© Maxime Perron, 2020
Bases neurobiologiques de la perception de la parole
dans le bruit chez les chanteurs et les non-chanteurs
Mémoire
Maxime Perron
Maîtrise en sciences cliniques et biomédicales - avec mémoire
Maître ès sciences (M. Sc.)
Bases neurobiologiques de la perception de la
parole dans le bruit chez les chanteurs et les
non-chanteurs
Mémoire
Maxime Perron
Sous la direction de :
ii
Résumé
Percevoir la parole dans le bruit devient difficile en vieillissant. La nature de ces difficultés est incertaine, mais le déclin lié à l’âge de la structure de la voie dorsale de la parole pourrait être un facteur contributif. La pratique du chant choral pourrait aider à ralentir le vieillissement de la voie dorsale de la parole, mais les effets du chant choral sont peu connus. Les objectifs de ce mémoire étaient d’investiguer la relation entre le déclin structurel de la voie dorsale et les difficultés de perception de la parole, et d’identifier comment la pratique du chant choral modifie cette relation. Dans le cadre de ce mémoire, 44 chanteurs et 41 non-chanteurs jeunes et âgés ont complété une séance d’imagerie par résonance magnétique, ainsi que des évaluations cognitives et auditives. Un sous-groupe a effectué une tâche de discrimination de syllabes dans le bruit. Dans une première étude, des avantages comportementaux ont été observés pour des chanteurs âgés avec certaines caractéristiques liées à leur pratique, ce qui suggère que le chant bénéficie à la perception de la parole seulement dans certaines conditions. Ces avantages ont été associés à la structure corticale de régions de la voie dorsale de la parole. Dans une deuxième étude, le déclin structurel de segments fronto-temporaux du faisceau arqué, qui constitue l’un des principaux faisceaux de matière blanche de la voie dorsale de la parole, a été associé aux difficultés de perception de la parole. De plus, des différences dans la structure du faisceau arqué ont été identifiées entre les groupes, mais ces différences n’étaient pas associées à la perception de la parole. En somme, ce mémoire apporte de nouvelles connaissances sur la nature des difficultés de perception de la parole, les effets du chant choral sur cette capacité ainsi que les mécanismes neurobiologiques sous-tendant ces effets.
iii
Abstract
Perceiving speech in noise becomes difficult in aging. The causes of these difficulties are mixed, but age-related decline of the dorsal speech stream may be a contributing factor. The practice of choral singing may help slow brain aging, but the effects of choral singing are not well known. The objectives of this master’s thesis were to investigate the relationship between the structural decline of the dorsal speech stream and difficulties in speech perception, and to identify how this relationship is modified by the practice of choral singing. 44 singers and 41 non-singers, young and old, completed a magnetic resonance imaging session, as well as cognitive and auditory assessments. A subgroup completed a syllable discrimination in noise task. In the first study, behavioural benefits were observed for older singers with certain characteristics related to their practice, suggesting that choral singing has the potential to benefit speech perception only under certain conditions. These benefits were particularly associated with the structure of regions of the dorsal speech stream. In a second study, the decline in the structure of several frontotemporal segments of the arcuate fasciculus, one of the main white matter fasciculi constituting the dorsal speech stream, was associated with the decline in speech perception. In addition, differences in structural asymmetry of the arcuate fasciculus were identified between groups, but these differences were not related to speech perception. In sum, these studies clarify the origin of speech perception difficulties in noise, the effects of choral singing on this ability, and the mechanisms underlying these effects.
iv
Table des matières
RÉSUMÉ ... II ABSTRACT ... III TABLE DES MATIÈRES ... IV LISTE DES FIGURES ... VIII LISTE DES TABLEAUX ... X LISTE DES ABRÉVIATIONS ... XI REMERCIEMENTS ... XIV AVANT-PROPOS ... XV
INTRODUCTION ... 1
CHAPITRE 1 IMAGERIE PAR RÉSONANCE MAGNÉTIQUE ET VIEILLISSEMENT CÉRÉBRAL ... 3
1.1IMAGERIE PAR RÉSONANCE MAGNÉTIQUE ... 3
1.1.1 Principe ... 3
1.1.2 Imagerie par résonance magnétique structurelle ... 4
1.1.2.1 Morphométrie basée sur le voxel ... 5
1.1.2.2 Morphométrie basée sur la surface ... 5
1.1.3 Imagerie par résonance magnétique de diffusion ... 6
1.1.3.1 Imagerie par tenseur de diffusion ... 7
1.1.3.2 Imagerie à haute résolution angulaire ... 9
1.2VIEILLISSEMENT CÉRÉBRAL ... 10
1.2.1 Imagerie par résonance magnétique structurelle ... 10
1.2.2 Imagerie par résonance magnétique de diffusion ... 11
CHAPITRE 2 PERCEPTION DE LA PAROLE ET VIEILLISSEMENT ... 14
2.1PERCEPTION DE LA PAROLE ... 14
2.1.1 Définition ... 14
2.1.2 Paradigmes expérimentaux de perception de la parole ... 15
2.1.3 Modèles à deux voies ... 17
2.2VIEILLISSEMENT DE LA PERCEPTION DE LA PAROLE ... 19
2.2.1 Facteurs affectant la perception de la parole dans le vieillissement ... 19
2.2.1.1 Présence de différents bruits ... 20
2.2.1.2 Intensité du bruit ... 21
2.2.2 Facteurs explicatifs des difficultés de perception de la parole liées à l’âge ... 21
2.2.2.1 Vieillissement de l’audition périphérique ... 21
2.2.2.2 Déficit du traitement central lié au vieillissement cérébral ... 23
2.2.2.2.1 Vieillissement de la cognition ... 23
2.2.2.2.2 Vieillissement de l’audition centrale ... 26
2.2.2.2.3 Vieillissement de la voie dorsale de la parole ... 27
CHAPITRE 3 EFFETS BÉNÉFIQUES DE LA PRATIQUE MUSICALE ... 29
v
3.2EFFETS BÉNÉFIQUES DE LA PRATIQUE MUSICALE SUR LA PERCEPTION DE LA PAROLE DANS LE
VIEILLISSEMENT ... 31 OBJECTIFS ET HYPOTHÈSES ... 34 ÉTUDE 1 ... 34 Objectifs ... 34 Hypothèses ... 34 ÉTUDE 2 ... 35 Objectifs ... 35 Hypothèses ... 35
CHAPITRE 4 ÉTUDE 1 : AGING OF SPEECH PERCEPTION IN NOISE IN SINGERS AND NON-SINGERS: A SURFACE-BASED MORPHOMETRY STUDY ... 36
4.1RÉSUMÉ ... 36
4.2ABSTRACT ... 37
4.3INTRODUCTION ... 38
4.4.MATERIAL AND METHODS ... 39
4.4.1 Participants ... 39
4.4.2 Musical activity questionnaire ... 43
4.4.3 Experimental design ... 44
4.4.4 Audiometric evaluation ... 44
4.4.5 SPiN task ... 44
4.4.6 Cognitive evaluation ... 46
4.4.7 MRI data acquisition ... 47
4.4.8 MRI data processing ... 47
4.4.9 Statistical analyses ... 49
4.5RESULTS ... 51
4.5.1 SPiN in aging singers and non-singers ... 51
4.5.2 SPiN in aging singers ... 52
4.5.3 SPiN in aging singers and non-singers based on singing parameters ... 54
4.5.4 The neurobiological correlates of singing-related parameters on SPiN ... 55
4.6DISCUSSION ... 59
4.6.1 Aging SPiN and musical practice ... 59
4.6.2 Neurobiological correlates of singers on SPiN ... 61
4.6.3 Thicker cortex ... 63 4.6.4 Limitations ... 63 4.5.5 Conclusion ... 64 4.7ACKNOWLEDGEMENTS ... 64 4.8AUTHORS CONTRIBUTIONS ... 65 4.9REFERENCES ... 65 4.10SUPPLEMENTARY MATERIALS ... 70
CHAPITRE 5 ÉTUDE 2 : THE FRONTO-TEMPORAL ORGANIZATION OF THE ARCUATE FASCICULUS AND ITS RELATIONSHIP WITH SPEECH PERCEPTION IN YOUNG AND OLDER AMATEUR SINGERS AND NON-SINGERS ... 71
5.1RÉSUMÉ ... 71
vi
5.3INTRODUCTION ... 73
5.4.MATERIAL AND METHODS ... 75
5.4.1 Participants ... 75
5.4.2 Experimental Design ... 78
5.4.3 Audiometric Evaluation ... 78
5.4.4 SPiN task ... 78
5.4.5 MRI Data Acquisition ... 79
5.4.6 MRI Data Processing ... 79
5.4.7 AF extraction ... 81
5.4.8 Statistical analyses ... 82
5.5RESULTS ... 83
5.5.1 AF Extraction ... 83
5.5.2 Age Effects and Group differences in the AF ... 84
5.5.2.1 Age effects ... 84
5.5.2.2 Group Differences ... 85
5.5.3 Relationship between AF and SPiN in Aging ... 87
5.5.4 Relationship between AF, Singing and SPiN ... 88
5.6DISCUSSION ... 89
5.6.1 Aging of the AF and SPiN ... 89
5.6.2 Amateur singing and the AF ... 91
5.6.3 Limits ... 93 5.6.4 Conclusion ... 93 5.7ACKNOWLEDGEMENTS ... 94 5.8AUTHORS CONTRIBUTION ... 94 5.9REFERENCES ... 95 5.10SUPPORTING INFORMATION ... 102
CHAPITRE 6 SYNTHÈSE DES RÉSULTATS, RETOMBÉES ET PERSPECTIVES ... 103
6.1SYNTHÈSE DES RÉSULTATS ... 103
6.1.1 Étiologie des difficultés de perception de la parole dans le vieillissement ... 103
6.1.2. Relation entre vieillissement, perception de la parole et chant ... 105
6.1.2.1 Effets comportementaux ... 105
6.1.2.2 Mécanismes neurobiologiques sous-tendant les effets comportementaux ... 106
6.2RETOMBÉES ET PERSPECTIVES DE L’ÉTUDE ... 109
6.2.1 Connaissances scientifiques et études futures ... 109
6.2.2 Perspectives cliniques ... 110
CONCLUSION ... 112
BIBLIOGRAPHIE ... 113
ANNEXE A RÉSUMÉ DES DIFFÉRENCES STRUCTURELLES ENTRE LES MUSICIENS ET LES NON-MUSICIENS IDENTIFIÉS DANS LA LITTÉRATURE À L’AIDE DE (A) L’IRM STRUCTURELLE ET (B) L’IRM DE DIFFUSION ... 130
ANNEXE B AGING OF SPEECH PERCEPTION IN NOISE IN SINGERS AND NON-SINGERS: A SURFACE-BASED MORPHOMETRY STUDY – SUPPLEMENTARY MATERIAL ... 139
vii
ANNEXE C THE FRONTO-TEMPORAL ORGANIZATION OF THE ARCUATE FASCICULUS AND ITS RELATIONSHIP WITH SPEECH PERCEPTION IN YOUNG AND OLDER AMATEUR SINGERS AND NON-SINGERS – SUPPORTING INFORMATION ... 147
viii
Liste des figures
FIGURE 1.SÉQUENCE D’IMAGERIE PAR RÉSONANCE MAGNÉTIQUE PONDÉRÉE EN T1 D’UN CERVEAU HUMAIN DE JEUNE ADULTE.LA MATIÈRE BLANCHE APPARAÎT HYPERINTENSE (GRIS CLAIR), LE LIQUIDE CÉRÉBRO-SPINAL APPARAÎT HYPOINTENSE (NOIR) ET LA MATIÈRE GRISE SE SITUE ENTRE LES DEUX (GRIS FONCÉ). ... 5
FIGURE 2.REPRÉSENTATION D’UN TENSEUR DE DIFFUSION ET DU CALCUL DES DIFFÉRENTES MÉTRIQUES DE DIFFUSION.(A)
REPRÉSENTATION MATHÉMATIQUE DU TENSEUR DE DIFFUSION CORRESPONDANT À UNE MATRICE SYMÉTRIQUE 3 X 3 OBTENUE PAR L’ACQUISITION D’AU MOINS 6 DIRECTIONS DE DIFFUSION.(B)VALEURS PROPRES DU TENSEUR DE DIFFUSION
(Λ1, Λ2, Λ3) REPRÉSENTANT LA DIFFUSION DES MOLÉCULES D’EAU DANS TROIS DIRECTIONS LINÉAIREMENT INDÉPENDANTES DE L’ESPACE.(C)CALCUL DES MÉTRIQUES DE DIFFUSION À PARTIR DES VALEURS PROPRES DU TENSEUR DE DIFFUSION. ... 8
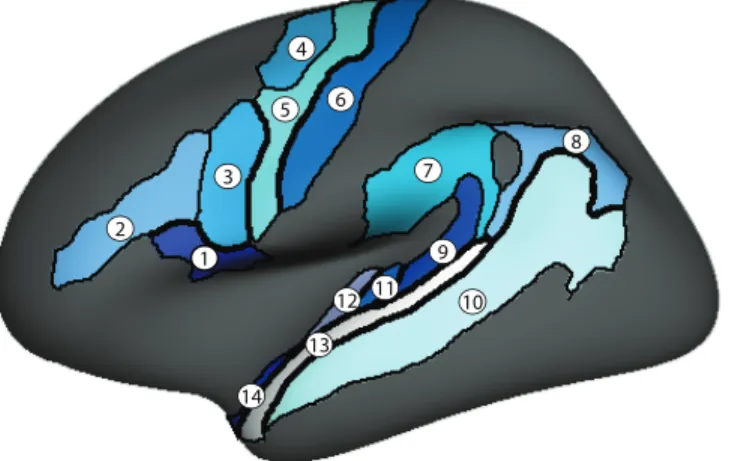
FIGURE 3.LOCATION OF THE REGIONS OF INTEREST (ROIS) REPRESENTED ON THE GROUP AVERAGE INFLATED SURFACE (ONLY THE LEFT HEMISPHERE IS SHOW).1) THE PARS OPERCULARIS OF THE INFERIOR FRONTAL GYRUS (PIFG),2) THE INFERIOR FRONTAL SULCUS (IFS),3) THE DORSAL PRECENTRAL SULCUS (DPRS),4) THE VENTRAL PRECENTRAL SULCUS (VPRS),5) THE
PRECENTRAL GYRUS (PRG),6) THE CENTRAL SULCUS (CS),7) THE SUPRAMARGINAL GYRUS (SMG),8) THE ANGULAR GYRUS
(ANG),9) THE PLANUM TEMPORALE (PT),10) THE SUPERIOR TEMPORAL SULCUS (STS),11) THE TRANSVERSE TEMPORAL GYRUS (TTG),12) THE TRANSVERSE TEMPORAL SULCUS (TTS),13) THE LATERAL ASPECT OF THE SUPERIOR TEMPORAL GYRUS (LSTG) AND 14) THE PLANUM POLARE (PP). ... 48
FIGURE 4.STATISTICAL MODELS FOR THE MODERATION AND MEDIATION ANALYSES.(A)CONCEPTUAL DIAGRAM AND (B)
STATISTICAL DIAGRAM FOR THE MODERATION ANALYSIS.(C)CONCEPTUAL DIAGRAM AND (D) STATISTICAL DIAGRAM OF THE MODERATED MEDIATION ANALYSIS. ... 49
FIGURE 5.AGE EFFECTS ON SPIN.THE BAR CHART SHOWS SPIN MEASURES (SENSITIVITY (D’), ACCURACY OR REACTION TIME (T)) ON THE Y-AXIS AND AGE ON THE X-AXIS.(A)RESULTS OF THE MODERATION ANALYSES CONDUCTED ON ALL PARTICIPANTS.(B)
RESULTS OF THE LINEAR REGRESSION ANALYSES CONDUCTED ONLY ON SINGERS.(C)RESULTS OF THE MODERATION ANALYSES CONDUCTED ON ALL PARTICIPANTS EXCLUDING SINGERS WITH INFREQUENT PRACTICE AT HOME.DOTTED LINE REPRESENTED
95% CONFIDENCE INTERVALS OF THE MEAN OF THE REGRESSION LINE. ... 52
FIGURE 6.RELATIONSHIP BETWEEN AGING, SINGING PARAMETERS AND SPIN.THE BAR CHART SHOWS SPIN MEASURES
(SENSITIVITY (D’) OR ACCURACY) ON THE Y-AXIS AND AGE ON THE X-AXIS.(A)MAIN EFFECT OF PRACTICE FREQUENCY ON
SPIN.(B)MODERATING EFFECT OF THE NUMBER OF SINGING LANGUAGES ON THE RELATIONSHIP BETWEEN AGE AND SPIN.
(C)MODERATING EFFECT OF FORMAL SINGING TRAINING ON THE RELATIONSHIP BETWEEN SPIN AND AGE. ... 54
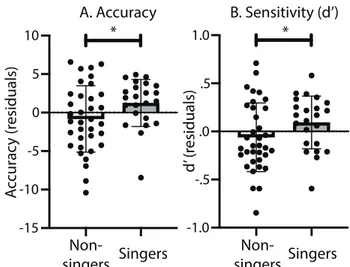
FIGURE 7.THE BOX PLOTS ILLUSTRATE THE EFFECT OF SINGING ON (A) ACCURACY AND (B) SENSITIVITY (D’), SEPARATELY FOR ALL NON-SINGERS AND FOR SINGERS WITH FREQUENT PRACTICE AT HOME (>1/WEEK).EACH BLACK DOT REPRESENTS A PARTICIPANT.ASTERISKS INDICATE SIGNIFICANCE AT P<0.05.ERROR BARS REPRESENT REPRESENTED THE 95% CONFIDENCE INTERVALS OF THE MEAN. ... 55
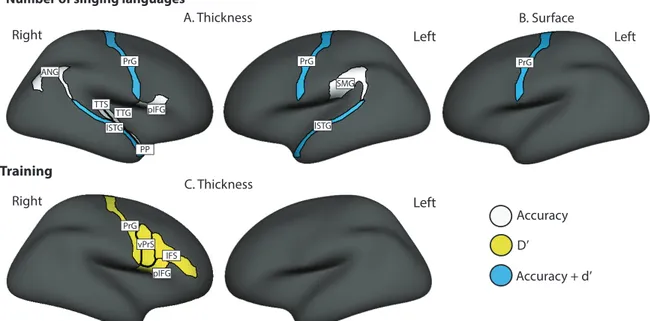
FIGURE 8.ILLUSTRATION OF THE ROIS SHOWING A SIGNIFICANT MODERATED MEDIATION, PRESENTED ON THE GROUP AVERAGE INFLATED BRAIN SURFACE.(A)ROIS SHOWING AN INDIRECT EFFECT OF AGE ON SPIN THROUGH CORTICAL THICKNESS
(POSITIVE MODERATION BY THE NUMBER OF SINGING LANGUAGES).(B)ROIS SHOWING AN INDIRECT EFFECT OF AGE ON
SPIN THROUGH CORTICAL SURFACE (POSITIVE MODERATION BY THE NUMBER OF SINGING LANGUAGES).(C)ROIS SHOWING AN INDIRECT EFFECT OF AGE ON SPIN THROUGH CORTICAL THICKNESS (POSITIVE MODERATION BY FORMAL SINGING TRAINING).IN THE FIGURE, WHITE COLOUR IDENTIFIES ROIS SHOWING A MODERATED INDIRECT EFFECT OF AGE ON ACCURACY THROUGH ROI STRUCTURE, YELLOW IDENTIFIES MODERATED INDIRECT EFFECTS OF AGE ON SENSITIVITY (D’) AND BLUE IDENTIFIES REGIONS IN WHICH MODERATED INDIRECT EFFECTS OF AGE WERE FOUND ON BOTH ACCURACY AND D’. .... 57
FIGURE 9.EXAMPLE OF A LEFT ARCUATE FASCICULUS (PARTICIPANT ID:S029) DIVIDED INTO TEN SEGMENTS FROM THE TEMPORAL LOBE (1) TO THE FRONTAL LOBE (10).THE FASCICULUS IS DISPLAYED ON THE MNI152C 2009 NONLINEAR SYMMETRICAL TEMPLATE (FONOV,EVANS,MCKINSTRY,ALMLI,&COLLINS,2009), WHICH WAS WARPED TO THE INDIVIDUAL ANATOMICAL SCAN (T1) OF THE PARTICIPANT. ... 82
ix
FIGURE 10.AVERAGE OF THE RIGHT AND LEFT AF OF ALL PARTICIPANTS.THE AVERAGE AF ARE DISPLAYED ON THE MNI152C
2009 NONLINEAR SYMMETRICAL TEMPLATE (FONOV ET AL.,2009) IN STANDARD SPACE. ... 84
FIGURE 11.AGE-RELATED AND HEMISPHERIC EFFECTS FOR THE TEN SEGMENTS OF THE BILATERAL AF.(A)AGE EFFECTS.AN UPWARD-POINTINGARROW INDICATES THAT AGE IS ASSOCIATED WITH AN INCREASED VALUE WHILE A DOWNWARD -POINTINGARROW INDICATES THAT AGE IS ASSOCIATED WITH A DECREASED VALUE.A GREEN CIRCLE SIGNALS A BENEFICIAL AGE EFFECT WHILE A RED CIRCLE SIGNALS A DETRIMENTAL AGE EFFECT.(B)HEMISPHERIC DIFFERENCES.THE LETTER IN THE CIRCLES INDICATES THE HEMISPHERE (L: LEFT;R: RIGHT) WITH THE HIGHEST VALUE.A GREEN CIRCLE SIGNALS A BENEFICIAL EFFECT ON THE WHITE MATTER WHILE A RED CIRCLE SIGNALS A DETRIMENTAL EFFECT.(C)RESULTS OF THE LINEAR
REGRESSION ANALYSES CONDUCTED TO DECOMPOSE THE AGE X HEMISPHERE INTERACTIONS.AN UPWARD-POINTINGARROW INDICATES THAT AGE IS ASSOCIATED WITH AN INCREASED VALUE WHILE A DOWNWARD-POINTINGARROW INDICATES THAT AGE IS ASSOCIATED WITH A DECREASED VALUE.A DOUBLE ARROW INDICATES THE HEMISPHERE WITH THE STRONGEST AGE EFFECT.A GREEN CIRCLE SIGNALS A BENEFICIAL AGE EFFECT ON THE WHITE MATTER WHILE A RED CIRCLE SIGNALS A
DETRIMENTAL AGE EFFECT. ... 85
FIGURE 12.SUMMARY OF THE INTERHEMISPHERIC ASYMMETRY DIFFERENCES FOUND BETWEEN GROUPS.(A)AGE-INDEPENDENT DIFFERENCES ARE REPRESENTED IN WHITE CIRCLES.THE LETTER IN THE CIRCLES INDICATES THE DIRECTION OF THE ASYMMETRY FOR SINGERS (L: LEFTWARD;R: RIGHTWARD).IN 50% OF THESE CASES, SINGERS PRESENTED GREATER WHITE MATTER STRUCTURE IN THE LEFT HEMISPHERE COMPARED TO THE RIGHT (FA, SEGMENT 3;RD, SEGMENT 1;AD, SEGMENT 4; VOLUME, SEGMENT 6).IN THE OTHER CASES, SINGERS PRESENTED GREATER WHITE MATTER STRUCTURE IN THE RIGHT HEMISPHERE COMPARED TO THE LEFT (FA, SEGMENTS 5 AND 6;RD, SEGMENT 5,MD, SEGMENT 4).(B).AGE-DEPENDENT ASYMMETRY DIFFERENCES.THE LETTER IN THE CIRCLES INDICATES THE DIRECTION OF THE AGE-RELATED INCREASING ASYMMETRY FOR SINGERS.A GREEN CIRCLE SIGNALS A BENEFICIAL AGE EFFECT FOR SINGERS AND A RED CIRCLE SIGNALS A DETRIMENTAL AGE EFFECT FOR SINGERS (SEE RESULTS AND DISCUSSION SECTIONS FOR EXPLANATION). ... 87
FIGURE 13.RESULTS OF THE MEDIATION ANALYSES.THE STATISTICAL DIAGRAMS ILLUSTRATE EACH OF THE INDIRECT EFFECTS (AB) OF AGE ON SENSITIVITY THROUGH WHITE MATTER OF THE WHOLE AF(A) AND THROUGH SPECIFIC SEGMENTS (B-E).FOR EACH PATH IN THE GRAPHS, THE UNSTANDARDIZED COEFFICIENTS AND THE PROBABILITY VALUE ARE REPORTED.THE BOOTSTRAPPED 95% CONFIDENCE INTERVAL IS PROVIDED FOR THE INDIRECT EFFECTS (AB). ... 88
x
Liste des tableaux
TABLE 1.PARTICIPANTS CHARACTERISTICS SEPARATELY FOR EACH GROUP.INDEPENDENT T-TESTS WERE CONDUCTED TO COMPARE THE GROUPS ON EACH VARIABLE. ... 41
TABLE 2.PARTICIPANTS’ SINGING EXPERIENCE. ... 43
TABLE 3.RESULTS OF THE SIGNIFICANT MODERATED MEDIATIONS FOR A) ACCURACY AND B) D’ FOR THE NUMBER OF SINGING LANGUAGES. ... 58 TABLE 4.RESULTS OF THE SIGNIFICANT MODERATED MEDIATIONS FOR D’ FOR THE FORMAL SINGING TRAINING.
... 59 TABLE 5.GROUP CHARACTERISTICS.INDEPENDENT T-TESTS WERE CONDUCTED TO COMPARE GROUPS. ... 77
TABLEAU 6.RÉSUMÉ DES DIFFÉRENCES STRUCTURELLES ENTRE LES MUSICIENS ET LES NON-MUSICIENS IDENTIFIÉS DANS LA LITTÉRATURE À L’AIDE DE (A) L’IRM STRUCTURELLE ET (B) L’IRM DE DIFFUSION. ... 130
xi
Liste des abréviations
ABRÉVIATION SIGNIFICATION FRANÇAISE SIGNIFICATION ANGLAISE
AD Diffusivité axiale Axial diffusivity
AF Faisceau arqué Arcuate fasciculus
AFD Densité apparente de fibres Apparent fiber density
ANG Gyrus angulaire Angular gyrus
ANTs - Advanced Normalization Tools
bet - Brain Extraction Tool
C Biais de réponse Response biais
CS Sulcus central Central sulcus
d’ Sensibilité Sensitivity
dB Décibel Decibel
DIVA - Directions Into Velocities of
Articulators Model dPrS Sulcus précentral dorsal Dorsal precentral sulcus
DTI Imagerie par tenseur de diffusion Diffusion tensor imaging DW Pondération en diffusion Diffusion weighted
FA Anisotropie fractionnelle Fractionnal anisotropy FDR Taux de fausses découvertes False discovery rate
fODF Fonction d’orientation de distribution de fibres Fiber orientation distribution function
FSL - FMRIB Software Library
GAI Inventaire d'anxiété gériatrique Geriatric Anxiety Inventory GDS Échelle de dépression gériatrique Geriatric Depression Scale HARDI Imagerie de diffusion à haute
résolution angulaire
High angular resolution diffusion imaging
HCP - Human Connectome Project
HG Gyrus de Heschl Heschl gyrus
xii
IFS Sulcus frontal inférieur Inferior frontal sulcus
LMM Modèle linéaire mixte Linear mixed model
lSTG Gyrus temporal supérieur latéral Lateral aspect of the superior temporal gyrus
MD Diffusivité moyenne Mean diffusivity
MoCA - Montreal Cognitive Assessment
MRI (IRM) Imagerie par résonance magnétique Magnetic resonance imaging NuFO Nombre d'orientations de fibre Number of fiber orientations OPERA - Overlap, Precision, Emotion, Repetition, Attention Model
pIFG Gyrus frontal inférieur - pars opercularis Inferior frontal gyrus - pars opercularis
PMv Prémoteur ventral Ventral premotor cortex
PP Planum polare Planum polare
PrG Gyrus précentral Precentral gyrus
pSTG Gyrus supérieur temporal postérieur Posterior superior temporal gyrus
PT Planum temporale Planum temporale
PTA Moyenne des sons purs Pure tone average
RD Diffusivité radiale Radial diffusivity
RMS Valeur moyenne quadratique Root mean square
ROI Région d'intérêt Region of interest
RT Temps de réaction Reaction time
SBM Morphométrie basée sur la surface Surface-based morphometry
SCILPY - Sherbrooke Connectivity Imaging Lab PYthon
SMG Gyrus supramarginal Supramarginal gyrus
SNR Ratio signal sur bruit Signal to noise ratio SPiN (PPb) Perception de la parole dans le bruit Speech perception in noise
STG Gyrus supérieur temporal Superior temporal gyrus STS Sulcus temporal superieur Superior temporal sulcus
xiii
TAiL - Test of Attention in Listening
TMS Stimulation magnétique
transcrânienne Transcranial magnetic stimulation TTG Gyrus temporal transverse Transverse temporal gyrus
TTS Sulcus temporal transverse Transverse temporal sulcus VBM Morphométrie basée sur le voxel Voxel-based morphometry
xiv
Remerciements
Je tiens à remercier ma directrice de maîtrise, Pascale Tremblay, professeure au département de réadaptation de l’Université Laval, pour m’avoir transmis sa passion pour les neurosciences et la neuroimagerie. Je la remercie également de m’avoir conseillé, orienté et aidé tout le long de ma maîtrise, mais également depuis le début de mon baccalauréat. Toutes ses années passées dans son laboratoire en tant que stagiaire et étudiant aux cycles supérieurs furent riches en apprentissages.
Je remercie également les collaborateurs qui ont participé à mon projet de maîtrise. Merci à Josée Vaillancourt, professeure à la Faculté de musique de l’Université Laval, pour son aide et ses conseils et à Maxime Descoteaux, professeur au département d’informatique de l’Université de Sherbrooke, pour son support et son aide dans le traitement des données de diffusion.
Je tiens également à remercier tous mes collègues de laboratoire qui m’ont soutenu et qui ont enrichi mon expérience de laboratoire. Un merci particulier à Valérie Brisson, Élisabeth Maillard, Émilie Belley, Julie Poulin, Catherine Denis, Lisa-Marie Deschênes et Chloé Gingras avec qui j’ai passé d’excellents moments et qui, grâce à leur joie de vivre et leur support, m’ont aidé à rester motivé.
Un énorme merci à ma mère qui m’a toujours encouragé dans mes projets et cru en moi.
Je remercie également mes amis en dehors du laboratoire, particulièrement Maxime Lavoie, Audrée Laroche, Justine Pellerin, Camille Gélinas, Christine Vézina et Joëlle Landry, qui m’ont toujours écouté et supporté.
xv
Avant-propos
Ce mémoire est issu de ma maîtrise en sciences cliniques et biomédicales que j’ai réalisée sous la supervision de Pascale Tremblay au centre de recherche CERVO à Québec. Les chapitres 4 et 5 présentent les résultats des travaux que j’ai effectués pendant ma maîtrise et soumis (ou que j’aurai prochainement soumis) en tant que premier auteur :
Le chapitre 4 contient l’article « Aging of speech perception in noise in singers and non-singers: a surface-based morphometry study ». Cette publication a été réalisée lors de mon projet de maîtrise. Elle n’a pas encore été soumise puisque des analyses supplémentaires sont en cours. L’article a été rédigé par moi-même. J’ai également contribué à toutes les étapes de l’élaboration du projet, au recrutement des participants, à la passation des tests et à l’analyse des données comportementales et de neuroimagerie. Les coauteurs sont Josée Vaillancourt et Pascale Tremblay. Josée Vaillancourt a apporté ses conseils sur l’aspect du chant choral et a participé à la rédaction de l’article. Pascale Tremblay a supervisé chaque étape de l’élaboration du projet, de la collecte de données et de l’analyse des données, ainsi que participé à la rédaction de l’article en bonifiant chaque section et en proposant des modifications au besoin.
Le chapitre 5 contient l’article « The fronto-temporal organization of the arcuate fasciculus and its relationship with speech perception in young and older amateur singers and non-singers » qui est issu du même projet que l’article du Chapitre 4. Cette publication a été réalisée lors de mon projet de maîtrise. Elle a été soumise au journal Humain Brain Mapping le 4 août 2020. Elle a été rédigée par moi-même. J’ai également contribué à toutes les étapes de l’élaboration du projet, au recrutement des participants, à la passation des tests et à l’analyse des données comportementales et de neuroimagerie. Les coauteurs sont Guillaume Theaud, Maxime Descoteaux et Pascale Tremblay. Guillaume Theaud et Maxime Descoteaux ont collaboré au traitement, à l’analyse des données de neuroimagerie et à la rédaction de l’article. Maxime Descoteaux a également supervisé l’analyse des données de neuroimagerie. Pascale Tremblay a supervisé chaque étape de l’élaboration du projet, de la collecte de données et de l’analyse des données, ainsi que participé à la rédaction de l’article en bonifiant chaque section et en proposant des modifications au besoin.
1
Introduction
L’accroissement continu de l’espérance de vie au cours du dernier siècle est attribuable en grande partie aux grandes avancées de la science moderne. Statistique Canada estime que le vieillissement de la population du Canada devrait s’accélérer dans les prochaines années. Ainsi, la proportion de personnes âgées de 65 ans et plus devrait être environ 4,2 % à 12,3 % supérieure en 2068 comparativement à 2018 (Statistique Canada, 2019). Plus spécifiquement au Québec, Statistique Canada évalue que la proportion de la population active, c’est-à-dire les personnes âgées entre 15 à 64 ans, diminuera d’environ 5 % d’ici les 10 à 15 prochaines années. Dans un contexte de vieillissement de la population, il devient donc de plus en plus important d’investiguer les changements qui surviennent au cours du vieillissement normal, de comprendre l’origine de ces changements et d’étudier des stratégies pouvant les réduire, les renverser ou les prévenir afin de promouvoir un vieillissement sain pour l’ensemble de la population.
Lors du vieillissement normal, des changements surviennent dans la structure du cerveau. Ces changements se caractérisent par une dégradation de la structure de la matière grise et de la matière blanche et une augmentation de la quantité de liquide cérébro-spinal. Ce déclin structurel a souvent été associé et corrélé aux déclins d’habiletés cognitives et langagières chez les aînés, tels que le déclin de la capacité à percevoir la parole dans le bruit (PPb) (Bilodeau-Mercure, Lortie, Sato, Guitton, & Tremblay, 2015; Du, Buchsbaum, Grady, & Alain, 2016; Eckert et al., 2008; Erb & Obleser, 2013; Harris, Dubno, Keren, Ahlstrom, & Eckert, 2009; Hwang, Li, Wu, Chen, & Liu, 2007; Manan, Yusoff, Franz, & Mukari, 2017; Peelle, Troiani, Grossman, & Wingfield, 2011; Rudner, Seeto, Keidser, Johnson, & Rönnberg, 2019; Sheppard, Wang, & Wong, 2011; Tremblay et al., 2019; Vaden, Kuchinsky, Ahlstrom, Dubno, & Eckert, 2015; Vaden et al., 2016; Vaden et al., 2011; Wong, Ettlinger, Sheppard, Gunasekera, & Dhar, 2010; Wong et al., 2009).
Les difficultés de PPb sont l’une des plaintes les plus fréquentes des personnes âgées. Puisque les environnements bruyants sont communs (p. ex. restaurants, rues, centres commerciaux, etc.), ces difficultés peuvent entraîner des problèmes de communication ayant un impact négatif sur la vie quotidienne et la qualité de vie. Malheureusement, les causes de ce déclin sont encore mal comprises, ce qui rend l’identification de stratégies d’intervention difficile. Plusieurs hypothèses ont été proposées pour expliquer la diminution liée à l’âge de la PPb, mais il n’existe pas de consensus sur les mécanismes sous-jacents. Dans ce mémoire, la contribution du déclin du traitement auditif périphérique et central, du traitement cognitif et du traitement de la parole sera discutée.
2
De récentes études scientifiques suggèrent que pratiquer une activité musicale, soit jouer un instrument de musique ou chanter dans une chorale, pourrait être une stratégie efficace pour atténuer les difficultés de PPb au cours du vieillissement normal (Alain, Zendel, Hutka, & Bidelman, 2014; Anderson, White-Schwoch, Parbery-Clark, & Kraus, 2013; Bidelman & Alain, 2015; Dubinsky, Wood, Nespoli, & Russo, 2019; Fleming, Belleville, Peretz, West, & Zendel, 2019; Fostick, 2019; Parbery-Clark, Strait, Anderson, Hittner, & Kraus, 2011; White-Schwoch, Woodruff Carr, Anderson, Strait, & Kraus, 2013; Zendel & Alain, 2012; Zendel, West, Belleville, & Peretz, 2019). Ces résultats sont conformes à la notion selon laquelle le déclin cognitif peut être atténué par la pratique d’activités physiques, intellectuelles et sociales (Scarmeas & Stern, 2003; Vaden et al., 2015), un phénomène que l’on appelle la plasticité dépendante de l’expérience.
La plasticité dépendante de l’expérience est un phénomène de réorganisation neuronale résultant de l’expérience de vie d’une personne. Les activités musicales pourraient donc prévenir le déclin de la PPb à travers une réorganisation et/ou un maintien de la structure du cerveau avec l’âge. Toutefois, à ce jour, aucune étude n’a étudié les changements dans la structure du cerveau liés à la pratique d’activités musicales et bénéfiques pour la PPb lors du vieillissement. L’imagerie par résonance magnétique (IRM) est une technique qui a le potentiel d’améliorer les connaissances sur les mécanismes de vieillissement cérébral, mais également de fournir des renseignements sur les mécanismes de plasticité structurelle associés aux avantages d’une pratique musicale sur la PPb.
L’objectif principal de ce mémoire est de mieux comprendre la nature des difficultés de PPb avec l’âge en explorant les effets du vieillissement de la structure de la voie dorsale de la parole, impliquée dans le traitement de la parole, et d’examiner comment les effets du vieillissement cérébral affectent la PPb chez des non-chanteurs et des chanteurs en utilisant l’IRM structurelle et de diffusion.
Dans ce mémoire, un survol des techniques d’IRM permettant d’étudier la structure de la matière grise et de la matière blanche sera d’abord présenté. Ce survol sera suivi d’une courte revue de l’état des connaissances sur le vieillissement du cerveau. Ensuite, les mécanismes par lesquels nous percevons la parole dans le bruit ainsi que l’impact du vieillissement de ces mécanismes sur la PPb seront détaillés. Finalement, les derniers chapitres lieront vieillissement cérébral, pratique musicale et PPb à l’aide de revues de la littérature et des deux études incluses dans ce mémoire.
3
Chapitre 1 Imagerie par résonance magnétique et
vieillissement cérébral
1.1 Imagerie par résonance magnétique
L’IRM est une technique non invasive et non douloureuse qui permet d’étudier la structure des différents organes et tissus du corps, incluant le cerveau. Le cerveau est divisé en trois catégories de tissus distincts : la matière grise – ou cortex cérébral –, la matière blanche et le liquide cérébro-spinal. La matière grise comprend les corps cellulaires des neurones impliqués dans la génération et le traitement d’information sous forme d’influx nerveux. La matière blanche comprend les prolongements des corps cellulaires impliqués dans la transmission de l’information dans tout le système nerveux. Le liquide cérébro-spinal correspond au liquide dans lequel le système nerveux se situe. Ces trois tissus sont essentiels au maintien des fonctions cérébrales et sont susceptibles d’être affectés par le vieillissement normal. Ce chapitre abordera le fonctionnement de l’IRM et les différentes modalités d’imagerie utilisées dans ce mémoire pour étudier la structure du cerveau, et proposera un bref aperçu du vieillissement cérébral identifié dans la littérature à l’aide de ces techniques.
1.1.1 Principe
L’IRM se base sur le phénomène de résonance magnétique nucléaire. Ce phénomène se produit lorsque les tissus sont placés dans un fort champ magnétique en présence d’impulsions de radiofréquence. L’IRM tire profit de la forte prévalence de l’atome d’hydrogène dans le corps et des propriétés magnétiques de son proton (+). En effet, le corps est majoritairement composé de molécules d’eau possédant deux atomes d’hydrogène chacun. Le proton de l’hydrogène induit un faible champ magnétique en raison de sa rotation sur lui-même. En absence d’un champ magnétique, les axes de rotation des protons sont distribués aléatoirement dans l’espace. À l’inverse, lorsque les protons sont en présence d’un champ magnétique externe, leur axe de rotation s’aligne parallèlement ou antiparallèlement au champ magnétique.
4
Le phénomène de résonance magnétique nucléaire a lieu lorsqu’une impulsion de radiofréquence de courte durée est appliquée dans ce champ magnétique externe. L’onde de radiofréquence procure de l’énergie aux protons d’hydrogène, ce qui change l’orientation de leur axe de rotation de telle sorte qu’ils ne sont plus alignés avec le champ magnétique. Ensuite, l’onde de radiofréquence est retirée. Les protons d’hydrogène libèrent l’énergie acquise et se réalignent avec le champ magnétique toujours présent. Cette libération d’énergie est captée par une antenne réceptrice et permettra la formation d’images à la suite d’un traitement ultérieur. Le temps de réalignement de l’axe de rotation des protons d’hydrogène dépend du type de tissu dans lequel ils se trouvent, ce qui permet de distinguer les tissus entre eux. En faisant varier plusieurs paramètres d’acquisition (p. ex. l’intervalle entre deux impulsions de radiofréquence), il est possible d’obtenir diverses modalités d’imagerie.
Parmi les différentes modalités d’imagerie utilisées pour étudier la structure des tissus, on retrouve l’IRM structurelle et l’IRM de diffusion. Ces deux techniques offrent des avenues intéressantes pour étudier les changements dans la structure du cerveau au cours de la vie. Ces deux modalités d’imagerie seront décrites dans les prochaines sections de ce chapitre.
1.1.2 Imagerie par résonance magnétique structurelle
L’IRM structurelle réfère à l’acquisition d’images de la structure du cerveau — ou images anatomiques. L’IRM structurelle emploie généralement une séquence d’acquisition dite « pondérée en T1 » puisque cette dernière offre une bonne distinction entre la matière grise, la matière blanche et le liquide cérébro-spinal. Dans une séquence pondérée en T1 (Figure 1), la matière blanche apparaît hyperintense (gris clair), le liquide cérébro-spinal apparaît hypointense (noir) et la matière grise se situe entre les deux (gris foncé). L’IRM structurelle combinée à des techniques dites de « morphométrie » (voir section 1.1.2.1 et 1.1.2.2) permet d’extraire des informations caractérisant la structure des tissus. Plusieurs approches semi-automatisées de morphométrie existent et nécessitent plusieurs étapes de traitement. Dans ce mémoire, la morphométrie basée sur le voxel (« voxel-based morphometry », VBM) et la morphométrie basée sur la surface (« surface-based morphometry », SBM) seront présentées dans les deux prochaines sections.
5
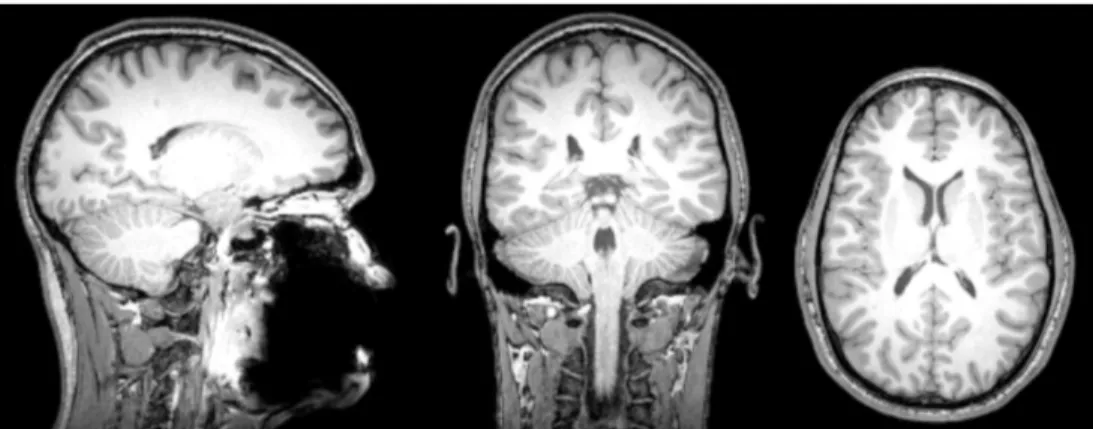
Figure 1. Séquence d’imagerie par résonance magnétique pondérée en T1 d’un cerveau humain de jeune adulte. La matière blanche apparaît hyperintense (gris clair), le liquide cérébro-spinal apparaît hypointense (noir) et la matière grise se situe entre les deux (gris foncé).
1.1.2.1 Morphométrie basée sur le voxel
La VBM est l’une des techniques les plus utilisées pour extraire des informations sur la structure des différentes régions du cerveau. La VBM se base sur une représentation volumétrique du cerveau, c’est-à-dire qu’en utilisant cette méthode il est possible d’obtenir le volume de différentes régions sur la base du calcul de l’intensité et du nombre de voxels (pixels volumétriques) (Ashburner & Friston, 2000; Whitwell, 2009; Wright et al., 1995). Il est également possible de calculer la densité de la matière grise. Cette mesure est dérivée de la segmentation de l’image et est liée à l’intensité du signal T1. La densité de la matière grise est une mesure qui décrit le nombre de voxels qui sont segmentés comme étant de la matière grise par rapport au nombre de voxels qui sont segmentés comme étant de la matière blanche, ou un autre type de tissu cérébral. Bien que cette technique offre plusieurs applications intéressantes pour caractériser et comparer la structure cérébrale entre diverses populations, elle possède plusieurs limites importantes (Mechelli, Price, Friston, & Ashburner, 2005), dont le fait que des changements de volume peuvent être sous-tendus par des changements d’autres mesures morphologiques telles que la surface, l’épaisseur et la gyrification. Ces trois mesures peuvent être extraites par des techniques de SBM.
6
Le développement de la SBM a permis l’obtention de mesures morphologiques corticales plus précises que le volume, et ce, à une meilleure résolution que le voxel (sous-voxel) (Dale, Fischl, & Sereno, 1999 ; Fischl, Sereno, & Dale, 1999). Ces mesures sont obtenues par des méthodes d’extraction dans lesquelles des modèles déformables de la surface corticale sont utilisés pour créer des surfaces internes et externes du cortex (Dale, Fischl, & Sereno, 1999 ; Fischl, Sereno, & Dale, 1999). Ces mesures comprennent l’épaisseur, la surface et la gyrification corticale. La surface et l’épaisseur sont deux sous-mesures du volume (surface x épaisseur = volume) alors que la gyrification est un index du repliement du cortex. Selon l’hypothèse de l’unité radiale (Rakic, 1988), la surface et l’épaisseur représentent des caractéristiques morphologiques distinctes des propriétés structurelles de la matière grise. La surface corticale serait déterminée par le nombre de colonnes verticales de neurones dans le cortex alors que l’épaisseur corticale serait déterminée par le nombre et la taille des cellules dans une colonne. Il a été proposé que ces deux mesures sont génétiquement indépendantes (Panizzon et al., 2009) et que leur développement est modulé par des mécanismes cellulaires distincts (Pontious, Kowalczyk, Englund, & Hevner, 2008). En effet, des études chez les modèles animaux ont montré que le développement de l’épaisseur corticale peut être modifié, et ce sans affecter le développement de la surface corticale, par des mutations génétiques affectant les cellules progénitrices intermédiaires1 impliquées dans la neurogenèse (Pontious et al., 2008). Ainsi, en
étudiant et comparant ces trois mesures (surface, épaisseur, gyrification) avec celle du volume, il est possible d’obtenir des informations plus détaillées sur les mécanismes sous-tendant la structure cérébrale.
1.1.3 Imagerie par résonance magnétique de diffusion
L’IRM de diffusion réfère quant à elle à l’acquisition d’images du déplacement des molécules d’eau dans les tissus. Cette séquence d’acquisition se base sur le principe selon lequel les déplacements des molécules sont contraints par l’organisation macro- et microstructurelle de leur environnement. En effet, la direction de diffusion des molécules d’eau est restreinte par des obstacles moléculaires tels que la présence de macromolécules, de fibres et/ou de membranes (Le Bihan, Turner, & Douek, 1993). La vitesse de diffusion diminue lorsque les molécules d’eau rencontrent un obstacle perpendiculaire à leur trajectoire. Ainsi, la structure des fibres de la matière blanche confère
1 Cellules capables de se différencier, mais dont le potentiel de différenciation est limité à certains types de cellules du système nerveux. Elles sont présentes dans le cortex cérébral en développement et ont la capacité de produire des neurones par l’intermédiaire d’un processus appelé neurogenèse.
7
à l’eau une diffusion anisotropique, c’est-à-dire une diffusion non équivalente dans toutes les directions puisque les molécules sont restreintes dans leurs déplacements par la présence des membranes et de la myéline des axones. Dans d’autres régions du cerveau, telles que les ventricules, la diffusion de l’eau est dite isotropique, soit une diffusion égale dans toutes les directions puisque l’eau ne se situe pas dans une région restrictive en termes d’obstacles. Ainsi, en modifiant les paramètres des séquences d’acquisition des images, notamment en laissant les molécules d’eau diffuser pendant un court laps de temps, il est possible de mesurer la distribution des directions de diffusion dans chaque voxel de l’image (Le Bihan, 2003). En rassemblant l’information de chaque voxel, l’IRM de diffusion reconstruit les faisceaux de fibres de matière blanche à l’aide de techniques de suivi 3D (Mori & van Zijl, 2002). Le processus par lequel les faisceaux de matière blanche sont reconstruits se nomme tractographie alors que le processus par lequel les métriques de diffusion sont extraites se nomme tractométrie. Dans ce mémoire, deux techniques d’IRM de diffusion seront présentées : l’imagerie par tenseur de diffusion (« diffusion tensor imaging », DTI) et l’IRM de diffusion à haute résolution angulaire (« high angular resolution diffusion imaging », HARDI).
1.1.3.1 Imagerie par tenseur de diffusion
À partir des patrons de diffusion de l’eau, il est possible de calculer un tenseur de diffusion (Basser & Jones, 2002; Basser, Mattiello, & LeBihan, 1994). Mathématiquement, un tenseur de diffusion est une matrice symétrique 3 x 3 obtenue par l’acquisition d’au moins six mesures de diffusion dans différentes directions (Figure 2A). L’acquisition d’un minimum de six mesures de diffusion est suffisante puisque le tenseur est symétrique. Par une décomposition en éléments propres (« eigendecomposition »), le tenseur de diffusion décrit les propriétés de diffusion de l’eau dans trois axes par trois valeurs propres (λ1, λ2, λ3) (Figure 2B). Ces trois valeurs permettent le calcul de mesures
de diffusion qui renseignent indirectement sur les propriétés microstructurales de la matière blanche (Alexander, Lee, Lazar, & Field, 2007; Mori & Zhang, 2006) (Figure 2C). Ces mesures comprennent l’anisotropie fractionnelle (« fractionnal anisotropy », FA), la diffusivité axiale (« axial diffusivity », AD), la diffusivité radiale (« radial diffusivity », RD) et la diffusivité moyenne (« mean diffusivity », MD), aussi appelée coefficient apparent de diffusion. La FA est l’une des mesures les plus utilisées. Elle varie entre 0 et 1 et exprime le degré de diffusion de l’eau qui contribue à l’anisotropie, mais sans fournir d’information sur l’orientation de la diffusion. Des valeurs de FA élevées indiquent une diffusion anisotropique alors que des valeurs de FA faibles indiquent une diffusion isotropique. L’AD correspond à la valeur propre principale du tenseur de diffusion et réfère ainsi à l’axe principal de
8
diffusion. Des valeurs d’AD élevées suggèrent une diffusion plus unidirectionnelle dans l’axe principal de diffusion. Une structure « intacte » de la matière blanche peut se refléter par des valeurs de FA ou AD élevées. La RD est la moyenne des valeurs propres secondaires du tenseur de diffusion. De valeurs faibles de RD suggèrent une diffusion moins importante dans les axes secondaires. Finalement, la MD est la moyenne de toutes les valeurs propres du tenseur de diffusion. Une structure « intacte » de la matière blanche peut se refléter par des valeurs de RD ou MD faibles. Des changements associés à chacune de ces métriques ou à des combinaisons de ces métriques peuvent révéler des phénomènes biologiques distincts tels que l’inflammation, la démyélinisation, l’œdème, la neuroplasie, etc. (Alexander et al., 2007).
Figure 2. Représentation d’un tenseur de diffusion et du calcul des différentes métriques de diffusion. (A) Représentation mathématique du tenseur de diffusion correspondant à une matrice symétrique 3 x 3 obtenue par l’acquisition d’au moins 6 directions de diffusion. (B) Valeurs propres du tenseur de diffusion (λ1, λ2, λ3) représentant la diffusion des molécules d’eau dans trois directions linéairement indépendantes de l’espace. (C) Calcul des métriques de diffusion à partir des valeurs propres du tenseur de diffusion.
Bien que la DTI soit grandement utilisée en clinique et en recherche, elle présente une limite importante rendant l’interprétation des mesures difficiles. En effet, la DTI ne permet pas de caractériser correctement la diffusion des molécules d’eau dans des régions où l’architecture de la matière blanche est complexe, comme un milieu dans lequel plusieurs fibres de matière blanche se
9
croisent ou se touchent (Alexander, Hasan, Lazar, Tsuruda, & Parker, 2001; Tuch et al., 2002). Le phénomène de croisement de fibres peut avoir un impact très important sur les mesures obtenues, car au moins 60 % des voxels de matière blanche contiennent des fibres qui se croisent (Jeurissen, Leemans, Tournier, Jones, & Sijbers, 2013). Notamment, la FA est grandement affectée par les régions de croisement de fibres.
1.1.3.2 Imagerie à haute résolution angulaire
Le développement récent de techniques d’IRM de type HARDI permet désormais aux algorithmes de tractographie de reconstruire plusieurs populations de fibres qui se croisent dans un même voxel. Une acquisition de type HARDI nécessite l’acquisition de plusieurs directions de diffusion (plus de 50) contrairement à l’acquisition de type DTI qui ne nécessite qu’un minimum de 6 directions de diffusion. La résolution angulaire fournit donc un modèle plus précis de la diffusion des molécules d’eau dans un même voxel comparativement à la DTI et offre ainsi une alternative plus robuste au phénomène de croisement de fibres (Raffelt et al., 2012).
Il existe plusieurs méthodes pour reconstruire l’architecture des fibres de matière blanche à partir des données HARDI. Une approche consiste à estimer la densité apparente des fibres (« apparent fiber density », AFD) dans chaque voxel (Descoteaux, Angelino, Fitzgibbons, & Deriche, 2006; Raffelt et al., 2012) à l’aide de la fonction d’orientation de distribution de fibres (« fibre Orientation Distribution Function », fODF). La fODF une fonction de la probabilité de la présence de fibres dans chaque orientation d’un voxel. L’AFD représente la fraction de l’espace occupé par un faisceau de fibres et représente un marqueur des lésions axonales (Raffelt et al., 2012). Par exemple, une diminution significative de l’AFD a été observée dans le faisceau corticospinal et le corps calleux chez un groupe de patients atteints d’une maladie affectant les motoneurones comparativement à un groupe de participants en santé (Raffelt et al., 2012). Une mesure complémentaire à l’AFD est le nombre d’orientations de fibres (« number of fiber orientations », NuFO). Cette mesure représente le nombre d’orientations de fibres dans chaque voxel de l’image (Dell'Acqua, Simmons, Williams, & Catani, 2013). Cette mesure est un indicateur de la complexité de la matière blanche. Une diminution de NuFO indique ainsi une diminution de la complexité de la matière blanche.
10
1.2 Vieillissement cérébral
Le vieillissement cérébral est associé à de multiples changements structurels, dont beaucoup peuvent être visualisés par l’IRM structurelle et de diffusion. Les deux prochaines sections fourniront un bref aperçu des différences observées dans le cerveau d’adultes âgés comparativement à celui de jeunes adultes à l’aide de ces deux techniques.
1.2.1 Imagerie par résonance magnétique structurelle
Plusieurs études d’IRM structurelle ont investigué les trajectoires de changements volumétriques des trois types de tissus cérébraux (matière grise, matière blanche et liquide cérébro-spinal) au cours du vieillissement normal (p. ex. Allen, Bruss, Brown, & Damasio, 2005; Bajaj, Alkozei, Dailey, & Killgore, 2017; Courchesne et al., 2000; Good et al., 2001; Guttmann et al., 1998; Liu et al., 2016; Pfefferbaum et al., 1994; Raz et al., 1997; Scahill et al., 2003). Certaines de ces études révèlent que le volume du cerveau diminue au cours du vieillissement normal (Courchesne et al., 2000; Scahill et al., 2003). Par exemple, Courchesne et al. (2000), en comparant 116 volontaires âgés entre 16 mois et 80 ans, ont observé une diminution linéaire, mais lente, du volume du cerveau entre 16 ans et 80 ans. L’étude longitudinale de Scahill et al. (2003), en comparant la structure cérébrale de 39 volontaires âgés entre 39 et 84 ans, a identifié un taux de diminution du volume du cerveau de 0,32 % par année. Plusieurs de ces études suggèrent également que le volume de la matière grise diminue linéairement avec l’âge (Allen et al., 2005; Bajaj et al., 2017; Courchesne et al., 2000; Good et al., 2001; Pfefferbaum et al., 1994). Courchesne et al. (2000) ont notamment observé une diminution de 5 % par décennie à partir de l’âge de 10 ans. Des études ont aussi observé que le volume de la matière blanche diminue également avec l’âge, soit à partir de 40 (Courchesne et al., 2000), 50 (Allen et al., 2005; Liu et al., 2016) ou 59 ans (Guttmann et al., 1998), alors que d’autres études ne révèlent aucun changement significatif du volume de la matière blanche avec l’âge (Good et al., 2001; Pfefferbaum et al., 1994). De plus, le déclin du volume de la matière grise et de la matière blanche au cours du vieillissement semble largement hétérogène à travers le cerveau avec des patrons de vieillissement variant d’une région à l’autre (Allen et al., 2005; Liu et al., 2016; Raz et al., 1997; Raz et al., 2005). Toutefois, de manière générale, ces études reflètent une vulnérabilité des régions du lobe frontal comparativement aux régions du lobe occipital. Enfin, pour le liquide cérébro-spinal, il a été observé que le volume du liquide cérébro-spinal ventriculaire ou extra-ventriculaire augmente avec
11
l’âge (Courchesne et al., 2000; Guttmann et al., 1998; Pfefferbaum et al., 1994; Scahill et al., 2003). Notamment, l’équipe de Scahill et al. (2003) a observé une augmentation du volume du liquide cérébro-spinal ventriculaire de 650 mm3 par année ; un taux qui s’accélère significativement après
l’âge de 70 ans. Cette augmentation liée à l’âge du volume du liquide cérébro-spinal corrèle fortement avec l’augmentation liée à l’âge du volume de la matière blanche (Guttmann et al., 1998), suggérant un mécanisme commun dans les changements de la structure du cerveau au cours du vieillissement normal.
D’autres changements dans la structure du cerveau ont été identifiés à l’aide des métriques de SBM (surface, volume et épaisseur). Une diminution globale de l’épaisseur corticale a notamment été observée dans plusieurs études (p. ex. Bajaj et al., 2017; Fjell et al., 2009; Lemaitre et al., 2012; Long et al., 2012; Salat et al., 2004; Zheng et al., 2019), mais de façon plus marquée dans les régions des lobes frontal et temporal (Lemaitre et al., 2012; Salat et al., 2004) ainsi que dans les régions du lobe pariétal et les régions sous-corticales (Long et al., 2012). Toutefois, des résultats de la littérature suggèrent que le vieillissement normal peut également être associé à une augmentation de l’épaisseur corticale dans le cortex dorsolatéral préfrontal et le cortex cingulaire antérieur (Dotson et al., 2015). Cette étude propose qu’une augmentation de l’épaisseur corticale liée à l’âge pourrait refléter un processus d’inflammation chronique. De plus, le vieillissement normal a aussi été associé à une diminution de la surface corticale (p. ex. Dotson et al., 2015; Lemaitre et al., 2012; Long et al., 2012) dans plusieurs régions. La surface corticale est une mesure qui semble toutefois moins affectée par le vieillissement que l’épaisseur corticale (Long et al., 2012). À titre indicatif, l’étude de Lemaitre et al. (2012) a identifié une réduction de l’épaisseur corticale moyenne de 0,004 mm par an et une réduction de la surface corticale totale de 3,68 cm2 par an.
En résumé, les études d’IRM structurelle révèlent majoritairement une vulnérabilité de la structure (volume, surface et épaisseur) des trois tissus cérébraux au cours du vieillissement normal avec des trajectoires de changements distinctes pour la matière grise, la matière blanche et le liquide cérébro-spinal. D’autres changements dans la structure du cerveau peuvent également être observés à l’aide de l’IRM de diffusion.
12
Des changements liés à l’âge dans la structure des faisceaux de matière blanche sont observés à l’aide de l’IRM de diffusion. De manière générale, les études révèlent une diminution de la FA dans plusieurs régions cérébrales et faisceaux de matière blanche (e.g. Bennett, Madden, Vaidya, Howard, & Howard, 2010; Burzynska et al., 2010; Head et al., 2004; Hsu et al., 2008; Pelletier et al., 2015; Pfefferbaum, Adalsteinsson, & Sullivan, 2005; Salat et al., 2005). Plusieurs de ces études identifient la présence d’un gradient antéro-postérieur dans le patron de diminution de la FA (Bennett et al., 2010; Burzynska et al., 2010; Head et al., 2004; Pelletier et al., 2015; Pfefferbaum et al., 2005; Salat et al., 2005), c’est-à-dire que les régions antérieures sont plus affectées par le vieillissement que les régions postérieures, supportant la vulnérabilité des régions frontales face au vieillissement normal.
Par ailleurs, les autres mesures de diffusion (RD, MD et/ou AD) augmentent dans plusieurs régions cérébrales et faisceaux de matière blanche au cours du vieillissement normal (p. ex. Bennett et al., 2010; Burzynska et al., 2010; Head et al., 2004; Hsu et al., 2008; Pelletier et al., 2015; Pfefferbaum et al., 2005). Une diminution de la FA accompagnée d’une augmentation des autres mesures de diffusion peut refléter plusieurs processus distincts (p. ex. Arfanakis et al., 2002; Sun et al., 2006), dont une démyélinisation (FA↓ / RD↑) et une dégénérescence axonale (FA↓ / AD↑). Ce type de changements dans la structure de la matière blanche peut altérer la conduction des signaux neuronaux dans tout le système nerveux central, et ainsi entraîner une transmission plus lente ou incomplète des impulsions neuronales (Bartzokis, 2004). Bien que ces deux patrons discutés plus haut soient souvent observés dans la littérature, d’autres combinaisons de changement sont également présentes (p. ex. FA↓ MD↓ AD↓ pouvant réfléter une augmentation de la densité de la membrane cellulaire : Burzynska et al., 2010).
Une seule étude a investigué le vieillissement normal de la matière blanche à l’aide de la HARDI (Tremblay et al., 2019). Dans cette étude, effectuée par notre groupe de recherche, l’avancement en âge a été associé à des effets positifs sur la structure du faisceau arqué (AF, « arcuate fasciculus ») et du faisceau longitudinal moyen, soit une augmentation des valeurs d’AFD et de NuFO. Les auteurs expliquent ces résultats par le fait que le groupe d’adultes âgés était hautement scolarisé. Bien que des effets positifs soient observés sur les métriques de fODF, des effets négatifs liés au vieillissement ont été observés sur les métriques de DTI (↓FA ↑MD).
En somme, à l’aide de l’IRM structurelle et de diffusion, de nombreux changements hétérogènes sur la structure du cerveau au cours du vieillissement normal ont été identifiés. Comme
13
il sera discuté à la fin du chapitre 2, plusieurs de ces changements ont été associés et corrélés à des différences de PPb. Auparavant, le chapitre 2 détaillera les facteurs impliqués dans la capacité à percevoir la parole, les modèles neurobiologiques de la perception de la parole ainsi que l’effet du vieillissement sur cette capacité.
14
Chapitre 2 Perception de la parole et vieillissement
2.1 Perception de la parole
2.1.1 Définition
La perception de la parole repose sur plusieurs étapes de traitement distinctes et complexes, impliquant le traitement auditif périphérique et central, le traitement cognitif et le traitement de la parole
Pour percevoir la parole, le signal de la parole produit par un locuteur, qui est, entre autres, hautement acoustiquement variable, doit d’abord être capté par le système auditif périphérique. Le système auditif périphérique comprend les trois compartiments de l’oreille (externe, moyenne, interne) et le nerf auditif. Il est responsable de capter et de transmettre les ondes sonores sous forme d’influx nerveux au système auditif central. Le système auditif central comprend plusieurs relais neuronaux situés dans le tronc cérébral et le thalamus qui ont pour objectif d’extraire les propriétés du signal acoustique et de sa localisation. De façon simplifiée, le nerf auditif innerve les noyaux cochléaires ventral et dorsal situés dans le tronc cérébral. Les cellules du noyau cochléaire ventral projettent ensuite bilatéralement dans le noyau olivaire supérieur du tronc cérébral, qui projette à son tour dans le colliculus inférieur du mésencéphale. Les cellules du colliculus inférieur innervent par la suite le corps genouillé médian du thalamus, qui innerve le cortex auditif primaire (gyrus de Heschl) gauche et droit. Le cortex auditif primaire correspond à la porte d’entrée du signal acoustique dans le cortex. Il est responsable d’intégrer les informations reçues et d’effectuer une analyse spectrotemporelle du signal acoustique, c’est-à-dire d’extraire les propriétés spectrales (fréquences) et temporelles (variations dans le temps) de tous les sons, incluant la parole.
Les propriétés spectrales et temporelles de la parole doivent continuellement être gardées en mémoire et mises en relation avec des représentations linguistiques. La perception de la parole fait référence aux mécanismes par lesquels les propriétés du signal acoustique sont mises en correspondance avec des représentations linguistiques, qu’il s’agisse de phonèmes ou de syllabes. Ces représentations linguistiques doivent ensuite être assemblées de manière à reconnaître des mots pour finalement accéder à la signification de ces mots et les combiner pour comprendre le sens de ce qui est dit. Le phénomène par lequel un sens est donné aux représentations linguistiques fait référence au processus de compréhension de la parole.
15
Plusieurs auteurs ont modélisé les différentes étapes du traitement de la parole dans le cerveau sous la forme d’un traitement à double voie (Fridriksson et al., 2016; Hickok & Poeppel, 2007; Rauschecker & Scott, 2009). Une voie serait impliquée en compréhension de la parole et l’autre voie serait potentiellement impliquée en perception de la parole selon les modèles/auteurs. Avant d’aborder ces modèles neurobiologiques (section 2.1.3), les différents paradigmes expérimentaux utilisés pour étudier la perception de la parole seront détaillés à la section suivante.
2.1.2 Paradigmes expérimentaux de perception de la parole
Les tâches de perception de la parole utilisées en recherche incluent principalement la répétition, l’identification et la discrimination (McGuire, 2010). Tous ces paradigmes consistent à présenter des sons dans les deux oreilles, dans la même oreille ou dans des oreilles différentes. La présentation des sons peut se faire soit à l’aide d’un casque d’écoute soit en champ libre à l’aide de haut-parleurs positionnés à différentes distances et dans différents angles.
Répétition : La plupart des tâches de perception développées et utilisées en recherche incluent de la répétition d’unités linguistiques en présence ou en l’absence de bruit. Par exemple, le « Quick Speech in Noise Test » (Killion, Niquette, Gudmundsen, Revit, & Banerjee, 2004), le « Hearing in Noise Test » (Nilsson, Soli, & Sullivan, 1994) et le « Bamford-Kowal-Bench Speech-in-Noise Test » (Bench, Kowal, & Bamford, 1979) impliquent de la répétition de phrases. D’autres tests tels que le « Arthur Boothroyd words Test » (Boothroyd, 1984) et le « Words in Noise Test » (Wilson, 2003) impliquent de la répétition de mots. Récemment, un test impliquant de la répétition de phrases en français nommé « Test de Phrases dans le bruit » a été développé (Lagacé, Jutras, Giguère, & Gagné, 2010) et testé auprès de 29 jeunes adultes et 38 adultes âgés (Lagace, Geffray, & Gagné, 2017). Bien que le paradigme de répétition soit largement utilisé dans la littérature, il présente une limite importante. En effet, une tâche de répétition nécessite une perception de la parole préservée, mais également un réseau articulatoire intact. Ainsi, une erreur d’articulation liée au vieillissement pourrait être interprétée comme une erreur de perception de la parole. D’autres paradigmes tels que la discrimination et l’identification permettent d’étudier plus précisément la perception de la parole sans contribution du réseau articulatoire.
16
Discrimination : Les tâches de discrimination consistent à présenter deux ou plusieurs unités linguistiques, identiques ou différentes, dans un court laps de temps. La tâche consiste précisément à indiquer si les unités présentées sont différentes ou identiques. Plusieurs variantes de ce paradigme existent (McGuire, 2010). La forme la plus simple (AX) consiste à indiquer si deux stimuli sont identiques ou différents. Une autre variante (AXB) consiste à présenter trois unités et à indiquer si l’unité du milieu est identique à la première ou à la dernière unité. Finalement, d’autres versions consistent à présenter deux unités et à indiquer quelle unité a été présentée en premier/dernier, ou à présenter deux paires d’unités et à indiquer quelle paire contient des unités différentes. Plus le nombre d’unités présentées augmente, plus la tâche devient difficile et nécessite l’implication de la mémoire de travail afin de retenir et de comparer tous les éléments. Dans le cadre d’études sur le vieillissement, étant donné que la mémoire de travail verbale subit un déclin avec l’âge (e.g. Bopp & Verhaeghen, 2005), l’emploi de tâches de discrimination plus simple (p.ex. AX) est à prioriser afin d’éliminer tous facteurs confondants.
Identification : Les tâches d’identification consistent quant à elle à présenter une unité linguistique et à identifier l’unité présentée parmi un choix ouvert ou fermé (McGuire, 2010). La forme la plus simple de la tâche d’identification consiste à indiquer si un stimulus était présent ou non ou si le stimulus était /ba/ ou /da/, par exemple. Dans une autre version, la tâche d’identification consiste à présenter un seul stimulus à chaque essai et à demander au sujet d’appliquer une étiquette au stimulus, soit à partir d’un ensemble fermé ou d’un ensemble ouvert de choix. Tout comme pour les tâches de discrimination, il est important d’utiliser des tâches d’identification simples dans le cadre d’études sur le vieillissement. Par exemple, un trop grand nombre de choix dans un ensemble fermé peut rendre la tâche plus difficile à comprendre et faire appel à une plus grande capacité de mémoire de travail afin de comparer l’unité présentée avec l’ensemble des choix.
Tous ces paradigmes permettent d’étudier les différents processus impliqués dans le traitement de la parole. L’utilisation de l’un ou l’autre de ces paradigmes dépend des objectifs et des populations à l’étude. Par exemple, une étude voulant étudier l’implication de la mémoire de travail en perception de la parole pourrait opter pour une tâche de discrimination ou d’identification complexe. De plus, selon les unités linguistiques présentées, les paradigmes peuvent nécessiter à la fois les processus de perception de la parole et de compréhension de la parole alors que d’autres peuvent nécessiter seulement le processus de perception de la parole. Notamment, la voie de la compréhension de la parole est davantage impliquée lorsque les unités linguistiques utilisées sont à
17
un niveau lexical (p. ex. mots ou phrases) que lorsque les unités linguistiques sont à un niveau sous-lexical (p. ex. non-mots, syllabes ou phonèmes). Selon des modèles neurobiologiques, le traitement de la perception de la parole et celui de la compréhension de la parole s’effectueraient dans deux voies distinctes (Fridriksson et al., 2016; Hickok & Poeppel, 2007; Rauschecker & Scott, 2009) : la voie dorsale et la voie ventrale, respectivement. Les modèles neurobiologiques de ces voies, ainsi que les rôles de chaque voie dans le traitement de la parole sont abordés dans la prochaine section.
2.1.3 Modèles à deux voies
Le premier modèle neurobiologique du traitement de la parole a été proposé au 19e siècle par
Carl Wernicke et a ensuite été élaboré par plusieurs autres scientifiques. Vers la fin du 20e siècle, le
modèle a été modifié et popularisé par le neurologue américain Norman Geschwind (Geschwind, 1970). Ce modèle dit « Broca-Wernicke-Litchtheim-Geschwind » comprend un centre de production de la parole situé dans le lobe frontal inférieur gauche et un centre de compréhension de la parole situé dans le lobe temporal supérieur gauche. Ces régions seraient connectées par l’intermédiaire du AF. Toutefois, de nombreuses études en neuroimagerie révèlent que le traitement de la parole implique davantage de régions cérébrales et de faisceaux de matière blanche que ce qui est identifié par le modèle Broca-Wernicke-Litchtheim-Geschwind (pour des revues, voir Dick, Bernal, & Tremblay, 2014; Tremblay & Dick, 2016).
Des modèles du traitement de la parole à double voie ont notamment affiné notre compréhension des bases neurobiologiques du traitement de la parole (Fridriksson et al., 2016; Hickok & Poeppel, 2007; Rauschecker & Scott, 2009). Ces modèles proposent plusieurs sous-étapes s’effectuant dans différentes régions cérébrales. À la suite de l’analyse spectrotemporelle du signal acoustique décrite dans la section 2.1.1, le traitement de la parole s’effectuerait à l’intérieur de deux voies ayant des fonctions différentes : une voie ventrale et une voie dorsale. Le traitement dans les deux voies s’effectuerait parallèlement (Fridriksson et al., 2016; Hickok & Poeppel, 2007) ou cycliquement (Rauschecker & Scott, 2009). Bien que les régions impliquées dans les deux voies et les différentes étapes proposées par chacun des modèles diffèrent, les bases sont similaires. En effet, les modèles proposent l’existence d’une voie ventrale connectant les gyri/sulci temporaux moyen, inférieur et/ou postérieur au gyrus frontal inférieur. Les faisceaux de matière blanche qui ont été proposés pour relier ces régions sont le faisceau unciné, la capsule extrême, le faisceau longitudinal moyen, le faisceau longitudinal inférieur et le faisceau fronto-occipital inférieur. La voie ventrale