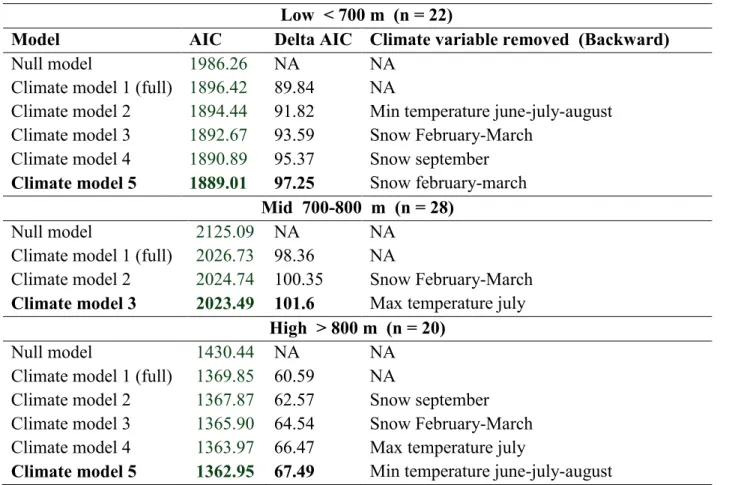
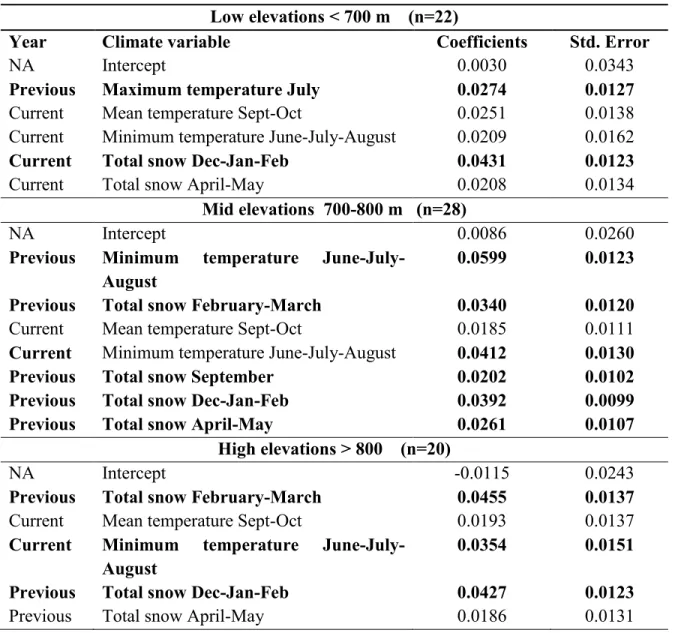
La limite de répartition supérieure de l'érable à sucre et du bouleau jaune sous contrôle climatique: étude dendroécologique le long d'un gradient d'élévation
79
0
0
Texte intégral
(3)
(4)
(5)
(6)
(7)
(8)
(10)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(30)
(31)
(32)
(33)
(34)
(35)
(36)
(37)
(38)
(39)
Figure




+6



Outline
Documents relatifs
