crowns from branch measurements. Our results showed that crown lengths did not differ between these two species given the same tree size and growth conditions. However, these two species establish under different growth conditions resulting in crown length differences. Further, differences in crown radii and profiles were found even under the same growth conditions. White spruce had wider crown radii and profiles changed from a parabola to a cone shape under increased density. As a result, differences in crown surface areas were found.
11
1.3
Introduction
Black (Picea mariana (Mill.) B.S.P.) and white spruce (Picea glauca (Moench) Voss) are two important coniferous species of North America and they share a sirnilar geographical distribution. The shade tolerance of black spruce is considered to be intermediate to high, whereas white spruce is generally considered to be slightly Jess shade tolerant (Burns and Honkala 1990). The maximum crown width of black spruce has been reported as being narrower than that of white spruce (Marie-Victorin 1995). Black spruce generally grows to heights between 12 and 25 rn, while white spruce can reach heights of 30 rn or more (Burns and Honkala 1990). The volume at maturity for primarily black spruce stands varies from 100 to 300 m3 ha-1, while predominant! y white spruce stands can approach 400 m3 ha-1• Differences in maximum volume between these two species on sirnilar sites may be explained by a number of factors including differences in the crown characteristics and architecture.
Crown characteristics refer to changes m morphology, in particular, crown length, maximum width, shape or profile, surface area, volume, and· biomass, whereas crown architecture refers to the arrangement and distribution of branches and foliage within tree crowns. Crown characteristics and architecture affect photosynthetic rates of tree via light interception and, therefore, affect forest productivity (Cannell et al. 1987). Oker-Blom and Kellomaki (1983) studied effects of crown shape on light interception in a simulated stand of Scots pine (Pinus sylvestris L.) and found that interception varied not only with stand characteristics but also with the foliage distribution within trees. Wang et al. (1990) studied the light interception of Sitka spruce (Picea sitchensis (Bong.) Carr.) using model of photosynthesis and found that crown shape was less important for affecting light interception than the individual effects of total leaf area, foliage distribution within the crown, and leaf angles. The lesser impact of crown shape on light interception was also reported by Duursma and Makela (2007); their mode! for Scots pine light interception was influenced more by
crown smface area than by crown shape. However, since crown shape and crown smface area are strongly conelated, likely both characteristics affect light interception.
Von Gadow and Hui (1999) stated that tree crown characteristics both detennine and respond to shading and physical constraints between neighboring trees. As a result, crown characteristics are commonly used in growth models at the tree level (e.g., TASS, Mitchell 1975; CROBAS, Makela 1997) and are important variables in process-based or hybrid forest
growth models (Valentine and Makela 2005). Consequently, many authors use maximum
crown radius or crown shape to deve1op competition indices ( e.g. Hegyi 1974; Daniels et al. 1986; Biging and Dobbertin 1992; Nepal et al. 1996). In sorne models, leaf area index (LAI) defined as the ratio between total leaf area and the ground smface area, has been used as an indicator of photosynthetic productivity (Landsberg and Waring 1997). However, LAI and crown characteristics are strongly related.
Overall, crown architecture and characteristics are strongly related to photosynthetic
capability and, therefore, to tree productivity. These characteristics have been shown to vary
among species and growth conditions. Characterizing and comparing the differences in
crowns of two of the most abundant tree species of the Canadian Boreal Forest is essential for improving understanding of growth and productivity in this large forest area.
In this study, differences between black and white spruce crown characteristics were
exarnined. As noted, crown characteristics have not been extensively studied for these two
species, in spite oftheir importance. The specifie crown morphological characteristics studied in this paper were crown length, profile, and surface area. Since growth conditions are known to affect tree crowns, we examined species differences in the context of stand density and site productivity differences. A second objective was, therefore, to examine how changes in growth conditions affect crowns of these two important spruce species.
13
1.4
Materials and methods
1.4.1 Data
The study dataset included 65 white spruce and 57 black spruce trees. The white spruce data were obtained from sites located in Ontario and Alberta, Canada, whereas data for the
black spruce trees were collected in Québec, Canada (Table 1.1). Sampling at the various sites took place during the growing season (May through September) in 2008 through 2010.
The sampling sites in Alberta were spruce-dominated stands that contained a minor
component of trembling aspen (Populus tremuloides Michx.) within the Central Mixedwood Natural Sub-region (Beckingham and Archibald 1996) and had existing 0.1 ha permanent sample plots (PSPs). These stands developed from natural regeneration and were not
influenced by silvicultural treatments. The three PSPs represented a range of stand densities; however, all were located on sites with mesic soil moisture and medium soil fertility. At each
location and adjacent to the existing PSP, a minimum of four dominant or co dominant trees
with undamaged crowns were selected as sample trees. Using the selected tree as the plot centre, local competition from neighbouring trees was measured by establishing a 5.64 rn radius circular plot. Within the plot, the diameter outside bark at 1.3m above ground (DBH)
and total tree height (Ht) were measured and species was recorded for alllive trees 2 7.5cm
DBH. Unlike sampling in the other two provinces, smaller trees from 1.1 to 7.5 cm DBH were not measured. However, plots selected from Alberta sites were all in mature stands (85
to 140 years old), and there were few trees of this size. The mean of sample tree ages was calculated at each location (hereafter termed stand age). The data from the fixed-radius plots, including the subject tree, were used to calculate plot-level variables presented in Table 1.2.
White spruce trees from Ontario were sampled from existing plantations that received spacing or thinning treatments; however, the last thinning was done in 1982 and growth was assumed to be similar to that of natural white spruce-dominated stands. Sample trees from the
Petawawa site were taken inside or from the periphery of existing PSPs ranging in size from 0.03 to 0.08 ha. For alllive trees ~ 1.1 cm DBH in the PSP, species was recorded and the
DBH was re-measured; heights of the previously measured trees were re-measmed. To
obtain a distribution of samples across tree size, trees were purposively sampled by selecting one tree from each of small and large DBH classes and two trees from the medium DBH class, resulting in fom trees per plot. Each DBH class included the same number of trees. Sample trees were limited to those with undamaged crowns. Stand age was the plantation age adjusted to age at 1 rn above ground, using estimates provided by Pothier and Savard (1998). The measmes for all live trees in each PSP were used to calculate the plot-leve! variables presented in Table 1.2.
Black spruce trees from Québec were selected from even-aged stands that natmally regenerated following clearcutting or a stand-replacing fire. Stands of different ages, densities (i.e. trees per ha), and site indices were selected. For each selected stand, a variable-radius plot with basal area factor of 1 m2 ha-1 was established at a location previously deterrnined on the forest map. Species was recorded and DBH was measmed for alllive trees ~ 1.1 cm DBH. As with white spruce from Ontario, trees in the plot were divided into small, medium and large DBH classes each class including the same number of trees. Height was measmed for 20 to 90 trees across the DBH classes. For each plot, one sample tree was selected from each DBH class resulting in three trees per plot. Age at 1 rn above ground for three dominant and undamaged trees of the target species was recorded. The mean age of the three dominant trees was used as stand age. The information from the variable-radius plot was used to calculate the plot-level smnmary statistics (Table 1.2).
For all sites, selected sample trees were hand-felled. Information about the selected sample trees is presented in Table 1.3. Height and height to live crown were measured on trees after felling. Height to live crown was considered as the height of the lower branch that presented green foliage and above which all the whorls included at least one living branch. The vertical position of each live branch along the tree bole (i.e., main stem) was recorded. Branch basal diameter, branch insertion angle, and whether the branch was a nodal or an inter-nodal branch were recorded for each living branch.
15
For the trees sampled in Ontario and Québec, the crown was divided into 10 sections of equallength and one living branch from each section was randomly selected with the use of a random table and measured for branch length. For the trees from Alberta, the crown was divided in two sections of equal length and branch length was measured on 10 randomly selected branches from each section. A random selection of branches was made from a random sampling performed after the measurement of allliving branches.
1.4.2 Calculations of plot- and tree-level variables including imputations of missing heights
Sample data were compiled to obtain the plot- and tree-level variables used in modeling and examining crowns (Table 1.2). All plot-level variables were expanded to per ha measures. No attempt was made to correct for the difference in minimum DBH between Alberta and other sites in calculating plot-level variables, since all plots from Alberta were in mature stands with few stems in the 1.1 to 7.5 cm DBH range. Also, for all models, random-effects at the site-, plot-, tree, and branch-level (where appropriate) were included, thereby accounting for correlation and heteroscedasticity due to the four-leve! (or three-level) sampling hierarchy.
Unmeasured heights of trees in each plot from Québec and Ontario were imputed using the following models for black spruce (Eq. 1.1) and white spruce (Eq. 1.2):
(1.1)
(1.2)
HtiJk =-8.5523+0.6216DBHiJk -0.0184DBH~k +1.6978PDBHiJ -0.0619PDBH~ +0.0005PHTiJ +0.0298DBHiJkPDBHiJ +&; +&j(il +&kWJ
HtiJk
=
8.5477 +0.2638DBHiJk -0.0079DBH~k +0.2660PDBHiJ-0.0163PDBH~ +0.0659PHTü +0.0182DBHiJkPDBHiJ +&; +&jUJ +&kWJ
Where: PDBH is the plot mean DBH, all live trees; PHt is the plot meàn height,
height-measured n·ees only; ijk are the subscripts for tree k in plot j in site i and Ei, Ej(i}. and Ek(i})
are the random error terrns at the site, plot within site and tree within plot levels, respectively.
Eq. 1.1 and 1.2 were fitted using the height and DBH measured on the adjacent sample
plot and the lme function of R (Pinheiro et al. 2010). Subject-specific (i.e., site and plot
nested in site levels) random effects on the intercept were added to each predicted total
height. Tree height predictions were used in imputing any rnissing heights in each plot. The error terrns at the site and plot levels were assumed to follow normal distributions with the
hierarchical structure of plots within sites accounting for any correlation between plots. The
final error terrn at the tree level was also assumed to follow a normal distribution with the
hierarchical structure accounting for correlations of trees within plots. Tree-level error terrns
were also checked for homogeneity of variance using residual plots and for norrnality using
normality plots. Where heteroscedasticity of the tree-level errors was detected, a function to
model the variance of the tree-level residuals was added to the model (Pinheiro et al. 2000).
Further, the chosen models were linear in the parameters, but nonlinear in the variables (i.e.,
a linear model) to address the expected curvilinear relationship of height with DBH. Therefore, since these height predictions equations were only used to impute unmeasured
heights and should not be used with other datasets, these linear were not further discussed.
The dominant height (DH) for each plot was calculated by averaging the height of the
100 largest trees (by DBH) per hectare. Quadratic mean DBH (QDBH, cm), density (stems
- - - -- - -- - - - -- -- - - --- - - - -
-17 comprised the main cohort of the ·stand. We excluded saplings as part of the regeneration cohort for all stands except for three young stands of black spruce (i.e., 20 to 30 years old) where saplings were considered as part of the main cohort. The site index (SI; site height at 50 years total age) for each plot was calculated using the HD, stand age and SI equations by Pothier and Savard (1998).
1.4.3 Crown reconstruction
In order to analyze the crown characteristics, we reconstructed the crowns of each sample tree. Crown reconstruction involved two steps: first, the lengths of allliving branches were imputed by fitting models using measured branch lengths (Bl); and second, horizontal distances between the bole of the tree and the tip of each branch (Hl) were calculated using the branch lengths and branch angles (Fig. 1.1 ).
To estimate the lengths of the living branches, nonlinear rnixed~effects models were fitted using sampled branches separately for black and white spruce using the nlme function in R (Pinheiro et al. 2000). To fit these models, first models of branch length using branch diameter (Bd) and relative crown depth (Cd) as predictor variables and assuming a single error terrn were developed, where relative crown depth was the relative position in the living crown equal to 0 at the top of the tree and equal to 1 at the base of the living crown. The residuals at the plot, tree and branches level from these models were graphed against the tree-and plot-level variables presented in Table 1.2 for each species and these graphs were used as a guide for selecting additional predictor variables. Each additional predictor variable was entered in the mode! and the new models were tested against the original model (i.e., Bd and Cd only) using likelihood ratio tests ( a=0.05).
Once predictor variables were selected, random effects at the site, plot, tree and branch levels were used to modify all fixed-effects parameters with a single error term at the branch level. Errors at all four levels in the hierarchy (i.e., site, plot, tree, and branch) were initially considered. However, these additional error terms are equivalent to adding an intercept to the model for subject-specific branch length estimates. Since this could result in negative
estimated branch lengths, only the error term at the branch level was retained. All random effects were considered normally distributed with equal variances; these assumptions were
verified for the branch-level enor tei:m using the same methods as for 1.1 and 1.2.
The final modelS included the predictor variables Cd, Bd and SI for black spruce (Eq.
1.3) and only Cd and Bd for white spruce (Eq. 1.4).
(1.3) Bl ijkl
=
f/1 RBdcijkl f32+b;+bj(i)+bk(ij))c,~uijPJ.kl s1iJ P. 4+
c.lC' (ijk)(1.4) BI iJkl
=
P1 aBdcf3iJkl 2+b;+bj(i)+bk(ij))C'..Jf3uiJkl 3+
&l(iJk)Where ijkl is the subscript for branch l in tree k in plot j in site i;
fJ
are fixed-effectsparameters; b are random-effects parameters at the site, plots within site, and trees within
sites levels; and E~Wk:) is the random enor at the branch level.
Eq. 1.3 and 1.4 included random effects on
P
2 at the site, plot nested in site and treenested in site and plot levels for tree-specific estima tes of the parameter
/3
2 only. These finaltree specifie branch length models were applied to each tree and living branch position to
obtain the subject-specific estimated branch length.
In the second step of the crown reconstruction, we calculated the estimated horizontal
distance (Hl) between the tree bole and the tip of the living bran ch using the estimated BI and
the measured branch angle (8) (Eq. 1.3,1.4, 1.5).
19
1.4.4 Crown lengthTo study differences in crown lengths between the two spruce species, first, a nonlinear rnixed-effects model was fitted using the nlme function of R (Pinheiro et al. 201 0) using the pooled data for both species (hereafter termed the base model). Once the base model was selected, dummy variables were used to alter the parameters of the model to be species-specific. The test for species differences was then performed by comparing the base model to the species-specific model using a likelihood ratio test (a=0.05). The modeling approach perrnitted the use of non-linear forms of equations, forms of equations frequently used in allometry, this can be compared to the analysis of co-variance where the relationship between the co-variables and the dependent variable is linear.
For the base model using pooled data for both species, different nonlinear model forms from Davies and Pommerening (2008), Antos et al (2010), Thorpes et al. (2010), and Sattler and LeMay (2011) along with different predictor variables were fitted and compared using Akaike's information criterion (AIC; Akaike 1973) as a measure of model fit. The Akaike information cri teri on allows us to compare the likelihood of two models while penalizing for the number of parameters included in the model. This method can be used to compare two models built with the same dataset and the same random structure but with differences in
their fixed effects. However, the Akaike information criterion does not provide an evaluation of the fit of the models to the data. Random effects at the site, plot and tree levels were included in all models, with the same distributional assumptions as for the height imputation models. Eq. 1.6 was selected as the base model for the pooled data of both species. This model was then modified by including a dummy variable for species (Dm is 0 for black spruce and 1 for white spruce) to alter all fixed-effects parameters (Eq. 1.7).
Where DIH is the ratio ofDBH over tree height; ijk is the subscript for tree k located in plot} located in site i;
f3
are fixed-effect parameters; and E represents the error terrns (i.e. random-effects) of the model at the site (Ei.), plot within site (Ej(i)) and tree within plot (EkUJ)) levels.A likelihood ratio test (a=0.05) was then used to detennine whether there were differences between the two species by comparing Eq. 1.6 (reduced model) and Eq. 1.7 (full model). The likelihood ratio test allows us to perform a hypothesis test on the fit of two models where one of the two models is a special case of the second one. To further examine differences between the two species, graphs of the predicted crown length using fixed effects only (i.e., population-averaged level) versus combinations ofpredictor variables were used.
1.4.5 Crown profile
For our study, crown profiles were defined by ~rown radii (Cr) calculated as the average of the four longest horizontal branch lengths within each inter-whorl segment defined by the nodal and inter-nodal branches beginning with whorls at the top of the segment. For these models, crown depth was redefined using the position of the mid-point of the whorl segment.
A nonlinear mixed-effects model fitted using the nlme function in R (Pinheiro et al. 201 0) was used to model the crown radii representing the tree crown profile. The random effects were errors at the site, plot and tree and whorllevels, with similar assumptions as for the branch length imputation models.
Initially, the same methods used to fit the crown length models (i.e., full versus reduced models) were used for the crown profile model. However, difficulties were encountered in fitting the species-specific model. As a result, separate crown profile models were fitted for
21
each species (Eq. 1.8) and then for the two species combined. Because the comparison of the three models could not be done with the likelihood ratio test or the Akaike criterion, the residual sum of squares (RSSE; alllevels of enors included) for the model using the pooled data was compared to the sum of RSSEs for separate black and white spruce models. Large differences were used to indicate that the crown profiles differ between species.Where ijkl is the subscript for whorl l in tree k in plot j in site i; /31 to f35 are fixed-effects parameters; SPH is the stems per ha; andE represents the enor terms of the model at the site
( Ej(i)), plot within site ( Ej(i)) tree within plot ( E~.: (ti)) and bran ch within tree ( E~(i.jk)) levels;
and all other variables were previously defined.
This model provided better results than a power or exponential models (i.e., lower AIC) using the pooled data for both species. The same method as with crown length was used to examine differences between species. Further, subject-specific crown radii predictions (i.e., tree-level branch predictions) were used in calculating crown surface areas.
1.4.6 Crown surface area
The crown surface area (Cs) was also compared between the two species. Crown surface area was obtained by integrating the crown circumferences from the base of live crown to the tree tip. The subject-specific estimated radii from the crown profile
equation were used in the equation provided in Husch et al. (2003) for a parabola shape(Eq.
" fHt ~~
dCr.
.kl
2 (1.9)CsiJk
=
2n h
CriJkt
1+ (
IJ )d(xiJkt)
dxijkl
Where x is the distance from the base of live crown (Cb) to a maximum at tree height (Ht);
ijkl is the subscript for whorl l in tree k, plot j and site i; and Cr is the predicted crown
radius.
The crown surface areas were compared between species using graphs of the crown surface area plotted against crown length.
1.5
Results
1.5.1 Crown reconstruction
The root mean squared error (RMSE) including all errors of the branch length model (Eq. 1.4) combined (i.e., site+plot+tree+branch errors) was 16.77 cm for black spruce (Eq. 1.3) and 22.31 cm for white spruce. As expected, branch diameter was the most important variable in estimating branch lengths (Eq. 1.3 and 1.4) with longer branches associated with larger branch diameters. For a given branéh diameter, branch lengths increased from the tree top with a crown depth of 0 to the crown base with a crown depth of 1 (Table 1.4). Site index had a slightly positive effect on black spruce branch lengths, but this effect was not detected for white spruce where site index was not statistically significant ( a=0.05).
1.5.2 Crown length models
The RMSE of all errors combined was only slightly larger for the reduced model without species (2.090 rn) (Eq. 1.6) than for the full model with species (2.058 rn) (Eq. 1.7). Also, the
likelihood ratio test (a=0.05) for the reduced model versus the full model with species
23
Eq. 1.7, p-value = 0.3098). Crown lengths did not vary between black and white spruce after
accounting for effects ofheight, D/H and QDBH on crown length (Table 1.4).
Using the fitted crown length model, a positive, monotonically increasing relationship
between crown length and tree height was observed (Fig. 1.2 A). Crown length also
increased with D/H, indicating a longer crown length for trees with greater taper. Conversely,
crown length decreases with QDBH.
Although differences between the two species were not detected given the same height,
D/H, and QDBH values, the sample data and previous literature indicate that the two species
grow in different characteristics. Using the sample data, the 1 st and 3'd quartiles of the D/H
and QDBH distributions were lower for black spruce (i.e., lower than for the pooled data)
than for white spruce (i.e., higher than for the pooled data). Using these species-specific D/H
and QDBH values, for a given height, black spruce crowns were longer than white spruce
crowns (Fig. 1.2 B and C).
1.5.3 Crown profile models
The crown profile model (Eq. 1. 8) fitted using the poo led data resulted in a larger RSSE
(i.e., all errors pooled) (313.925) than the sum of the RSSE obtained by fitting separate equations for the two species (282.7454), indicating that crown profiles differ between
species (Table 1.4). For both species, the maximum predicted crown radius was located at
the base of the crown.
Given the same crown depth and crown length, black spruce crowns were generally
narrower than white spruce crowns except for stands of high density and site index (Fig. 1.3). Site index had a small negative effect on white spruce crown radius while showing a small positive effect on black spruce crown radius. Stand density showed a stronger negative influence on crown radii for white spruce than for black spruce.
To compare the crown shape of both species, the relative crown radius was graphed against the relative crown depth (Fig. 1.4). Black and white spruce both had parabolic shapes
for stands with low stand density; however, white spruce crown shapes were more sensitive
to changes in stand density. That is, an increase in stand density induced a shift from a parabolic to a conical shape for white spruce.
1.5.4 Crown surface area
The noted differences in crown profiles between the two spruce species resulted in differences in crown surface areas (Fig. 1.5). The crown surface area increased nonlinearly
with crown length for both species. White spruce tended to have a 1arger crown surface area
than black spruce under low stand density, while black spruces had larger crown surface area
under high stand density. As for crown profile, white spruce crown surface area decreased with site index while black spruce crown surface area showed the opposite trend.
1.6
Discussion
1. 6.1 Crown length
Crown length dynarnics are a result of the interaction of two processes, crown recession and
height growth. Crown recession occurs when the branches located at the base of the living crown die. The rate of crown recession is known to be largely influenced by light availability
at the crown base (Sorrensen-Cothem et al. 1993), by stand density, and by height growth
(Valentine et al. 1994; Kantola and Makela 2004). Physical interactions between branches of neighboring trees are also known to influence the crown recession rates (Putz et al. 1984).
Trees that are growing in stands with higher stem densities or trees with larger crowns may
experience more physical interactions with crowns of their neighbouring trees resulting in
25
trees with a larger maximum crown radius and lower shade tolerance would have shorter
crown length than black spruce.
However, no significant differences were found between black and white spruce after
accounting for differences in 0/H and QOBH (i.e., using model predicted values given 0/H
and QOBH). The negative relationship between crown length and QOBH (Fig. 1.2 A)
coupled with larger values of QOBH for white spruce (Table 1.2) could explain the shorter
crowns of white spruces under species-specific growth conditions (Fig. 1.2 B and C). As
trees grow, heights increase resulting in increases in potential crown lengths. As a result,
tree height was an important variable for estimating crown length for both species.
Oavies and Pommerening (2008) reported that the addition of spatial and non-spatial
competition indices improved the prediction of crown length for Sitka spruce. In our models,
the plot-level variable QOBH was an important variable. QOBH can be mathematically
interpreted as: i) the diameter of the tree with average basal area; ii) the mean of squared
diameters, bence the term "quadratic mean OBH"; or iii) a measure of basal area per hectare
relative to stems per ha. As such, QOBH is indicative of competition at the plot level. The
inclusion of 0/H in the model can be interpreted as an indicator of tree social position and
competitive environment as noted by Clyde and Titus (1987), factors known to influence
crown length (Maguire and Hann 1990). In our sample data, dominant trees had higher 0/H
values than co dominant and intermediate trees. Once 0/H and QOBH were included in the
model, other measures of plot-level competition (i.e., stems per ha) and non-spatial tree-level
competition (i.e., basal area of larger trees) did not improve the model.
1.6.2 Crown profile
Site quality also impacts the crown profile (Gillespie et al. 1994). In our study, the opposite effect of site index on crown radius between the two species may be attributed to the
difference in the expected ranges of site index values for black and white spruce as reflected
(Burns and Honkala 1990); in this study, sampled black spruce stands had lower site indices thau sampled white spruce stands. The trend towards shorter branches for black spruce trees (with implied shorter crown radii) in stands with lower site indices may reflect a choice in resource allocations where investing less carbon in branches may allow the tree to invest more in the root system, a common trend for trees growing on poor sites (Cannell 1985).
Conversely, for white spruce that grows on better sites, an increase in site index may be associated with an increase in competition (Husch et al. 1982) and therefore less space for growing large branches.
Competition (Ducey 2009) and social class (Hann 1999) are factors that are known to affect maximum crown radius. In our model, density was included as a measure of competition in crown profile models. The stronger effect of stand density on white spruce may be attributed to both lower shade tolerance and to wider crowns, since wider crowns are more subject to physical interactions and increased crown recession rates (Putz et al. 1984).
Other variables representing the inter-tree competition were absent from the crown profiles models. However, since crown length was shown to be impacted by D/H and crown length was the most important predictor variable in the crown profile models, the impacts of competition on crown profile may be reflected in changing crown lengths. Deleuze et al. (1996) proposed that trees may slow their branch growth and consequently conserve long crowns even under high competition. However, the positive association of crown length and crown radius in our models is consistent with results obtained by Valentine et al. (1994) for Sitka spruce, Gilmore and Seymour (1997) for balsam fir (Abies balsamea (L.) Mill.) in Maine, US, and with Sattler and LeMay (2011) for white spruce in rnixed-species stands of British Columbia, Canada.
1. 6.3 Crown shape
Shape can be defined as the relationship between two different measures of an object (Niklas 1994). In our case, crown shape was defined as the re1ationship between crown radius