Taxonomic and functional traits responses of Sphagnum peatland testate amoebae to experimentally manipulated water table
10
0
0
Texte intégral
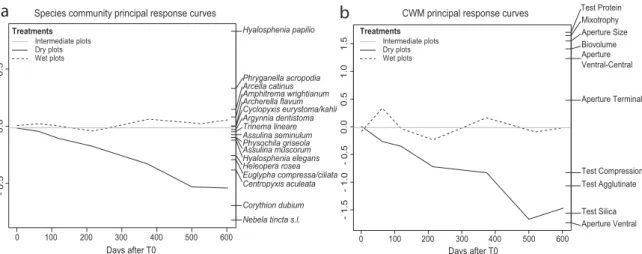
Figure
Documents relatifs
