The yeast transcription elongation factor Spt4/5 is a sequence-specific RNA binding protein
Texte intégral
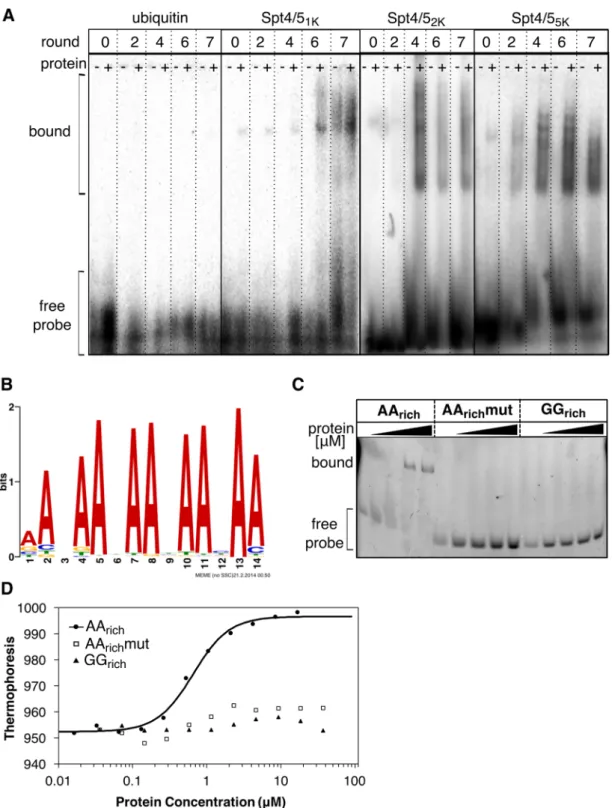
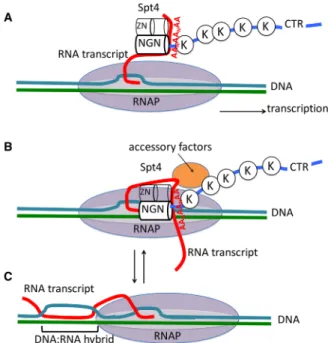
Figure
Documents relatifs
using other records in the lake sediment, including pollen (vegetation change), diatoms (lake water pH and chemistry), SOOt (coal combustion), charcoal (wood combustion),
Complétez les phrases suivantes avec le présent simple ou le présent
If the breast is pendulous elevate the breast (infra-mammary area/undersurface of the breast). Notice any changes in the breast skin such as dimpling, puckering, or asymmetry..
Various behavioral techniques have been employed and researched in terms of enhancing the social behavior of children with ASD such as: trial training, making use of stereotypic
Instead, Peterson’s theorem should (only) be interpreted in the following way: it formally shows that, given two other technical premises, the Archimedian condition and (as I argue
ASSOCIATION OF CANADA LANDS SURVEYORS - BOARD OF EXAMINERS WESTERN CANADIAN BOARD OF EXAMINERS FOR LAND SURVEYORS ATLANTIC PROVINCES BOARD OF EXAMINERS FOR LAND SURVEYORS ---..
Since the given 3-dimensional P-Sasakian manifold is of constant scalar curvature r = 6 , therefore, by virtue of Theorem(4.1), it implies that it is locally φ
The antibiotic ointment (tetracycline) has to be applied relatively frequently and repeatedly for many years in order to prevent blinding disease.. It is, therefore,
