Stability and Change of Social Relationship Quality in Captive Chimpanzees ( Pan troglodytes )
17
0
0
Texte intégral
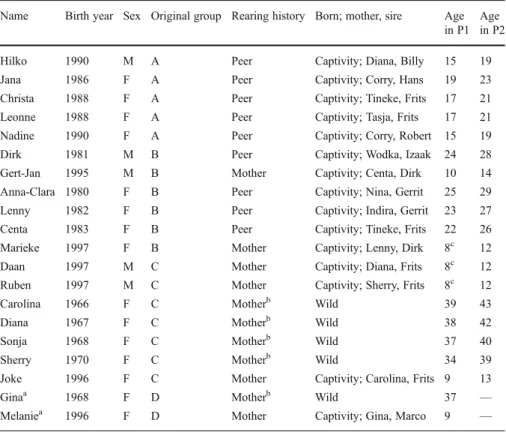
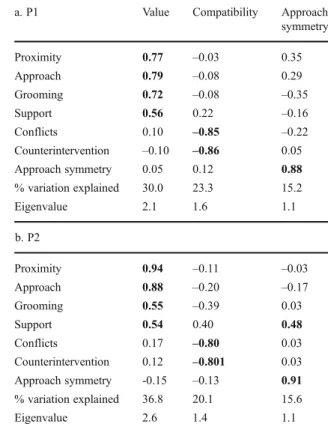
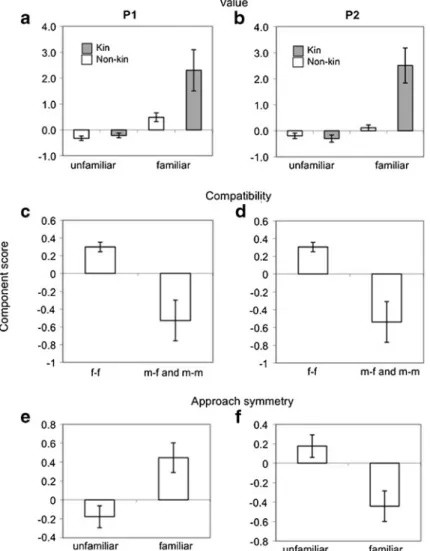
Figure
Documents relatifs