SYLVAIN LERAT
ÉTUDE DES RELATIONS SOURCE/PUITS DE
CARBONE DANS LA SYMBIOSE
ENDOMYCORHIZIENNE À ARBUSCULES
Thèse présentée
à la Faculté des études supérieures de l'Université Laval pour l’obtention du grade de Philosophiae Doctor (Ph. D.)
DÉPARTEMENT DE BIOLOGIE FACULTÉ DES SCIENCES ET DE GÉNIE
UNIVERSITÉ LAVAL QUÉBEC
AVRIL, 2003
Résumé long
Le carbone (C) est l’un des éléments clé sur lequel repose toute symbiose mycorhizienne. La présente thèse regroupe plusieurs aspects des relations source/puits de C mettant en jeu des champignons mycorhiziens à arbuscules (MA) associés à diverses plantes hôtes. Les composantes essentielles régissant les mouvements de C entre partenaires fongiques et végétaux ont été étudiées à différents niveaux d’intégration.
Un premier volet éco-physiologique a mis en évidence le transfert de C en conditions naturelles entre espèces végétales de phénologie différente connectées par des champignons MA. Au printemps, des érythrones photosynthétiquement actifs fournissaient du C à des juvéniles d’érable à sucre au moment où ceux-ci développaient leurs feuilles de l’année. À l’automne, un transfert de C des juvéniles d’érable vers les érythrones renouvelant leur système racinaire a également été observé.
La force de puits de C des champignons MA a ensuite été étudiée en relation avec la diversité fongique et végétale à l’aide d’un système «split-root». La force de puits représentée par Gigaspora rosea, Glomus intraradices et Glomus mosseae a été comparée chez l’érable à sucre et l’orge. La force d’attraction du C de ces trois champignons variait en fonction de la combinaison d’espèces fongique et végétale. Trois souches de G. mosseae ont également été testées sur l’orge. Ces travaux ont révélé que la force de puits de C des champignons MA varie aussi en fonction de la souche fongique.
Finalement, le statut MA d’érables à sucre matures a été étudié dans un contexte de perturbation naturelle (verglas) ayant gravement endommagé leur houppier. Les érables à sucre affichant une mauvaise reprise de croissance l’année suivant le verglas avaient des taux de colonisation mycorhizienne supérieurs à ceux des érables à bonne reprise de croissance. Par contre, les populations de spores MA récoltées autour de ces deux catégories d’arbres ne différaient pas d’un point de vue taxinomique.
ii L’ensemble de ces travaux montre que les besoins en C des champignons MA dépendent de l’identité des partenaires fongique et végétal impliqués dans la symbiose ainsi que du stade phénologique et de l’état de santé de ce dernier.
Long summary
Carbon (C) is a key element underlying the establishment of all mycorrhizal symbioses. The present thesis investigates several important aspects of C source-sink relationships between arbuscular mycorrhizal (AM) fungi and diverse host plants. The essential components ruling C movements between fungal partners and host plants have been studied at different levels of integration.
In an initial ecophysiological-based study carried under natural conditions, C transfer between plants exhibiting different phenologies but connected by a common network of AM fungal hyphae was documented. Briefly, in spring the photosynthetically active Erythronium americanum plants supplied sugar maple saplings with C during leaf expansion of the latter. By contrast, a C transfer from sugar maple saplings towards E. americanum roots was observed during the autumn.
In a second series of experiments, the effect of the fungal taxon and the host plant species on C sink strength of AM fungi was studied using a split-root system. The sink strength represented by Gigaspora rosea, Glomus intraradices and Glomus mosseae was compared using sugar maple and barley host plants. The C draining force of the three AM fungi varied with the plant host used. In a further study, three G. mosseae strains were tested with barley. This work revealed that C sink strength of AM fungi was also fungal strain dependent.
Finally, the AM status of mature sugar maple trees exhibiting either poor or good regrowth following severe crown damage during the 1998 ice storm was studied. Sugar maple trees exhibiting a poor regrowth in the year following the ice storm had higher root colonization levels than those exhibiting good regrowth. However, AM fungal spore populations collected around trees in both categories did not differ significantly.
iv In conclusion, the present study shows that the C demand of AM fungi depends on the identity of both the fungal and plant partners involved in the symbiosis, and on the phenological stage and health of the latter.
Résumé court
Les relations source/puits de carbone (C) dans la symbiose mycorhizienne à arbuscules (MA) ont été examinées sous différents aspects. Une première partie éco-physiologique rapporte l’existence d’échanges de C entre espèces végétales en conditions naturelles via des champignons MA répondant à la phénologie différente des plantes impliquées. Deux études physiologiques basées sur le système de «split-root» ont montré que la force de puits de C des champignons MA est dépendante de l’espèce fongique, de la souche fongique, ainsi que de l’hôte végétal. Enfin, une étude écologique se positionnant dans un contexte de perturbation naturelle (verglas) endommageant le houppier d’arbres matures a révélé qu’une mauvaise reprise de croissance post-traumatique peut être associée à des taux de mycorhization racinaire plus élevés que ceux d’arbres à bonne reprise de croissance. En résumé, les besoins en C des champignons MA varient en fonction des espèces impliquées dans la symbiose et du stade phénologique de chacune d’elles.
vi
Short summary
Different aspects of the carbon (C) source-sink relationships in the arbuscular mycorrhizal (AM) symbiosis were investigated. In an initial ecophysiological-based study carried under natural conditions, the existence of AM fungal mediated C exchanges between plant species with different phenology was reported. Two physiological studies based on a split-root system showed the C sink strength of AM fungi to be fungal species, fungal strain and plant species dependent. Finally, an ecological study carried out subsequently to the ice storm of 1998 (natural disturbance) which severely damaged mature tree crowns revealed that poor post-traumatic crown regrowth was associated with higher mycorrhizal colonization levels than in good regrowth trees. In conclusion, the C demand of AM fungi varies with the species involved in the symbiosis and with the phenological stage and health of the phytobiont.
Avant-Propos
Ce n’est qu’au moment de terminer cette thèse que je mesure l’ampleur du chemin parcouru depuis mon entrée à l’université il y a plus de dix ans déjà. Le travail présenté dans cet ouvrage ne compile pas seulement mes quelque trois années et demi passées à l’Université Laval mais représente l’aboutissement ultime du parcours scolaire de toute une vie. Alors bien évidemment, mes premières pensées vont à mes parents et à ma famille qui m’ont encouragé depuis toujours et continuent encore à me soutenir.
Je tiens en premier lieu à remercier ma directrice de doctorat, Line Lapointe. Il est impossible de réaliser un doctorat sans avoir une personne qui vous accorde sa confiance et qui fournit un support scientifique et financier. J’ai eu la chance que Line m’offre cette opportunité et je lui en suis très reconnaissant. Sa disponibilité, son dévouement et sa générosité m’ont été des atouts précieux.
Je remercie Yves Piché, mon co-directeur qui a su me guider dans le dédale du monde des champignons endomycorhiziens. Les discussions avec Yves furent toujours enrichissantes, bien au-delà du contexte scientifique.
Je remercie les autres membres de mon comité pour avoir généreusement accepté de corriger ma thèse: Gilles Houle, mon prélecteur (et conseiller en statistiques); John Klironomos, mon examinateur externe et J. André Fortin.
J’adresse une pensée particulière à Horst Vierheilig qui a largement contribué au contenu des travaux présentés ici. Il fut aussi et surtout un compagnon de tous les jours et un ami. Merci.
Il y a deux personnes qui s’ignorent peut-être mais qui, par leur présence et leur action lors de périodes difficiles, ont été déterminantes dans l’aboutissement de mon doctorat. Il y a tout d’abord Jean Guy Catford, qui a également été mon guide en mécanique automobile, et Karoliina Niemi.
viii
Andrew Coughlan fut une aide inestimable. Ses connaissances du travail de laboratoire et du monde des champignons furent tout autant appréciées que son éternelle gentillesse et son sourire.
Un grand merci à mes collègues de laboratoire qui ont rendu ces années agréables: Julie Samson, Marie-Claude Richer, Rachel Gauci, Amine Badri et Sylvain Gutjahr. Je suis également reconnaissant envers mes aides de terrain: Michel Bergeron, Marie-Claude Routhier et Simon Boudreault.
Finalement, j’éprouve une reconnaissance très spéciale envers ma dulcinée, Caroline Girard, sans laquelle je n’aurais pas finir de rédiger cette thèse et soutenir dans les meilleures conditions. Mille mercis à toi, Caroline.
Table des matières
Résumé long ... i
Long summary ... iii
Résumé court ...v
Short summary... vi
Avant-Propos ... vii
Table des matières ...x
Liste des tableaux... xii
Liste des figures ... xiii
Introduction...1
CHAPITRE I 14C transfer between the spring ephemeral Erythronium americanum and sugar maple saplings via arbuscular mycorrhizal fungi in natural stands ...10
1.1 Avant-Propos ...10
1.2 Résumé...10
1.3 Abstract...11
1.4 Introduction...12
1.5 Materials and methods ...15
1.5.1 Study site...15
1.5.2 Experimental design ...15
1.5.3 Labelling experiments with 14C ...16
1.5.3.1 Spring labelling...16
1.5.3.2 Autumn labelling ...17
1.5.4 Determination of radioactivity...17
1.5.4.1 Spring labelling...17
1.5.4.2 Autumn labelling ...18
1.5.5 Mycorrhizal colonisation levels...18
1.5.6 Statistical analysis...18
1.6 Results...18
1.7 Discussion...21
1.8 Conclusion ...26
CHAPITRE II Carbon partitioning in a split-root system of arbuscular mycorrhizal plants is fungal and plant species dependent...27
2.1 Avant-propos ...27
2.2 Résumé...27
2.3 Summary...28
2.4 Introduction...28
2.5 Materials and methods ...30
2.5.1 Experimental design ...30
2.5.2 Biological material and growing conditions...30
2.5.3 14CO2 labelling...32
2.5.4 Effect of AM fungal species on growth of barley ...32
2.5.6 Statistical analyses ...33
2.6 Results...34
2.6.1 Mycorrhizal colonization levels...34
2.6.2 14C partitioning ...34
2.6.2.1 Barley host plants...34
2.6.2.2 Sugar maple host plants ...38
2.6.3 Growth of mycorrhizal barley...38
2.7 Discussion...40
2.8 Conclusion ...43
CHAPITRE III Variable carbon-sink strength of different Glomus mosseae strains colonizing barley roots...44
3.1 Avant-propos ...44
3.2 Résumé...44
3.3 Summary...45
3.4 Introduction...45
3.5 Materials and methods ...47
3.5.1 Biological material and growing conditions...47
3.5.2 14CO 2 labelling...47
3.5.3 Measurement of root colonization ...48
3.5.4 Statistical analyses ...48
3.6 Results...49
3.7 Discussion...51
3.8 Conclusion ...52
CHAPITRE IV Arbuscular mycorrhizal colonization and spore populations associated with Acer saccharum exhibiting either good or poor crown regrowth following ice storm damage ...54
4.1 Avant-propos ...54
4.2 Résumé...54
4.3 Abstract...55
4.4 Introduction...55
4.5 Materials and methods ...57
4.5.1 Plots and tree selection ...57
4.5.2 Sampling ...58
4.5.3 Quantification of mycorrhizal colonization...58
4.5.4 Spore analysis ...59 4.5.5 Statistical analysis...59 4.6 Results...60 4.6.1 Mycorrhizal colonization...60 4.6.2 Spore survey ...60 4.7 Discussion...62 4.8 Conclusion ...68 Conclusion générale...69 Bibliographie ...75
Liste des tableaux
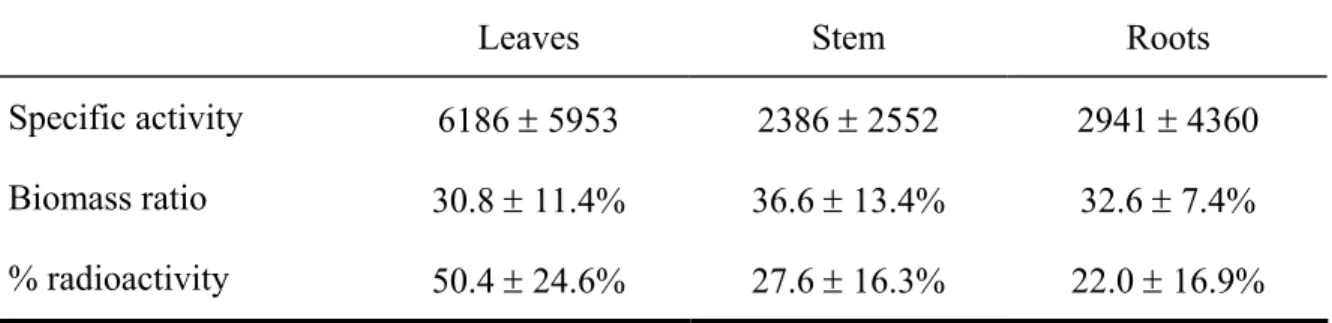
Table 1.1. Specific activity (dpm g-1 dry matter), biomass ratio, and distribution of radioactivity in leaves, stem, and roots of sugar maple saplings one wk after spring labelling. Data represent means ± 1 SD...21
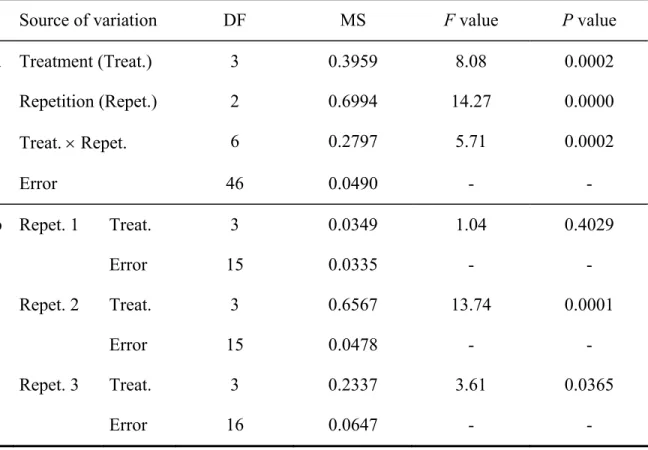
Table 2.1. Degrees of freedom (DF), mean sum of squares (MS), F values, and P values of (a) a two-way ANOVA and (b) one-way ANOVAS for each repetition
performed on percentage 14C allocated to M barley roots (data arcsin-transformed)...37
Table 2.2. Degrees of freedom (DF), mean sum of squares (MS), F values, and P values of a two-way ANOVA performed on percentage 14C allocated to M
sugar maple roots (data arcsin-transformed). ...40
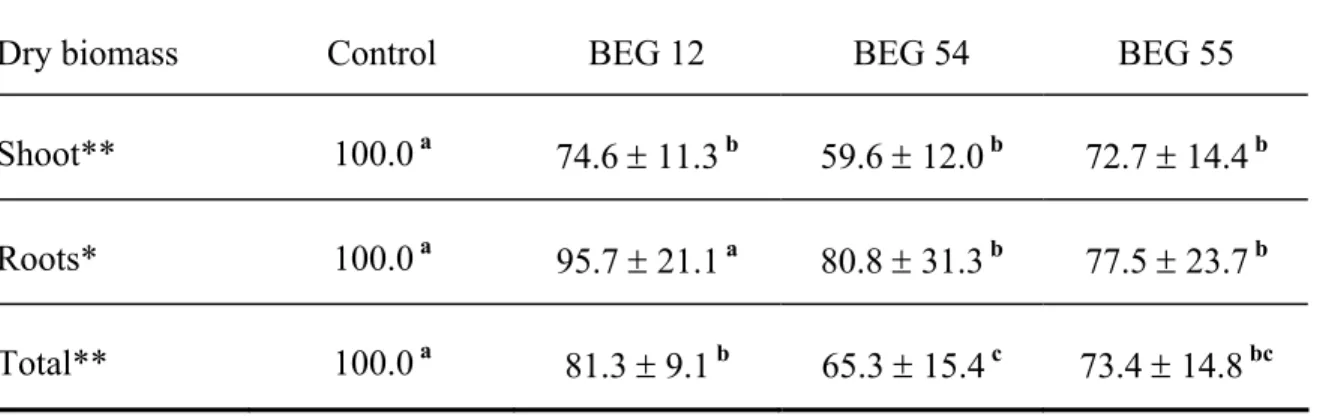
Table 3.1. Dry biomass (± 1 SD) of control and Glomus mosseae BEG 12, BEG 54 and BEG 55 treatments expressed as percentage of control. Treatments within a row with the same letter are not statistically different...51
Liste des figures
Figure 1.1. Phenology of Erythronium americanum and leaves of sugar maple saplings and mature trees (a) and relative PAR (photosynthetically active radiation) on the ground of a maple forest (b) over a growing season in the region of Québec City. Grey bars represent development (beginning of line) or senescence (end of line) of leaves. Dates of labelling with 14CO
2 are represented by arrows. ...14
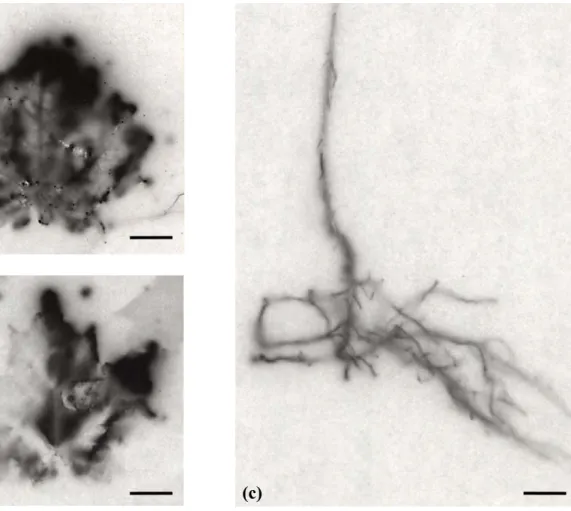
Figure 1.2. Macro-autoradiographs of leaves (a and b) and stem and root system (c) of labelled C in receiver sugar maple saplings. Dry plant material was exposed for six wk on Kodak scientific imaging films. Bars represent 1 cm. ...20
Figure 1.3. Relationship between the percentages of 14C exported to sugar maple saplings following spring labelling of Erythronium americanum and the fraction of
14C in the corm of labelled E. americanum. ...22
Figure 2.1. (a-c) Mean total 14C (± 1 SD) partitioned and (d-f) mean percentage dry weight (± 1 SD) of M (open bars) and NM (solid bars) barley roots, for each repetition, in control (compartments B and C), Gigaspora rosea, Glomus intraradices and Glomus mosseae treatments. Mycorrhizal colonization levels (± 1 SD) are shown within M bars (a-c). When the ANOVA within the repetition was significant (b,c) an
LSD test was performed (treatments with the same letter are not statistically different). Gi. rosea=Gigaspora rosea; G. intra.=Glomus intraradices; G. moss.=Glomus mosseae. ...35
Figure 2.2. (a) Mean total 14C (± 1 SD) partitioned and (b) mean percentage dry weight (± 1 SD) of M (open bars) and NM (solid bars) sugar maple roots in control (compartments B and C), Gigaspora rosea, Glomus intraradices and Glomus mosseae treatments. Mycorrhizal colonization levels (± 1 SD) are shown within M bars (a). As treatment showed significant differences (a) an LSD test was performed (treatments with the same letter are not statistically different). Abbreviations are as in Fig. 2.1. ...36
Figure 2.3. Relationship between percentage 14C allocated to M roots (arcsin-transformed) and the percentage of barley root length colonized by (a) Gigaspora rosea and (b) Glomus intraradices. Dots represent the two out of five plants per repetition used to assess M colonization levels. r2 values of the correlation are
xiv
Figure 3.1. (a) Mean total 14C (± 1 SD) partitioning and (b) mean percentage dry biomass (± 1 SD) between M (open bars) and NM (solid bars) barley roots in control and Glomus mosseae BEG 12, BEG 54 and BEG 55 treatments. Means are calculated from the sum of the two repetitions. Treatments with the same letter are not statistically different. Mycorrhizal colonization levels (± 1 SD) are shown within M bars (a). ...50
Figure 4.1. Mean percentage (± 1 SD) AM root colonization by internal hyphae, coils, arbuscules and vesicles and percentage total AM root colonization as a function of tree health (good regrowth, solid bars; poor regrowth, open bars; n=23). *, P<0.05; **, P<0.01...61
Figure 4.2. Log-transformed mean spore number per kg of dry soil (± 1 SD) for the most frequently observed AM taxa as a function of tree health (good regrowth, solid bars; poor regrowth, open bars). The 11 fungi are Acaulospora lacunosa (A. lac.), Sclerocystis rubiformis (S. rub.), Glomus aggregatum (G. agg.), G. arborense (G. arb.), G. constrictum (G. con.), G. fasciculatum (G. fas.), G. macrocarpum (G. mac.), G. warcupii (G. war.), Glomus sp1, Glomus sp2, and Glomus sp3...63
Figure 4.3. Frequency of occurrence of the most frequently observed AM taxa as a
function of tree health (good regrowth, solid bars; poor regrowth, open bars). Abbreviations are as in Fig. 4.2. ...64
Introduction
Dans la nature, la majorité des végétaux terrestres vit en symbiose avec des champignons. Cette étroite relation entre les plantes supérieures et les micro-organismes que sont les champignons s’élabore au niveau des racines. Les organes résultant de cette association sont appelés mycorhizes, du grec mukês pour champignon et rhiza pour racine. On estime à 90% la proportion de familles de plantes possédant des représentants pouvant former des associations mycorhiziennes (Harley & Smith 1983). De plus, il a été avancé par Taylor & Osborne (1995) que les végétaux supérieurs n’auraient pu conquérir la terre ferme s’ils n’avaient été associés à des champignons mycorhiziens, soulignant ainsi le rôle crucial qu’ont joué, et jouent encore, les mycorhizes dans l’évolution des plantes terrestres.
La mycorhize est une association de type symbiotique, c’est-à-dire que chacun des partenaires engagés (symbiotes) tire des profits de cette union. Le terme symbiose exclut ainsi toute notion de parasitisme ou de commensalisme où seul l’un des individus recevrait de son hôte, voire sa victime, sans rien donner en retour. Ainsi, l’association mycorhizienne est basée sur des bénéfices réciproques et s’articule autour d’échanges bidirectionnels d’éléments nutritifs entre les symbiotes. Les champignons fournissent à la plante mycorhizée une grande partie des minéraux et de l’eau que celle-ci requiert et qu’elle se procurerait bien plus difficilement seule sur sol naturel non fertilisé. La plupart des minéraux se retrouvent dans le sol en quantités limitantes et, de plus, leur mobilité est souvent limitée (Marschner 1990). La structure mycélienne propre aux champignons fournit aux végétaux un bien meilleur accès à ces minéraux que ne le ferait le système racinaire nu. En contrepartie de ces avantages nutritionnels, les plantes alimentent leurs partenaires fongiques en squelettes carbonés. Les champignons sont dépourvus d’appareil photosynthétique et par conséquent hétérotrophes vis-à-vis du carbone (C). Les processus d’échanges de nutriments entre ces partenaires sont toujours demeurés au cœur de toute symbiose mycorhizienne.
Nous comprenons plus aisément les avantages que de telles associations procurent aux plantes dans l’acquisition des ressources minérales en observant l’agencement structural
2 des champignons mycorhiziens. Le mycélium extraradical de ces champignons, organisé en réseaux complexes, prospecte son environnement spatial dans les trois dimensions, de sorte qu’il permet une exploration très efficace du sol; ce que ne peuvent faire les plantes dont le système racinaire est plus limité. La longueur racinaire totale d’une plante est souvent relativement faible et son diamètre relativement gros. En revanche, les hyphes des champignons ont un diamètre beaucoup plus fin et peuvent devenir très longs. Pour un même volume de matière fraîche, la surface représentée par les hyphes fongiques est considérablement plus élevée que celle des racines (Rousseau et al. 1994).
On distingue plusieurs types d’associations mycorhiziennes basés sur le partenaire fongique impliqué dans la symbiose. Les symbiotes végétaux sont si nombreux et si diversifiés qu'une classification selon leur genre serait trop complexe (Smith & Read 1997). Les endomycorhizes à arbuscules constituent les symbioses végétales les plus communes et l’apparition des champignons mycorhiziens à arbuscules (MA) remonterait à une époque située entre 353 et 462 millions d’années (Simon et al. 1993). Une classification récente regroupe les champignons MA sous l’embranchement des Glomeromycota (anciennement Glomales) (Schüssler et al. 2001). Ce sont tous des symbiotes obligatoires incapables de compléter leur cycle vital sans hôte végétal. Malgré leur âge d’apparition très reculé et le fait qu’ils prolifèrent de par le monde, on n’en dénombre sur une base taxinomique qu’un peu plus de 150 espèces (Walker & Trappe 1993). Les champignons MA sont capables de s’associer à des familles d’angiospermes, de gymnospermes, de fougères, de lycopodes et de bryophytes (Smith & Read 1997). Ce sont donc des espèces à large spectre d’hôte, mycorhizant de très nombreuses espèces végétales et qui, par conséquent, peuvent se rencontrer dans des écosystèmes très divers (Morton et al. 1995).
Le terme «arbuscule» caractérisant les champignons MA décrit la structure typique formée par toutes les espèces de cet ordre. Les arbuscules prennent l’apparence d’un arbuste (Gallaud 1905; Brundrett et al. 1984) et on ne les retrouve qu’à l’intérieur des cellules corticales de la racine. Cette particularité permet de caractériser les champignons MA comme endomycorhiziens en opposition aux champignons ectomycorhiziens où les hyphes s’intercalent entre les cellules racinaires de leur hôte sans jamais y pénétrer. Grâce à leur
structure singulière très ramifiée, les arbuscules permettent d’accroître considérablement la surface de contact entre le champignon et la plante. Ceci a largement contribué à reconnaître les arbuscules comme étant le siège privilégié d’échanges de nutriments entre les symbiotes (Cox & Tinker 1976; Gianinazzi et al. 1979; Dexheimer et al. 1985). D’autres structures, les vésicules, se retrouvent chez quatre des six genres que comptent les champignons MA. À l’instar des arbuscules, les vésicules peuvent se retrouver à l’intérieur des cellules corticales mais également intercalées entre celles-ci. De forme variable, elles renferment d’abondants lipides et de nombreux noyaux (Jabaji-Hare et al. 1984; Smith & Read 1997). Il est par conséquent reconnu que les vésicules sont d’importants organes de réserve chez les champignons MA mais peuvent également endosser la fonction de propagule (Biermann & Linderman 1983; Bonfante-Fasolo 1984).
La remarquable amélioration de la nutrition minérale des plantes par les symbiotes fongiques s’accompagne d’un coût pour le partenaire végétal. Ce coût se traduit par la fourniture aux champignons de C dérivant directement des produits de la photo-assimilation (Ho & Trappe 1973). Il semble que le C est transféré de la plante vers le champignon sous forme d’hexoses (Shachar-Hill et al. 1995; Solaiman & Saito 1997). Ces squelettes carbonés sont immédiatement convertis en tréhalose, sucre exclusivement et universellement répandu dans le monde fongique, et en glycogène (Shachar-Hill et al. 1995; Bago et al. 2000b), deux formes de C inexploitables par la plante. Il n’est pas encore certain si l’entrée des hexoses (glucose et fructose) dans les structures fongiques s’accomplit de façon active ou passive (Bago et al. 2000a) mais leur disparition rapide a pour effet d’entretenir un gradient de concentration d’hexoses permanent entre les compartiments racinaires et fongiques, de telle sorte que le champignon se montre constamment demandeur de cette forme de C. D’importantes fractions du tréhalose et du glycogène sont mobilisées pour la synthèse de lipides fongiques. Toutes les structures des champignons MA contiennent des lipides (Sancholle et al. 2001) mais les vésicules et les spores en sont particulièrement riches (Jabaji-Hare et al. 1984; Bécard et al. 1991). Il apparaît clairement ici que le développement du champignon MA de même que la multiplication des structures impliquées dans sa prolifération (vésicules et spores) dépendent en exclusivité du C puisé chez l’hôte végétal. D’un point de vue «métabolisme
4 carboné», le champignon semble être ainsi placé sous la gouverne de la plante. La relation régnant entre les deux partenaires est alors qualifiée de type source/puits: la plante produit et fournit du C, c’est la source; le champignon en demande et en consomme, c’est le puits. Dès lors, le bon déroulement de la symbiose mycorhizienne se révèle intimement régi par ces relations source/puits.
La présente thèse regroupe différents aspects des relations source/puits de C dans la symbiose MA. Elle s’articule principalement autour de l’érable à sucre comme espèce végétale hôte mais d’autres plantes ont également été employées dans nos travaux. L’érable à sucre, Acer saccharum Marsh., est un arbre d’intérêt commercial des régions tempérées strictement colonisé par des champignons MA (Yawney & Schultz 1990; Cooke et al. 1993). Mais surtout, bien que la culture en serres de plantules d’érable à sucre à partir de semences soit erratique, cette essence permet de conjuguer à la fois expériences physiologiques en laboratoire et études éco(physio)logiques de terrain afin de considérer la physiologie du métabolisme carboné dans la symbiose MA à plusieurs échelles. Les différentes expérimentations qui sont exposées dans cet ouvrage se positionnent aux niveaux du système racinaire de plantules, de juvéniles entiers et d’individus matures d’érables à sucre.
Le travail présenté dans cette thèse a débuté en exploitant une des caractéristiques de la symbiose mycorhizienne. Partant du fait que les champignons mycorhiziens n’ont pas (ou peu) de spécificité d’hôte, il est reconnu que les réseaux mycéliens que ceux-ci développent peuvent être connectés à plusieurs plantes de même ou différente espèce. Les hyphes fongiques constituent alors des «ponts» reliant différentes plantes et deviennent ainsi des voies de passage de nutriments potentielles. En laboratoire, des études ont mis en évidence, grâce à l’utilisation de marqueurs isotopiques, des transferts entre plantes de C (Brownlee et al. 1983; Francis & Read 1984; Finlay & Read 1986), d’azote (Arnebrant et al. 1993; Bethlenfalvay et al. 1991) et de phosphore (Newman & Eason 1993; Wittigham & Read 1982) par l’intermédiaire de mycélium de partenaires fongiques communs. Ces mouvements semblent toujours gouvernés par des relations source/puits et dirigées dans ce sens. Cependant, les quantités d’isotopes détectées chez la plante receveuse restent minimes
(Duddridge et al. 1980; Brownlee et al. 1983; Francis & Read 1984; Frey & Shüepp 1992). Ainsi, Newman (1988) s’est interrogé sur la signification écologique et nutritionnelle de ces faibles flux de nutriments entre plantes. Mais dans une étude plus récente, Simard et al. (1997b) ont mesuré des transferts nets de C marqué en quantités significatives entre deux espèces ligneuses à ectomycorhizes (Betula papyrifera et Pseudotsuga menziesii), dont l’une était ombragée dans le but d’établir un puits de C. Une troisième espèce, de type MA (Thuja plicata), présente dans le même espace expérimental paraissait exclue de ce partage des ressources carbonées et ne recevait que de faibles quantités de C marqué.
Certains auteurs considèrent que dans le cas de transferts de C impliquant des champignons MA la plante dite «receveuse» ne reçoit en réalité pas de C (Robinson & Fitter 1999). Ces derniers s’appuient sur le fait que le C transféré chez la plante receveuse a toujours été détecté au niveau de ses racines. Ce C resterait donc uniquement cantonné aux structures fongiques. De plus, le passage de C à partir de champignons MA vers une plante hôte n’a jamais été mis en évidence. Cependant, Bidartondo et al. (2002) ont récemment montré que des plantes épiparasites (plantes non-photosynthétiques qui obtiennent leur C en parasitant des champignons mycorhiziens) étaient mycorhizées par des champignons MA du genre Glomus. Cette découverte suggère fortement que les champignons MA sont capables de fournir du C aux plantes vasculaires et relance l’intérêt de l’étude du transfert de C entre plantes connectées par ce type de champignons.
Un premier chapitre aborde l’étude de la possibilité de transfert de C entre plantes MA en conditions naturelles. Pour ce faire, il convenait d’exclure tout recours à la création artificielle d’une relation source/puits par l’ombrage de la plante receveuse potentielle. Le moteur du transfert de C se devant d’être naturel et de refléter une certaine réalité écologique, il a été opté d’étudier des espèces de phénologie différente se côtoyant naturellement sur le terrain. Les juvéniles d’érable à sucre passent la très grande majorité de leur saison de croissance dans des conditions lumineuses limitantes. Pourtant, elles sont capables de survivre de nombreuses années à ces conditions d’ombre de sous-bois (Canham 1988; Lei & Lechowicz 1990; Ellsworth & Reich 1992). Notre hypothèse était que les érables à sucre juvéniles pouvaient recevoir une fraction de C provenant de plantes mieux
6 exposées à la lumière via les champignons MA. L’érythrone d’Amérique est une plante pérenne de type MA commune des érablières qui, au printemps, tire profit des conditions éphémères de luminosité élevée précédant la fermeture de la canopée. C’est une plante qui à chaque automne renouvelle complètement son système racinaire (Brundrett & Kendrick 1990a; Lapointe & Molard 1997). À la condition que ces deux espèces soit connectées par des champignons MA, leurs différences de phénologie pourraient être à l’origine de l’établissement de relations source/puits. Une première relation aurait lieu au printemps lorsque les juvéniles d’érable à sucre développent leurs feuilles de l’année et que les érythrones sont déjà photosynthétiquement actifs. Une seconde relation s’établirait à l’automne au moment où les juvéniles d’érable à sucre profitent des bonnes conditions lumineuses laissées par la disparition de la canopée et que les érythrones développent leurs nouvelles racines.
La possibilité de transfert de C, en conditions naturelles, entre juvéniles d’érable à sucre et érythrones d’Amérique connectés par des champignons MA étant établie dans le premier chapitre, l’étape suivante a été consacrée à étudier plus précisément la force de puits de C des champignons MA. Les coûts en C attribués à la symbiose MA ont été estimés dans de nombreux travaux (Pang & Paul 1980; Kucey & Paul 1982; Snellgrove et al. 1982; Koch & Johnson 1984; Douds et al. 1988; Wang et al. 1989) et il est aujourd’hui considéré par plusieurs auteurs qu’entre 4 et 20% des photosynthétats d’une plante mycorhizée sont utilisés dans l’entretien des champignons MA (Bago et al. 2000a; Douds et al. 2000; Graham 2000). Les espèces fongiques et végétales utilisées dans les études mentionnées ci-dessus, de même que les méthode de calcul des coûts, divergeaient souvent mais les chiffres avancés (4-20%) témoignent néanmoins d’une grande variabilité. Cela signifie que soit les plantes affichent des capacités de fourniture de C variables entre les espèces, soit les forces de puits des champignons diffèrent d’une espèce (ou d’une souche) à l’autre, soit les deux.
Dans l’état actuel de nos connaissances, aucune étude n’a examiné les forces de puits de C des champignons MA en combinant les facteurs espèce fongique et espèce végétale. Seuls Koch & Johnson (1984) ont étudié les coûts de la mycorhization chez deux espèces
d’agrumes avec une souche de champignon MA. Cet objectif est atteint dans le deuxième
chapitre où les forces de puits de C de trois espèces de champignons MA (Gigaspora
rosea, Glomus intraradices et Glomus mosseae) sur deux espèces d’hôtes végétaux (l’érable à sucre et l’orge) ont été comparées.
Le troisième chapitre s’inscrit dans la continuité du chapitre précédent qui avait révélé que, contrairement aux deux autres espèces de champignon MA testées, G. mosseae ne se comportait jamais comme un puits fort, ni avec l’érable à sucre, ni avec l’orge. Afin de vérifier si ce caractère est spécifique à G. mosseae ou propre à la souche utilisée précédemment, trois souches de ce champignon d’origines géographiques diverses ont été testées sur l’orge.
Enfin, le quatrième chapitre, se positionne dans un contexte de perturbation naturelle affectant le houppier d’érables à sucre matures. En janvier 1998, une vague de verglas de forte ampleur frappe le sud du Québec et l’est de l’Ontario dévastant les érablières de ce secteur à un niveau de dommages sans précédent (Irland 1998). La grande majorité des arbres adultes touchés voient ainsi la partie supérieure de leur tronc brisée et leur cime presque totalement détruite. Au lendemain de ce sinistre, le Ministère des Ressources Naturelles du Québec (MRN) a effectué un suivi des érablières atteintes par le verglas dans des parcelles permanentes situées à l’intérieur de la zone dévastée. Des relevés réalisés sur chacun des arbres adultes de ces parcelles ont permis de dresser un «inventaire aérien» des érablières endommagées. Le volume de houppier vivant, exprimé en pourcentage du volume de houppier total théorique, a été noté aux étés 1998 et 1999. En comparant les valeurs de volumes de feuillage obtenues au cours des deux années de croissance qui ont immédiatement suivi le verglas, on a pu caractériser comment les arbres ont réagi au stress induit par cette perturbation. Ainsi, on a pu distinguer des individus ayant eu une bonne reprise de croissance à la suite du verglas (augmentation du volume de houppier) d’autres individus qui en ont eu une mauvaise (pas de changement, ou diminution, du volume de houppier). Curieusement, deux érables vivant côte à côte, ayant subi les mêmes niveaux de dommages et ayant approximativement la même taille (diamètres de troncs semblables) peuvent avoir réagi au verglas de façon totalement différente. L’un peut présenter une très
8 bonne reprise de croissance alors que son plus proche voisin en affiche une mauvaise (observations personnelles). Pour quelle(s) raison(s) la croissance de certains arbres a bien redémarré après le verglas leur permettant de reconstituer leur cime alors que d’autres ont eu plus de difficulté à faire de même et semblent végéter? Quels sont les facteurs influençant ce phénomène?
Plusieurs raisons expliquant ces différences de reprise de croissance peuvent être avancées: la vigueur des arbres avant le verglas, la présence de carie ou encore des facteurs génétiques. Cependant, une autre cause doit être prise en considération dans l’étude de l’état de santé post-verglas des érables à sucre: le fait que ces derniers vivent en symbiose avec des champignons MA. La destruction du feuillage des érables à sucre touchés par le verglas a entraîné une baisse de la fourniture de C alors que les champignons MA étaient déjà installés dans les racines de ces arbres. Les coûts d’entretien de ces champignons sont alors devenus proportionnellement plus élevés.
Comme il l’a déjà été montré dans les chapitres 2 et 3, les coûts d’entretien de ces champignons MA peuvent varier selon l’espèce. Il se pourrait qu’un arbre adulte n’héberge qu’un nombre limité de champignons mycorhiziens. En effet, une fois qu’un champignon a mycorhizé une plante hôte, celui-ci a tendance à s’y maintenir et à gêner de nouvelles colonisations par d’autres espèces (Pearson et al. 1993; Pinior et al. 1999; Vierheilig et al. 2000a) Ceci aurait pour effet de restreindre le nombre de champignons colonisant une même plante. Il est donc possible qu’un arbre soit mycorhizé en majorité par des espèces moins bénéfiques et plus coûteuses en C. À la suite d’un événement aussi stressant pour un arbre que la perte de sa cime, la composition de ses partenaires fongiques pourrait en influencer la reconstitution. Si un arbre est associé à des champignons coûteux en C et que ceux-ci continuent de représenter un puits de C important après la perte de la cime, cet arbre pourrait avoir moins de ressources à sa disposition pour régénérer son feuillage. Par conséquent, il serait intéressant d’examiner s’il existe une corrélation entre l’abondance et la composition en champignons MA que l’on retrouve dans la rhizosphère des érables endommagés par le verglas de 1998 et leur capacité de reprise de croissance.
L’ensemble de ces différentes études et des résultats qui en ont découlé a mené à l’élaboration d’une synthèse qui est présentée dans la conclusion générale. Quelques perspectives de recherche sont également proposées dans cette discussion.
CHAPITRE I
14
C transfer between the spring ephemeral Erythronium
americanum and sugar maple saplings via arbuscular
mycorrhizal fungi in natural stands
1.1 Avant-Propos
Le texte de ce chapitre a été publié dans la revue Oecologia (Lerat, Gauci, Catford, Vierheilig, Piché & Lapointe 2002). Rachel Gauci a participé au marquage au 14C sur le terrain, Jean Guy Catford a collaboré à l’analyse de la radioactivité et Horst Vierheilig a contribué à la rédaction de la discussion. Une partie de ce chapitre a fait l’objet d’une présentation orale au Colloque du Centre de Recherche en Biologie Forestière, Université Laval, (Québec, Canada) qui s’est déroulé les 22 et 23 février 2002 ainsi que d’une affiche présentée au congrès Mycorhize 2001 tenu à Montréal (Canada).
1.2 Résumé
Le transfert de carbone (C) entre des juvéniles d’érable à sucre (Acer saccharum) et une éphémère de printemps, l’érythrone d’Amérique (Erythronium americanum) via le mycélium de champignons mycorhiziens à arbuscules (MA) a été étudié sur le terrain. Des juvéniles d’érable à sucre et des plants d’érythrone ont été placés ensemble dans des pots placés dans le sol d’une érablière en 1999. Des bouleaux jaunes ectomycorhiziens (Betula aleghaniensis) y ont été ajoutés comme plantes témoins. Au printemps 2000, durant l’expansion foliaire des juvéniles d’érable à sucre, les feuilles d’érythrone ont été marquées au 14CO2. Sept jours après le marquage, la radioactivité a été détectée dans les feuilles, la
était 13 fois plus élevée que chez les bouleaux jaunes révélant l’existence d’un transfert direct de 14C entre les plantes MA. La quantité de 14C transférée aux juvéniles d’érable à sucre était négativement corrélée au pourcentage de 14C alloué à l’organe de réserve de l’érythrone. Un second marquage a été réalisé à l’automne 2000 sur les feuilles d’érable à sucre durant la croissance annuelle des racines d’érythrone. De la radioactivité a été détectée chez 7 des 22 systèmes racinaires d’érythrone mais était absente chez les bouleaux jaunes. Ces résultats suggèrent que les champignons MA connectant différentes espèces de sous-bois peuvent agir comme des ponts de transfert de C réciproque entre les espèces végétales en relation avec la phénologie des plantes impliquées.
1.3 Abstract
We investigated in the field the carbon (C) transfer between sugar maple (Acer saccharum) saplings and the spring ephemeral Erythronium americanum via the mycelium of arbuscular mycorrhizal (AM) fungi. Sugar maple saplings and E. americanum plants were planted together in pots placed in the ground of a maple forest in 1999. Ectomycorrhizal yellow birches (Betula alleghaniensis) were added as control plants. In spring 2000, during leaf expansion of sugar maple saplings, the leaves of E. americanum were labelled with
14CO
2. Seven d after labelling, radioactivity was detected in leaves, stem and roots of sugar
maples. Specific radioactivity in sugar maples was 13-fold higher than in yellow birches revealing the occurrence of a direct transfer of 14C between the AM plants. The quantity of
14C transferred to sugar maple saplings was negatively correlated with the percentage of 14C
allocated to the storage organ of E. americanum. A second labelling was performed in autumn 2000 on sugar maple leaves during annual growth of E. americanum roots. Radioactivity was detected in 7 of 22 E. americanum root systems and absent in yellow birches. These results suggest that AM fungi connecting different understorey species can act as reciprocal C transfer bridges between plant species in relation with the phenology of the plants involved.
12
1.4 Introduction
Mycorrhizal fungi are symbiotic partners associated with the great majority of land plant species (Smith & Read 1997). They colonise roots and improve plant nutrition mainly by transferring phosphate (P) from the soil to the plant. The plants provide the fungi with carbohydrates (Smith & Read 1997). Because of low host specificity of mycorrhizal fungi their mycelium can form a network that interconnects host plants of the same (Brownlee et al. 1983; Newman & Eason 1993; Graves et al. 1997) or different species (Francis & Read 1984; Arnebrant et al. 1993). This means fungal hyphae can provide pathways to transfer of compounds between mycorrhizal plants. Isotope tracer studies have documented transfer of water (Duddridge et al. 1980) and nutrients such as P (Newman & Eason 1993), N (Arnebrant et al. 1993) and C (Francis & Read 1984; Finlay & Read 1986).
The occurrence of direct C transfer from donor to receiver plants through fungal mycelium has been mostly studied under laboratory and greenhouse conditions (Francis & Read 1984; Finlay & Read 1986; Simard et al. 1997a) with arbuscular mycorrhizal (AM) and ectomycorrhizal (ECM) fungi. On most of these studies receiver plants were artificially shaded demonstrating that C transfer is strongly governed by a source-sink relationship. Nevertheless, physiological importance of such transport in ecosystems has not been proven (Robinson & Fitter 1999). Brownlee et al. (1983) hypothesised that newly germinated seedlings of Pinus sylvestris could receive photoassimilates from mature trees in their vicinity and overcome forest understorey shade conditions. Recently, Simard et al. (1997b) measured net transfer of C in the field between seedlings of two ECM tree species Betula papyrifera and Pseudotsuga menziesii. There was a much smaller C transfer (18% of that between the ECM species) towards AM tree seedlings of Thuja plicata present in the vicinity. They also found that the amount of C exported from the donor plant was positively correlated with the level of shading on the receiver plant. To our knowledge, C transfer between species with different developmental phenology has not been reported within a plant community.
In natural plant communities dominated by endomycorrhizal species, AM fungi rapidly colonise roots of newly germinated seedlings and plants that renew their entire root system
every year, for example Liliaceous spring ephemerals. Spores of AM fungi are an improbable inoculum because of their low viability in the soil (Read et al. 1976; Zahka et al. 1995). Therefore, the most likely source of inoculation of new roots is hyphae from the roots of neighbouring plants previously colonised by AM fungi.
Erythronium americanum Ker-Gawl (trout lily) is an abundant spring ephemeral in maple forests of North America that is colonised by AM fungi (Brundrett & Kendrick 1990b; Lapointe & Molard 1997). In the region of Québec City, epigeous development of E. americanum follows snow melt in late-April when the tree canopy is completely open (Fig. 1.1a, b). E. americanum takes advantage of these full light conditions for active carbohydrate assimilation until canopy closure. Carbohydrate storage in the corm (underground reserve organ) is completed three wk after leaf expansion but high levels of photosynthesis (10-14 µmol CO2 m-2 s-1) are maintained until the overstorey canopy starts
to close (Taylor & Pearcy 1976; Lapointe, unpublished data). Leaves of E. americanum then senesce during canopy closure. Leaf senescence is completed by mid-June followed by root senescence at the beginning of July (data not shown). In early autumn (mid-September) E. americanum produces new unbranched roots that are rapidly colonised by AM fungi (Brundrett & Kendrick 1990a; Lapointe & Molard 1997).
Sugar maple (Acer saccharum Marsh) is a common North American tree species associated with AM fungi (Yawney & Schultz 1990; Cooke et al. 1993). Bud burst of sugar maple saplings occurs about three wk after leaves of E. americanum have unfolded and started to photosynthesise whilst bud burst of mature sugar maple trees is delayed by two wk (Fig. 1.1a). Thus, the expansion of the leaves of sugar maple saplings occurs under high light conditions. Leaf development is a costly process in terms of carbohydrates (Poorter & Villar 1997) and sugar maple saplings could receive C during leaf expansion through mycorrhizal hyphae from neighbouring plants already photosynthetically active.
14
Figure 1.1. Phenology of Erythronium americanum and leaves of sugar maple saplings and mature trees (a) and relative PAR (photosynthetically active radiation) on the ground of a maple forest (b) over a growing season in the region of Québec City. Grey bars represent development (beginning of line) or senescence (end of line) of leaves. Dates of labelling with 14CO2 are represented by arrows.
(a) 14CO 14CO Aug. (b) PAR 14CO 2 presence of leaves
epigeous growth root growth
E. americanum
Sugar maple, saplings
Sugar maple, mature trees
Nov. Oct. Sep. Jul. Jun. May Apr. presence of leaves
In the autumn, mycorrhizal colonisation of E. americanum roots occurs during root growth, which represents a significant carbohydrate cost at a time when plants do not have leaves (Lapointe & Molard 1997). C required for root production in E. americanum could be partly supplied by photosynthetically active neighbouring sugar maple trees and juveniles via interconnecting mycelium.
The goal of this work was to test for reciprocal and temporal transfers of C between the spring ephemeral E. americanum and 1-year old saplings of the woody species sugar maple in a natural plant community. Such net transfers would be restricted in time to the period when one species is photosynthetically active while the other is not.
1.5 Materials and methods
1.5.1 Study site
The experiments were carried out in a sugar maple forest in Saint-Augustin-de-Desmaures (46°48’N, 71°23’W) in the vicinity of Québec City. The site has been previously described in Lapointe & Molard (1997). The canopy is dominated by Acer saccharum Marsh with A. rubrum L., Fraxinus americana L. and Ulmus rubra Mühl as companion species. The soil is a clay-loam with a thin moder type humic horizon. The most abundant understorey species are Erythronium americanum Ker-Gawl, Trillium erectum L. and Veratrum viride Ait.
1.5.2 Experimental design
In spring 1999 (mid-May), 60 organic fibre pots (v=3 1, d=16 cm, h=16 cm) were buried into the soil at the study site to a depth allowing the edge to remain 1-2 cm over ground level. Pots were filled with natural soil collected at the site. The soil included fine roots of mature trees as source of mycorrhizal inoculation for sugar maple seedlings. One year-old seedlings of sugar maple were carefully collected near the experimental site and transplanted into the middle of each pot (one seedling per pot). Five wk later, three corms of E. americanum were planted 5 cm deep into each pot. At that time, leaves of the
16 transplanted E. americanum had already senesced and corms had entered summer dormancy. In August, yellow birch saplings (Betula alleghaniensis Britton) (<12-cm high) were collected from a distant site at l’Île-aux-Grues (47°03’N, 75°28’W). Birches are typically colonised by ECM fungi rather that AM fungi. Presence of ECM features was visually checked and seedlings with the highest mycorrhization were transplanted into the pots (one per pot). Pots were regularly removed from soil and immediately replaced to avoid possible intrusion of roots from surrounding trees.
1.5.3 Labelling experiments with
14C
1.5.3.1 Spring labellingIn mid-May 2000, at bud burst of sugar maple and yellow birch saplings (Fig. 1.1a), 12 of the pots were randomly selected for 14C labelling. The leaf of one of the three E. americanum plants in the selected pots was enclosed in a 3.7-l transparent freezer bag (Ziploc®) that was firmly anchored between two wooden stakes. A 12-cm-rubber tube (d=5 mm) was fixed to an upper corner of the bag and tightly closed during the experiment. A sealing compound placed around the stem of the plant insured hermetic closure of the bag during labelling. A 29.5-ml cup containing a basic solution of 185 kBq (5 µCi) NaH14CO
3
(Amersham Pharmacia Biotech) was placed inside the plastic bag and the bag tightly closed. Pulse-labelling started when 1 ml lactic acid (85%) was syringe-injected through the bag into the cup to liberate gaseous 14CO2 (the release of 14CO2 in the bag was
estimated to increase the CO2 concentration by 4-6 ppm). Plants were left exposed to
sunlight, >1000 µmol m-2 s-1 PAR (photosynthetically active radiation), for 1.5 h. At the
end of the experiment, excess 14CO2 was evacuated through the rubber tube and trapped in
a 1 N NaOH solution.
After a 7-d chase period, which allowed complete expansion of sugar maple leaves, all plants in the pots were harvested and dried at 65°C for 24 h except for roots of non-labelled E. americanum plants, which were collected for determination of mycorrhizal colonisation.
1.5.3.2 Autumn labelling
In the autumn 2000, two consecutive labelling sessions were performed (Fig. 1.1a). In mid-September, 12 pots were chosen for labelling sugar maples. At this time in early autumn, leaves of sugar maple saplings were still green and capable of photosynthesis (assimilation rate: 1.7 ± 0.8 µmol CO2 m-2 s-1) while E. americanum corms had no functional roots. This
early session insured labelling of the sugar maple saplings before leaf senescence. The technical procedure was identical to that used in spring with the following modifications. Pots were moved to an open field where ambient sunlight reached 1200 µmol m-2 s-1 PAR
and the whole foliage of sugar maple 1-year saplings was pulse-labelled in plastic bags for 2 h. Three wk later, root samples harvested in other pots showed that E. americanum corms had formed new roots and these had been colonised by AM fungi. At this time leaves of the previously labelled sugar maples were still green, and the plants were labelled a second time. Pots were harvested two wk after the second labelling session (14-d chase period). Plant material was destructively harvested and dried with exception of all E. americanum roots which were stored in a FAA solution.
1.5.4 Determination of radioactivity
1.5.4.1 Spring labellingDried sugar maple, non-labelled E. americanum (root excepted) and yellow birch were exposed to autoradiography film (Kodak BioMax). The films were developed after six wk of exposure.
Leaves, stem and roots of sugar maple saplings and labelled E. americanum and leaves of non-labelled E. americanum were separated and ground in liquid nitrogen likewise whole yellow birch saplings. An aliquot of each material type (15-50 mg) was digested as in Clifford et al. (1973) and radioactivity determined by liquid scintillation spectrometry. Counts were standardised with a quench curve and radioactivity was expressed in dpm.
18
1.5.4.2 Autumn labelling
Yellow birch saplings and dried corms of E. americanum were exposed to autoradiography film. The films were developed after six wk of exposure.
Roots samples of E. americanum were fixed in paraffin and 5-µm thick sections were mounted on microscope slides. Paraffin was then dissolved with toluene and root sections were coated with LM-1 nuclear emulsion (Amersham Pharmacia Biotech; crystal size of 0.2 µm). The emulsion was processed after exposure at 4°C for two to four wk. The rest of E. americanum root samples were rinsed several times with distilled water, dried, weighed and assessed for radioactivity as described previously, using liquid scintillation. Radioactivity in yellow birch saplings was also assessed.
1.5.5 Mycorrhizal colonisation levels
Roots of E. americanum were cleared in a 10% (w/v) KOH solution for 12 min at 90°C prior to staining with a trypan blue solution (Koske & Gemma 1989). AM colonisation was assessed microscopically at 78× magnification using the method described by Trouvelot et al. (1986). The following parameters were calculated: frequency of colonisation, intensity of colonisation and arbuscular content.
1.5.6 Statistical analysis
One-way ANOVA was performed to compare radioactivity data in sugar maple and yellow birch saplings after spring labelling using the GLM procedure of the SAS statistical package.
1.6 Results
Following spring radiolabelling of E. americanum plants, the macro-autoradiography revealed traces of 14C in all sugar maple saplings (Fig. 1.2a-c). In contrast, no labelling was observed in yellow birch saplings and non-labelled E. americanum growing in the same pots as the labelled E. americanum (autoradiographs not shown), indicating absence of
transfer between E. americanum plants. Roots, stem or leaves of the sugar maple saplings were labelled with 14C. Five of the saplings were extensively labelled in the leaves. Two of the saplings showed only slight labelling in their roots.
Total radioactivity (dpm) and specific radioactivity (dpm g-1 dry matter) were much greater in sugar maple compared to yellow birch saplings (P<0.01). Average total radioactivity in sugar maple saplings was 738 ± 642 dpm (means ± 1 SD; range, 136 to 2233 dpm) and was 15 ± 26 dpm (range, 0 to 82 dpm) in yellow birch saplings. Average specific radioactivity was 3884 ± 3046 dpm g-1 dry matter in sugar maple saplings and was 290 ± 450 dpm g-1
dry matter in yellow birch saplings. Small amounts of radioactivity (224 ± 318 dpm g-1 dry
matter) were detected in the leaves of non-labelled E. americanum indicating that very few
14C was absorbed by photosynthetically active leaves during the chase period. In sugar
maple saplings specific activity was the highest in leaves (Table 1.1). Radioactivity was thus mainly localised in the shoot (leaves and stem), with less radioactivity measured in the roots (Table 1.1). In labelled E. americanum the 14C content was 1.12×106 ± 0.22×106 dpm
(18.6 ± 3.6 kBq) which was equivalent to ca. 10% of the 14CO
2 used for labelling. On
average, 53.9 ± 30.4% of the total was found in the corm of labelled E. americanum plants.
The total activity in the sugar maple saplings was 0.064 ± 0.049% of that in E. americanum while dry biomass was 195 ± 76 mg for sugar maple saplings and 292 ± 96 mg for E. americanum. It appeared that when the percentage of 14C in the corm of labelled E. americanum was the lowest, the 14C contents in sugar maple saplings was the highest. Consequently, a negative correlation (r2=0.61, P=0.0028) was established between the percentage of 14C translocated to the corm of labelled E. americanum and the percentage of
14C translocated to sugar maple saplings (Fig. 1.3).
The root systems of non-labelled E. americanum plants showed high levels of root colonisation by AM fungi in spring. Frequency of colonisation was 44.7 ± 27.0% (n=28), intensity of colonisation was 13.3 ± 14.7% and arbuscular content was 84.8 ± 19.6%.
20
Figure 1.2. Macro-autoradiographs of leaves (a and b) and stem and root system (c) of
labelled C in receiver sugar maple saplings. Dry plant material was exposed for six wk on Kodak scientific imaging films. Bars represent 1 cm.
(b) (a)
Table 1.1. Specific activity (dpm g-1 dry matter), biomass ratio, and distribution of radioactivity in leaves, stem, and roots of sugar maple saplings one wk after spring labelling. Data represent means ± 1 SD.
Leaves Stem Roots
Specific activity 6186 ± 5953 2386 ± 2552 2941 ± 4360 Biomass ratio 30.8 ± 11.4% 36.6 ± 13.4% 32.6 ± 7.4% % radioactivity 50.4 ± 24.6% 27.6 ± 16.3% 22.0 ± 16.9%
In autumn, after sugar maple labelling, 14C was detected with liquid scintillation in 7 of 22 E. americanum root samples. The average total radioactivity was 230 ± 269 dpm (range, 43 to 645 dpm) and the average specific radioactivity was 4244 ± 5288 dpm g-1 dry matter for
the 7 labelled root systems. No radioactivity was detected in the yellow birch saplings. 14C content in labelled sugar maple saplings was 1.12×106 ± 0.51×106 dpm (20.8 ± 8.5 kBq),
which was less than 6% of 14CO2 used for labelling the plants. The total activity in the
seven labelled E. americanum root systems was 0.018 ± 0.021% of that in sugar maple saplings.
Radioactivity was not detected on autoradiographs after six wk of exposure in E. americanum corms, or in yellow birch saplings. Microscope slides of E. americanum roots showed no trace of radioactivity after two to four wk of exposure to LM-1 nuclear emulsion. After four wk, the background was too high to detect 14C in the root and fungal tissues.
1.7 Discussion
This is the first report, to our knowledge, of source-sink relationships that were not governed by shading the receiver plants but by the phenology of receiver and donor plants.
22
14C translocated to corm/total 14C in E. americanum
0.0 0.2 0.4 0.6 0.8 % 14 C transloc ate d t o suga r m apl es 0.00 0.04 0.08 0.12 0.16 0.20
Figure 1.3. Relationship between the percentages of 14C exported to sugar maple saplings following spring labelling of Erythronium americanum and the fraction of 14C in the corm of labelled E. americanum.
In our experiment autoradiographs and quantitative analyses of radioactivity revealed 14C in all sugar maple saplings. Sugar maple saplings showed labelling not only in roots but also in leaves and stem clearly demonstrating that (i) 14C transfer from the AM fungi to the receiver plants occurred and (ii) the transferred 14C was translocated into the plant and thus available for plant growth. Earlier C transfer studies reported 14C only in the roots of the receiver plants (Francis & Read 1984, Finlay & Read 1986, Wu et al. 2001). However, because 14C could be confined to the AM fungal tissues colonising the roots, labelled C in the shoot is clear evidence for C transfer from a donor plant via the mycorrhizal mycelium to a receiver plant.
We found no visible radioactivity in yellow birches and non-labelled E. americanum on autoradiographs (data not shown) indicating that absorption of 14C-root exudates by ECM and AM fungi was insignificant. The low levels of 14C detected in the quantitative analyses
of 14C of yellow birch showed that passive absorption of radio-labelled material could
occur. However, average 14C specific activity in sugar maple saplings was 13-fold higher than in yellow birch saplings while the lowest 14C specific activities in sugar maples were comparable with yellow birches. It is likely that in the low activity sugar maples the 14C detected in those saplings was due to passive absorption of labelled root exudates by associated AM fungi or the saplings roots.
The fraction of 14C fixed (and retained) by the E. americanum and delivered to the sugar maples saplings (0.064% ± 0.049) seems tiny. However, we have estimated that over the 7 d of leaf expansion of sugar maple saplings with optimal sunlight conditions, an E. americanum plant with photosynthetic rates of 12 µmol CO2 m-2 s-1, 6 h per d, would
transfer up to 21 µg C to sugar maples. From results of another study (unpublished data) where starch and sucrose contents were measured in sugar maple saplings in late autumn after bud formation and spring we have assessed the cost of leaf expansion in our sugar maple sapling was about 1.32 mg C per sapling. Thus, the C transferred from one E. americanum could cover up to 1.6% of leaf expansion of 1-year sugar maple saplings. It is likely that in undisturbed ecosystems sugar maple saplings would be connected to several E. americanum plants, as E. americanum is often more abundant than sugar maple saplings
24 (personal observation). We can thus expect that sugar maple saplings would receive more C than measured in our one-to-one labelling experiment, and that the C contribution of E. americanum during sugar maple sapling leaf expansion is physiologically significant.
The distribution of radioactivity in sugar maple saplings was consistent with the concept of source-sink relationships in C transfers between donor and receiver plants. Seven d after the exposure of potential donor plants to 14C, the label had been transferred to the sugar maple saplings and half of it was localised in leaves, which represented less than a third of the total sapling biomass. This labelling pattern would be a consequence of bud burst in sugar maple saplings which occurred at the time the labelling experiment was performed. New leaves development acts as a strong sink for carbohydrates and thus directs high quantities of C towards these growing organs. Our results are consistent with those of Francis & Read (1984), Finlay & Read (1986) and Simard et al. (1997b) who underlined the essential role of source-sink relationships between plants connected by common mycelia. C transfer between plants was modified by applying different levels of irradiance to the receivers.
Robinson & Fitter (1999) argued that there was as yet no evidence of C transfer from AM fungi to the shoots of the receiver plant. They suggested that the low levels of 14C detected in the shoots of receiver plants (e.g. Francis & Read 1984) originated from 14CO2 respired
by roots and microbes and re-fixed during photosynthesis by the receiver plant. In our experiment, labelling was performed at bud burst of the receivers and the chased period (7 d) ended after leaf expansion of sugar maple. Photosynthetic 14CO2 fixation would not
explain our findings as photosynthetic rates are restricted in developing leaves, as in Acer campestre (Küppers 1984). Furthermore, the quasi-absence of radioactivity in non-labelled E. americanum, which would have had elevated photosynthetic rates during the chase period (Taylor & Pearcy 1976), argues against the possibility of net 14CO2 fixation by
photosynthesis in the receiver plants.
The close correlation between C allocated to the corm of E. americanum and C translocated to sugar maple saplings suggests that E. americanum export C once its own carbohydrate
storage is completed emphasising again the importance of source-sink relationship in C transfer. At the time of labelling, eight of the 12 E. americanum had apparently not completely filled up their starch reserve, while three E. americanum plants were translocating small amounts of C to the corm, suggesting that the storage process was about to end in the former eight E. americanum. We can estimate that if the labelling of E. americanum would have occurred 2-3 d later, more E. americanum would have then completed their storage of carbohydrates and we would have observed higher 14C movements towards sugar maple saplings. The magnitude of C transfer between E. americanum and sugar maple saplings is probably dependent on the climatic conditions prevailing at spring between the unfurling of E. americanum leaves and the canopy closure.
In spring, the E. americanum plants were highly mycorrhizal. The colonisation levels were, however, less than reported by Brundrett & Kendrick (1990b) and by Lapointe & Molard (1997) who found mycorrhizal levels of 67% and 82.8% respectively. Nevertheless, the presence of AM fungi in every tested plant reflected a sufficient inoculum potential in the experimental pots.
In autumn, the 14C detected in 7 of the 22 E. americanum root systems strongly supports the notion that AM fungi living on sugar maple roots can be a source of mycorrhization of E. americanum roots. If the source of activity had been sugar maple root exudates then all E. americanum would have been equally labelled. Therefore, we concluded that about one third of E. americanum plants were connected to the donor sugar maples in autumn 2001.
Notwithstanding, autumn labelling did not demonstrate whether C was transferred from sugar maple saplings to E. americanum root cells. Micro-autoradiographs with nuclear emulsion did not show any traces of radioactivity. The quantity of 14CO2 we used for
labelling donor plants (2×185 kBq per plant) was very low compared to micro-autoradiography of Bücking & Heyser (2001) who used 7.4 MBq (200 µCi) 14CO
2 per
plant. In our work, 14C detection threshold levels were below background. Hence, radioactivity in fungal and root cells was not visually documented.
26 Possible transfer of C in autumn from sugar maple saplings to E. americanum for the root formation was less clear than the spring case of C transfer from E. americanum to maple saplings. However, traces of 14C were detected in some of the E. americanum plants, suggesting a possible payback of C from sugar maples to E. americanum in autumn.
In conclusion, we have documented C transfer between sugar maple saplings and the spring ephemeral E. americanum connected by AM mycelium that appears reciprocal. An initial transfer occurs in spring from E. americanum to sugar maple saplings during leaf expansion of the saplings. A reciprocal transfer is highly likely in autumn. This second transfer was from the sugar maple saplings to newly developed roots of E. americanum plants, but we do not have conclusive proof at this point that imported C was used for the formation of E. americanum roots. Reciprocal transfers could be frequent in natural ecosystems where plants with different phenologies share AM fungi.
1.8 Conclusion
Ce chapitre fait pour la première fois la preuve de la possibilité à l’état naturel d’échanges bidirectionnels de C entre plantes de phénologie différente connectées par des champignons MA. Un premier transfert a lieu au printemps à partir d’érythrones d’Amérique photosynthétiques vers des juvéniles d’érable à sucre au moment de leur développement foliaire. L’amplitude de ce transfert de C est corrélée au niveau de stockage en amidon de l’organe de réserve des érythrones (corme). Il apparaît donc que l’érythrone ne délivre du C aux juvéniles d’érable à sucre par le biais de champignons MA qu’une fois le remplissage de son corme complété. Un second transfert semble très probable à l’automne à partir des juvéniles d’érable à sucre vers les racines d’érythrone nouvellement formées. Cependant, il a été impossible de prouver que le C importé a été utilisé dans la construction des racines d’érythrone. De tels transferts de C pourraient être fréquents dans les écosystèmes naturels où des plantes de phénologie différente partagent des champignons MA communs.
CHAPITRE II
Carbon partitioning in a split-root system of arbuscular
mycorrhizal plants is fungal and plant species dependent
2.1 Avant-propos
Ce chapitre a été publié par Lerat, Lapointe, Gutjahr, Piché & Vierheilig (2003) dans la revue New Phytologist. Horst Vierheilig a initié et a participé à la phase expérimentale de ce travail. Sylvain Gutjahr a réalisé l’étude de croissance de plants d’orge en serres. Le contenu de ce chapitre a également été présenté oralement au Colloque Canadien sur les Mycorhizes tenu en avril 2002 à Québec (Lerat, Lapointe, Piché & Vierheilig) et une partie a fait l’objet de la présentation d’une affiche au congrès Plant Biology 2001 tenu à Providence, Rhode Island, USA (Lerat, Lapointe, Piché & Vierheilig).
2.2 Résumé
La répartition racinaire de carbone (C) a été étudié chez deux espèces végétales hôtes colonisées par une parmi trois espèces de champignon MA. Des systèmes à racines dédoublées («split-root») d’orge (Hordeum vulgare) et d’érable à sucre (Acer saccharum) ont été mycorhizés d’un côté avec l’un des trois champignons MA. Trois semaines après inoculation, leurs feuilles ont été marquées à l’aide de 14CO2. Les plantes ont été récoltées
24 heures plus tard et le 14C a été analysé séparément dans les systèmes racinaires des côtés
mycorhizés (M) et non mycorhizés (NM). La répartition du 14C entre les côtés M et NM
variait en fonction de l’espèce fongique et de la plante hôte utilisées. Gigaspora rosea était un puits de C de forte capacité avec les deux espèces végétales, Glomus intraradices était un puits de forte capacité avec l’orge alors que Glomus mosseae ne modifiait la répartition