HAL Id: hal-02832728
https://hal.inrae.fr/hal-02832728
Submitted on 7 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Modélisation des chutes de litières en quantité et qualité
pour des clones d’Eucalyptus au Congo
Rémi d’Annunzio
To cite this version:
Rémi d’Annunzio. Modélisation des chutes de litières en quantité et qualité pour des clones d’Eucalyptus au Congo. Sciences du Vivant [q-bio]. 2004. �hal-02832728�
Diplôme d’Etudes Approfondies
de Biologie Forestière
Présenté le 2 septembre 2004 par
Rémi D’ANNUNZIO
Président du Jury : P. Dizengremel Rapporteurs : D. Epron, J.-C. Hervé
Responsable de stage : Laurent SAINT-ANDRE
L
L
aboratoire d’E
E
tude desR
R
essourcesF
F
O
O
rêt-B
B
oisUnité Mixte de Recherches INRA-ENGREF 1092
Modélisation des chutes de litières en quantité et
qualité pour des clones d’Eucalyptus au Congo
Résumé :
L’objectif de cette étude était d’intégrer le compartiment chute de litière à la chaîne de modèles dynamique de croissance Eucalypt Dendro mise au point pour les plantations clonales d’Eucalyptus au Congo
Une approche qualitative a également été développée pour dégager les principales caractéristiques de la litière à travers une expérience comparative de décomposition en conditions contrôlées. Trois critères ont été utilisés : le rapport C:N, la vitesse de décomposition et un facteur de qualité intégré.
Deux modèles de chutes ont été établis, au pas de temps annuel et mensuel. Le modèle annuel tient compte de l’effet de l’âge et peut désormais être intégré à la chaîne. Le modèle mensuel utilise des données climatiques et dendrométriques avec un bon pouvoir de prédiction, mais son intégration à la chaîne ne pourra se faire qu’après avoir introduit le climat et son influence sur la croissance en hauteur et en diamètre dans Eucalypt-Dendro.
L’étude qualitative révèle que l’origine clonale et les caractéristiques stationnelles ont un effet fort sur la qualité de la litière.
Des études complémentaires sur la décomposabilité de la litière devront être menées sur le terrain pour vérifier ces conclusions et pour appréhender dans son ensemble le processus de relaxation des éléments minéraux.
Mots-clefs : eucalyptus, cycles biogéochimiques, chutes de litière, décomposition, modèle de croissance
Abstract :
The aim of this study was to develop a litterfall model that could be dealt with by the parameters of a dynamic growth model (Eucalypt Dendro) fitted for eucalypt plantations. A parallel qualitative approach was undertaken to determine which were the principal characteristics of the litter quality through a decomposition experiment led in laboratory. Three criterions were considered : the decomposition rate, the C:N ratio and an integrative quality index.
Two models have been developed : a yearly one that can immediately be incorporated in the growth model and a monthly one which enables to understand how climatic and dendrometric factors influence litterfalls. However this model will not be used at once, for it needs the increases of basal area which are not included yet at a monthly pace (with climatic variations) in the chain model.
The qualitative study revealed that there was a strong effect of the genetic origin and the stand characteristics on litter quality.
Further studies are needed to check whether these result can be extended to field experiments. Key-words : eucalypt, nutrient cycles, litterfall, decomposition, growth model
1. INTRODUCTION_________________________________________________________________ 1
1.1 La durabilité des plantations et le maintien de la fertilité dans les sols : un enjeu pour la bio séquestration du carbone. _________________________________________________________________________________ 1 1.2 Présentation et objectifs du stage___________________________________________________________ 1
2. ELEMENTS BIBLIOGRAPHIQUES ET HYPOTHESES DE TRAVAIL __________________ 2
2.1 Généralités____________________________________________________________________________ 2 2.2 Aspect quantitatif_______________________________________________________________________ 2 2.3 Aspect qualitatif________________________________________________________________________ 3 3. MATERIEL ET METHODES ______________________________________________________ 3 3.1 Caractéristiques stationnelles______________________________________________________________ 3 3.2 Matériel végétal ________________________________________________________________________ 4 3.3 Quantification des chutes_________________________________________________________________ 4 3.3.1 Caractéristiques des parcelles _________________________________________________________ 4 3.3.2 Méthode de récolte des litières ________________________________________________________ 4 3.3.3 Données disponibles ________________________________________________________________ 5 3.3.4 Homogénéisation ___________________________________________________________________ 5 3.4 Qualité de la litière _____________________________________________________________________ 5 3.4.1 Caractéristiques des parcelles _________________________________________________________ 5 3.4.2 Méthode de récolte des feuilles ________________________________________________________ 6 3.4.3 Décomposition en laboratoire _________________________________________________________ 6 3.4.4 Analyses spectrophotométriques _______________________________________________________ 7 3.4.5 Analyses chimiques _________________________________________________________________ 8 3.4.6 Analyses physiques _________________________________________________________________ 8 3.4.6.1 Epaisseur des feuilles (e) _________________________________________________________ 8 3.4.6.2 Masse surfacique (LMA pour leaf mass area)_________________________________________ 9 3.4.6.3 Densité _______________________________________________________________________ 9
4. RESULTATS_____________________________________________________________________ 9
4.1 Modélisation des chutes de litières _________________________________________________________ 9 4.1.1 Analyse des chutes annuelles, effet de l’âge des arbres______________________________________ 9 4.1.2 Analyse des chutes mensuelles, effet du climat et de la croissance ___________________________ 10 4.1.2.2 Modèle pour les chutes de feuilles _________________________________________________ 10 4.2 Qualité des litières en fonction de la provenance et la fertilité ___________________________________ 12 4.2.1 Analyse des propriétés des feuilles avant décomposition ___________________________________ 12 4.2.2 Décomposition en microcosme _______________________________________________________ 12 4.2.2.1 Analyse spectrophotométrique____________________________________________________ 12 4.2.2.2 Profils de décomposition et modélisation ___________________________________________ 12
5 DISCUSSION ____________________________________________________________________ 15
5.1 Modélisation _________________________________________________________________________ 15 5.1.1 Modèle de chutes annuelles __________________________________________________________ 15 5.1.2 Modèle de chutes foliaires mensuelles__________________________________________________ 16 5.1.2.1 Choix des paramètres ___________________________________________________________ 16 5.1.2.2 Analyse des résidus ____________________________________________________________ 16 5.1.2.3 Perspectives __________________________________________________________________ 16 5.2 Qualité de la litière foliaire ______________________________________________________________ 17 5.2.1 Protocole d’expérience _____________________________________________________________ 17 5.2.2 Méthodes statistiques_______________________________________________________________ 17 5.2.3 Effet clone _______________________________________________________________________ 19 5.2.4 Effet station ______________________________________________________________________ 19 6 CONCLUSION __________________________________________________________________ 20 Bibliographie ______________________________________________________________________ 21
1. INTRODUCTION
1.1 LA DURABILITE DES PLANTATIONS ET LE MAINTIEN DE LA FERTILITEDANS LES SOLS : UN
ENJEU POUR LA BIO SEQUESTRATION DU CARBONE.
Depuis la conférence de Rio en 1992 portant sur la réduction des émissions de gaz à effet de serre (CO2, CH4, N2O), la communauté internationale s’est beaucoup investie pour étudier le potentiel de l’afforestation comme moyen de stocker durablement du carbone. Par le biais de la photosynthèse, le CO2 atmosphérique est fixé par les plantes ligneuses pour créer de la biomasse et le carbone gazeux est transformé en carbone organique. Ces ligneux sont ainsi considérés comme des réservoirs de carbone qui peuvent devenir importants à l’échelle des plantations. Par exemple l’installation d’un peuplement boisé d’eucalyptus sur une végétation originelle de type savane permet un stockage moyen de 7.5 tC/ha/an répartis en 6 t pour le compartiment aérien et 1.5 t en souterrain (Laclau 2001). Ce bilan ne tient pas compte des turn-overs (notamment les chutes de litière aériennes) ni de la respiration et leur étude est nécessaire afin d’établir un bilan de carbone ecosystèmique. De plus cette biomasse sur pied ne peut être considérée comme un puit réel que si la plantation est durable, c’est-à-dire si l’exploitation sur plusieurs générations est pérenne, en particulier si elle ne change pas la fertilité du sol sur lequel elle est installée. En cas de production trop intensive (durée de rotation courte, exportation des rémanents…) la quantité d’éléments minéraux disponibles pour la nutrition des arbres est susceptible de baisser. Or la fertilité est corrélée positivement à la biomasse potentielle et si en fin d’exploitation la fertilité a baissé, la capacité de la plantation à stocker le carbone se trouve diminue. En sus des implications d’une exploitation sylvicole à haut rendement sur la biodiversité du site, l’équilibre du bilan d’éléments minéraux nutritifs de l’écosystème apparaît comme fondamental pour la durabilité d’une plantation et sa capacité à séquestrer le carbone à long terme. La compréhension des cycles biogéochimiques apparaît sous cet angle comme un facteur clef dans l’élaboration des modèles décrivant le fonctionnement et la dynamique des peuplements forestiers.
1.2PRESENTATION ET OBJECTIFS DU STAGE
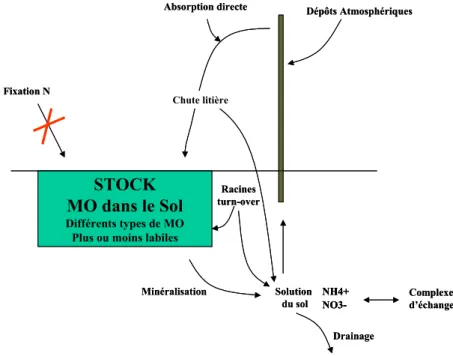
Depuis 1978, 42000 hectares de clones d’Eucalyptus ont été plantés sur le littoral de la région de Pointe Noire au Congo, pour la production de pâte à papier essentiellement. Ces plantations se trouvent sur un sol sableux avec de faibles réserves en éléments minéraux et une faible capacité de rétention d’eau (Laclau, 2001). En ce qui concerne le régime sylvicole et la gestion des ressources du sol, elles peuvent être considérées comme un cas extrême de ligniculture conduite sur très courte rotation (7ans). Il existe dans ces conditions un risque très élevé de déficit en éléments nutritif, même à moyen terme. C’est dans ce cadre que des recherches ont été menées depuis juin 2000 pour mettre au point un modèle dynamique des peuplements, qui vise à évaluer la productivité et la durabilité des plantations d’eucalyptus au Congo (Saint André et al. 2002). Schématiquement, la chaîne de modèles Eucalypt-Dendro comprend trois modules (figure 1), qui gèrent :
1/ la croissance des arbres. Ce module reprend une grande partie du modèle de croissance Fagacées, élaboré pour des futaies monospécifiques équiennes de hêtre et de chêne (Dhôte, 1996 ; Dhôte, 1990) ;
2/ la biomasse sur pied et les propriétés physiques du bois ;
3/ la teneur en éléments minéraux dans les différents compartiments de l’arbre.
Dans le contexte de Pointe-Noire, les apports atmosphériques sont faibles (le niveau de pollution industrielle est bas, les vents maritimes amènent surtout des sels), et peu d’éléments proviennent de l’altération de la roche (Nzila et al. 2003). Les éléments sont prélevés dans le sol pendant les premières phases de croissance puis recyclés par le biais des chutes de litière. On estime pour le clone PF1-41 que selon l’âge, 10 à 50 % des éléments prélevés pour sa croissance retournent au
sol via les chutes de litière alors que 2% au maximum proviennent d’apports atmosphériques (Laclau, 2001).
L’objectif principal de ce stage est de traiter le retour au sol des éléments minéraux via la quantification des chutes de litière. Au niveau de la modélisation il s’agit d’intégrer le compartiment chute de litière en sortie du module biogéochimique (figure 1).
Le deuxième objectif est de déterminer quels sont les facteurs qui influent sur la qualité de la litière et dans quelle mesure ils interviennent sur la quantité d’éléments minéraux bio-disponibles pour la croissance de l’arbre.
2. ELEMENTS BIBLIOGRAPHIQUES ET HYPOTHESES DE TRAVAIL
2.1GENERALITES
Les études sur les chutes de litière sont assez contraignantes en terme de protocole : elles exigent d’être faites sur des temps de suivi relativement longs et dans des conditions de croissance bien maîtrisées afin d’isoler les paramètres influant sur la variabilité observée. Pour ces raisons, il n’existe encore que peu de données disponibles permettant de développer des modèles de dynamique des chutes.
Une synthèse bibliographique portant sur différents peuplements homogènes en milieu tempéré et tropical a toutefois permis de dégager les points suivants :
1/ les quantités de chute litière sont une fonction croissante de l’âge avec un plafonnement ; 2/ il existe autour de cette tendance une forte variabilité inter-annuelle qui augmente avec l’âge. Comme ce schéma se retrouve pour diverses essences avec du matériel végétal contrôlé (peuplements équiennes et monospécifiques, matériel clonal etc.) et dans des conditions de fertilité et de densité maîtrisées, cette variabilité est a priori dues à des facteurs environnementaux (pluviométrie, vent, sécheresse, pathogènes) ;
L’exemple présenté sur les figures 3 et 4 concerne un peuplement monospécifique de Douglas (pseudotsuga menziesii) suivi pendant neuf années consécutives et illustre bien les remarques précédentes (Trofymow, 1991). On retrouve ces caractéristiques dans la plupart des études, en particulier pour l’eucalyptus au Congo (Laclau 2001).
2.2ASPECT QUANTITATIF
A partir de ces constatations, il a été possible de déterminer le type de données requises pour développer un modèle de chute de litière : des données issues des chronoséquences (peuplements d’âges différents installés sur des sites de fertilité équivalente et suivis sur une année) afin de traiter l’effet âge et des données longitudinales (peuplements suivis sur une longue période de temps) pour analyser l’effet des saisons.
Les plantations congolaises sur lesquelles est installé du matériel végétal clonal permettent de satisfaire à ces contraintes :
1/ le rythme d’exploitation est suffisamment élevé pour pouvoir suivre les chutes de litière sur une longue période au regard de la durée de rotation ;
2/ le suivi des données de croissance a été effectué pour les mêmes peuplements sur des plages de temps correspondantes ;
3/ les données climatiques (pluviométrie, rayonnement) font l’objet d’un suivi journalier à l’aéroport de Pointe Noire depuis 1970 et depuis 2000 sur différentes parcelles du massif. Le modèle étant de type dendrométrique, on a cherché à développer le module de chute de litière sous un angle qui rende compte de la fertilité et des conditions croissances via la hauteur
dominante et la surface terrière. Des données climatiques brutes ont été associées à ces paramètres pour expliquer la variabilité qui existe sur les résidus autour de la tendance âge. L’objectif final est de développer un modèle de type Chute=f(âge, climat , croissance) via deux approches :
1/ annuelle en utilisant principalement des données de chronoséquence 2/ mensuelle en utilisant des données longitudinales.
2.3ASPECT QUALITATIF
Si on ne tient pas compte de l’effet de l’âge, la composition chimique de la litière est relativement stable d’une année sur l’autre et la variabilité observée chez Laclau (2001) et Trofymow (1991) semble être avant tout fonction de l’origine génétique, de la qualité de la station (fertilité) et/ou du régime sylvicole (fertilisation). Deux facteurs ont été étudiés pendant ce stage : la fertilité et la provenance génétique qui sont tous deux des paramètres d’entrée du modèle de croissance (la fertilisation n’est pas encore gérée dans la chaîne de modèles).
La notion de litière dépend du stade auquel elle est considérée : au moment de la chute, après être tombée dans les bacs où elle subit un lessivage important, ou pendant les différentes phases de décomposition. La composition chimique et la capacité à rendre les éléments minéraux disponibles dépendent en premier lieu de cette définition. Par exemple le lessivage entraîne un drainage de certains éléments minéraux accompagné d’une perte de matière organique qui peut atteindre plus de 25% pendant les premiers jours où la litière est au sol ou dans des bacs (Reversat,1999).
Pour éviter ce facteur de variabilité supplémentaire, nous nous sommes intéressés, dans cette étude, à la litière au moment de la chute.
Le terme de qualité de la litière qu’on trouve dans la littérature prend des sens différents selon les points de vue adoptés par les auteurs. Trois approches de la définition de la qualité ont été adoptées :
Elle peut être définie par le rapport C/N qui caractérise de façon globale la teneur en matière organique facilement décomposable par rapport aux éléments plus réfractaires. Berg and McClaugherty (2003) suggère que, pour la plupart des espèces, un taux de C:N bas dans la litière fraîche indique une vitesse de décomposition initiale élevée.
Au niveau moléculaire, les atomes de carbone sont répartis dans la litière en plusieurs fractions en fonction de leur degré de solubilité : hydrosolubles, solubles à l’acide et réfractaires à l’acide. Agren et Bossata (1996) ont défini la qualité de la litière par l’accessibilité des décomposeurs à ces différentes fractions via le paramètre qo.
La qualité peut également être estimée au niveau dynamique par le biais de la vitesse de décomposition notée ici v (Olson 1963).
Dans la suite de l’étude et sauf mention spéciale, le terme qualité regroupe ces trois paramètres : C/N, qo et v. Une expérience comparative de décomposition en microcosme a été réalisée sous humidité et température contrôlées pour déterminer comment ces paramètres sont influencés par l’origine génétique du clone et la fertilité de la station.
3. MATERIEL ET METHODES
3.1CARACTERISTIQUES STATIONNELLES
Le massif d’Eucalyptus est planté sur les savanes côtières qui couvrent la façade atlantique sur une trentaine de kilomètres de largueur, par 4° de latitude Sud et 12° de longitude Est (figure 5). Le régime pluviométrique de la région est du type équatorial de transition appelé "bas congolais" (Jamet et Rieffel, 1976) avec une température moyenne annuelle de 25°C et une humidité relative moyenne de l’air de 85% .
Les plantations d’Eucalyptus sont mises en place essentiellement sur les savanes dominées par
Loudetia arundinacea (Hoschst) et Hypparenia diplandra (Hack). Le substratum géologique est de
type Ferralic Arenosols (FAO), constitué d’épaisses formations détritiques meubles d’origine continentale datées du plio-pléistocène (Jamet, 1967). D’un point de vue granulométrique, les sols de la région côtière sont essentiellement sableux (80-90%), les teneurs en argile (8-10 %) et en limons (2-2.5 %) sont très basses (Nzila, 1996). Ces sols sont grisâtres en surface et ocres en profondeur avec une structure particulaire.
Ils sont en outre caractérisés par une pauvreté chimique se manifestant par des teneurs en cations échangeables et une capacité d’échange cationique faibles (Laclau, 2001).
3.2MATERIEL VEGETAL
Trois clones différents ont été étudiés : le PF1 1-41, le 18-50 et le 18-52. Le clone PF1 1-41 (noté par la suite 1-41 dans la présente) provient d’un croisement naturel entre deux ou trois individus d’Eucalyptus alba congolais (Reinw. ex Blume, arbre femelle) et d’un groupe d’hybrides mal identifiés venant d’un arboretum brésilien (arbre mâle, Delwaulle 1988). Le 1-41, qui a pour qualité essentielle la robustesse, a été largement planté sur le massif (plus de 7000 hectares) et l’essentiel des calibrations du modèle de croissance ont été effectuées sur ce clone.
Les 18-50 et 18-52 sont des clones plein frères d’hybride E.urophylla*E.grandis. Ils ont été sélectionnés pour remplacer au niveau industriel l’hybride 1-41, moins productif.
3.3QUANTIFICATION DES CHUTES
3.3.1 Caractéristiques des parcelles
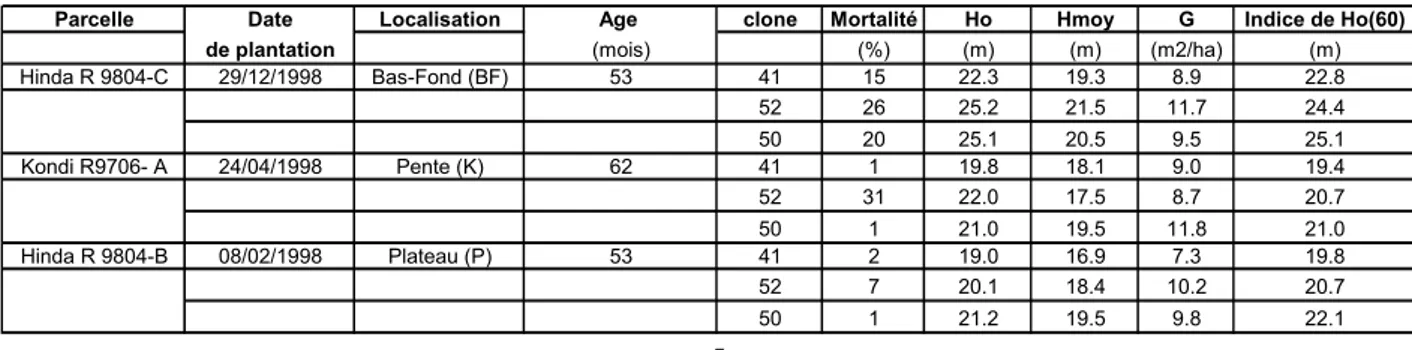
Des données provenant de 13 parcelles ont été synthétisées pour déterminer l’effet de l’âge sur les chutes de litières annuelles. Parmi ces essais, on trouve deux chronoséquences et quatre « site-atelier » suivis sur le long terme. Les caractéristiques de ces parcelles sont regroupées dans le tableau 1. On peut voir les périodes de suivi des parcelles sur la figure 6.
Pour la modélisation des chutes mensuelles trois des sites parmi les 13 satisfaisaient aux conditions énoncées dans l’introduction. Il s’agit des parcelles R00-13B, H98-07 et R92-80 sur lesquelles seul le 1-41 est planté. Le modèle de chute mensuel a donc été développé pour ce clone uniquement.
Tableau 1 . Caractéristiques des parcelles pour lesquelles des données de chute sont disponibles.
Parcelles Projet Date de plantation Fréquence Clones
R00-13 A Cycle jeune futaie mai-01 mensuel 18-52
R00-13 B Cycle jeune futaie mai-01 mensuel 1-41
R90-07 Chrono conv 98 décembre-89 mensuel 18-50
R96-03 Chrono conv 98 + Chrono JPL avril-97 mensuel 18-50
R92-04 Chrono conv 98 mars-93 mensuel 18-50
R95-07 Chrono conv 98 + Chrono JPL mars-96 mensuel 18-50
R89-06 C Chrono JPL novembre-89 mensuel 1-41
R92-05 Chrono JPL avril-93 mensuel 1-41
R92-05 C Chrono JPL avril-93 mensuel 1-41
R92-80 S Chrono JPL janvier-92 mensuel 1-41 et 18-52
R92-80 Cycle vieille futaie janvier-92 mensuel 1-41
H98-07 Site atelier carbone octobre-98 hebdomadaire 1-41
Cifor Cifor mars-98 hebdomadaire 1-41
3.3.2 Méthode de récolte des litières
Les litières ont été collectées dans des bacs à litière de 0.65m*0.65m ou 0.75m*0.75m. Pour toutes les parcelles, les différentes composantes ont été séparées à la main (feuilles, branches, écorces, divers) selon un protocole identique dont on trouvera un descriptif dans Laclau (2001).
3.3.3 Données disponibles
En ce qui concerne les données dendrométriques, seule la surface terrière (G) a été mesurée de façon systématique (mesures mensuelles) pour les sites sélectionnés. C’est cette variable qui a été choisie pour l’élaboration des modèles.
Le climat de Pointe-Noire est caractérisé par l’alternance d’une saison sèche qui va de juin à septembre et d’une saison humide qui va de octobre à mai (90% de la pluviométrie annuelle). Pour décrire les variations saisonnières, le paramètre le plus immédiat est le niveau de pluviométrie et une première solution pour distinguer les deux saisons consiste à déclarer un seuil critique de pluie en dessous duquel on considère que l’on est en saison sèche. Cependant ce niveau de pluviométrie n’est pas très bien adapté car selon les années, le seuil peut varier fortement.
Nous avons donc choisi d’utiliser, en plus de la pluviométrie, l’EvapoTranspiration Potentielle (ETP) qui intègre plusieurs variables climatiques. L’ETP de Penman-Monteith a été calculée sur la base de données disponibles pour le site atelier Carbone (parcelle H9807). Comme le jeu de données présentait des lacunes (mois entiers absents) l’ETP Turc a été calculée en parallèle. Ce modèle utilise des données climatiques réduites, disponibles à l’aéroport de Pointe-Noire : la température moyenne, le rayonnement global et l’humidité relative de l’air. Une régression été faite entre les deux ETP (R²=0.94) et à partir des coefficients, l’ETP Penman-Monteith a pu être prédite pour combler les données manquantes. Les courbes d’ETP obtenues ont ensuite été lissées à l’aide d’un filtrage FFT (Fast Fourrier Transform) pour dégager les grandes tendances saisonnières. Par la suite, nous avons plus précisément utilisé la dérivée de l’ETP comme variable indicatrice de la saison, alliée avec le seuil de pluviométrie.
3.3.4 Homogénéisation
Un travail de recalage de ces données (chute de litière, accroissements de surface terrière, ETP) a été effectué pour les ramener à un pas de temps mensuel. A titre d’exemple on calcule d’abord une quantité de chute journalière :
chute[ journalière entre t0 et t1] = chute[ à t1 ]/(t1-t0).
En additionnant les chutes journalières sur un mois donné, on obtient la chute mensuelle pour ce mois. La même opération a été effectuée pour recaler les accroissements en surface terrière et les données ETP.
Les chutes annuelles ont été calculées en sommant les chutes mensuelles sur une année, l’âge affecté étant celui du douzième mois. L’âge est connu sans ambiguïté (dates de plantation et d’inventaire connues)
3.4QUALITE DE LA LITIERE
3.4.1 Caractéristiques des parcelles
Pour l’expérience de décomposition en laboratoire, un plan factoriel complet a été élaboré. Les prélèvements ont été effectués sur trois sites de fertilités différentes où chacun des trois clones était planté. Les caractéristiques de ces sites sont regroupées dans le tableau 2.
Tableau 2. Caractéristiques des sites sélectionnés. Ho est la hauteur dominante, Hmoy la hauteur moyenne.
Parcelle Date Localisation Age clone Mortalité Ho Hmoy G Indice de Ho(60) de plantation (mois) (%) (m) (m) (m2/ha) (m)
Hinda R 9804-C 29/12/1998 Bas-Fond (BF) 53 41 15 22.3 19.3 8.9 22.8 52 26 25.2 21.5 11.7 24.4 50 20 25.1 20.5 9.5 25.1 Kondi R9706- A 24/04/1998 Pente (K) 62 41 1 19.8 18.1 9.0 19.4 52 31 22.0 17.5 8.7 20.7 50 1 21.0 19.5 11.8 21.0 Hinda R 9804-B 08/02/1998 Plateau (P) 53 41 2 19.0 16.9 7.3 19.8 52 7 20.1 18.4 10.2 20.7 50 1 21.2 19.5 9.8 22.1
Sur chaque site, l’unité expérimentale est une parcelle monoclonale de 12*4 plants et l’écartement entre les plants est de 4,70m *2,65m (densité de 800 t/ha qui correspond aux densités de plantation industrielles actuelles). Les prélèvements ont été faits sur les 20 arbres centraux.
Les valeurs de hauteurs dominantes varient entre 19 et 25.2 m à 50-60 mois. Ce sont des valeurs conformes à celles observées sur le massif. La comparaison directe entre les sites et les clones est difficile compte tenu de la forte mortalité observée pour le « bas-fond » et pour le clone 18-52. Nous avons donc utilisé l’indice de hauteur dominante des peuplements à 60 mois, pour une densité fixe de 800 tiges/ha qui est calculé à l’aide du dernier inventaire disponible pour chaque site et d’un modèle de croissance en hauteur dominante qui tient compte de la mortalité (Saint-André et al. 2002). Cet indice de performance potentielle en hauteur dominante permet de comparer les clones entre eux en s’affranchissant de l’âge et des variations de densité.
Sur la base de cet indice les trois stations sont classées comme suit : BF (IFmoyen = 24.1m) >> P (IFmoyen = 20.8m) ≈ K (IFmoyen = 20.4m),
Et pour chaque site, les clones se classent dans cet ordre 18-50 > 18-52 > 1-41.
La surface terrière qui dépend à la fois de la potentialité du clone sur le site et de la densité ne suit pas ce classement. Pour les localisations « bas-fond » et « plateau », sur lesquelles les clones ont à peu près le même taux de mortalité, le 52 présente une surface terrière supérieure au 18-50 (environ 4% d’écart sur le plateau et 20% d’écart dans le bas-fond) pour une hauteur dominante équivalente. Ce clone est donc le plus productif des trois mais semble plus sensible que les autres aux aléas. Le clone le moins productif est le 1-41.
3.4.2 Méthode de récolte des feuilles
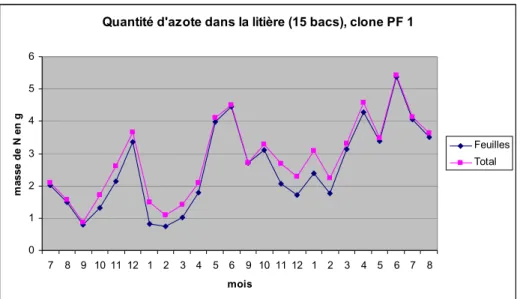
Dans le contexte de Pointe-noire, le facteur limitant majeur de la croissance est l’azote (outre le déficit hydrique en saison sèche). Or les feuilles fournissent environ 90% de l’azote de la litière totale (Laclau 2001).
Du fait de la petite taille des essais et du vent, le risque de pollution d’un bac à litière par les clones voisins est important. L’utilisation classique de bacs à litière a donc été exclue. Les feuilles ont été récoltées selon le protocole suivant : deux expérimentateurs travaillent en tandem pour récupérer les feuilles jaunes qui sont sur le point de tomber. L’un grimpe à l’aide d’échelles à mi-hauteur des arbres et les secoue afin de faire tomber les feuilles que l’autre ramasse. On a par ailleurs pu vérifier avec cette méthode que la dispersion des feuilles par le vent au moment de la chute se fait de façon isotrope et dans un rayon du même ordre que la hauteur de l’arbre.
La méthode employée permet de s’affranchir de ce doute sur la provenance de la litière puisque toutes ces feuilles ont été suivies à l’œil.
Les feuilles ainsi récoltées ont été mises à sécher à 23°C sous climatisation jusqu’à stabilisation de la masse. Sur la figure 8, le clone 18-52 est celui qui a perdu le moins d’eau (comparaison par station).
Des échantillons de sol de l’horizon [0-25 cm] ont également été prélevés sur l’ensemble des trois sites afin d’inoculer les substrats des microcosmes avec la microfaune locale. L’ensemble de ces échantillons de sol a été mélangé afin d’obtenir les conditions de décomposition les plus homogènes possibles entre microcosmes (substrats, température et humidité identique).
3.4.3 Décomposition en laboratoire
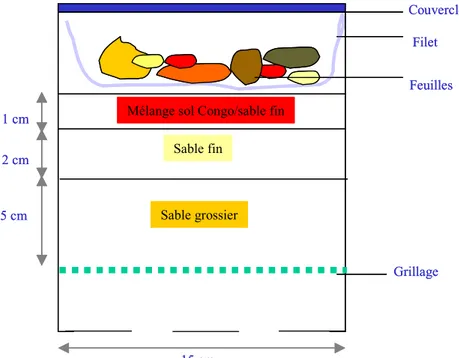
Le protocole utilisé reprend les modalités décrites dans Joffre et Agren (2001). La décomposition a été étudiée sur une période de 6 semaines avec 3 répétitions par couple site/clone et par semaine. Cent soixante deux microcosmes (6*3*9) ont ainsi été préparés comme indiqué sur la figure 9 avec des pots en PVC. Le fond des pots en PVC était percé de plusieurs orifices afin de reproduire la
forte capacité de drainage observée in situ (Laclau 2001). Trois grammes environ de feuilles fraîches ont été pesés pour chacun des 162 échantillons et déposés à la surface des pots sur un filet d’étamine de maille 1mm, afin de permettre la progression des décomposeurs vers les feuilles. Cette quantité de matière correspond dans les conditions contrôlées du microcosme à un recouvrement de la surface sans effet d’accumulation (optimisation de l’interface sol/feuille). Afin d’humidifier les feuilles de litière, elles ont été placées à lessiver pendant 72 heures dans 150 ml d’eau déminéralisée avant d’être placées dans les microcosmes. Cette quantité d’eau correspond à la capacité au champ du substrat utilisé. Pour la déterminer, une quantité d’eau suffisamment importante pour dépasser la saturation (200ml) a été versée dans un microcosme et la quantité d’eau écoulée en surplus mesurée. La différence de ces deux quantités donne la capacité au champ. La solution de lessiva riche en éléments hydrosolubles a été ensuite versée sur les feuilles afin de conserver les nutriments nécessaires à l’activité des décomposeurs (microflore et microfaune). Les pots ont été placés dans une chambre d’incubation maintenue à 22°C à l’obscurité. Une fois par semaine une série de 27 pots (9 couples clone/site avec 3 répétitions) était retirée. A la même date, les pots restants dans la chambre ont été ramenés au taux d’humidité de départ grâce à un ajout d’eau déminéralisée. L’humidité des microcosmes a donc été maintenue à la capacité au champ. Les feuilles décomposées ont été placées 48 heures à l’étuve à 55°C puis pesées et broyées (sauf pour la 4ème semaine de décomposition où le temps de passage à l’étuve a été de 96 heures). Le broyage a été effectué avec une centrifugeuse (Cyclotec®1093, filtre à maille de 1mm) pour éviter de perdre des composés chimiques par dégradation thermique.
Le sol du Congo étant un sable siliceux, on a utilisé pour le mélange un sable de rivière dépourvu de composés carbonatés et un gravier siliceux pour le sable grossier. Un test négatif au HCl à 30% a été fait sur ces deux sables (absence d’effervescence)
3.4.4 Analyses spectrophotométriques
Les poudres obtenues après broyage ont été analysées par spectroscopie en proche infra-rouge (NIRS) dans les laboratoires de l’UMR CEFE à Montpellier. Il s’agit d’une méthode d’analyse physique basée sur l’absorption sélective des radiations électromagnétiques de certaines longueurs d’onde situées dans le domaine du proche infra-rouge (1100 – 2500 nm) par les molécules organiques qui contiennent des liaisons entre atomes de carbone, hydrogène, oxygène et azote. L’absorbance est ainsi caractérisée par l’agencement des liaisons atomiques qui détermine lui-même la composition chimique du matériau étudié.
En pratique les poudres sont placées dans une cellule de mesure. Celle-ci est constituée d’une cuvette cylindrique avec un fond transparent en quartz et un couvercle en carton qui permet de compresser la litière broyée contre le quartz. Ces échantillons sont ensuite passés au spectrophotomètre (NIRSystem 6500) où ils sont illuminés par une source de radiations monochromatiques dont la longueur d’onde varie de 400 à 2500 nm avec une illumination tous les 2 nm. La quantité de radiations lumineuses réfléchie par la surface de l’échantillon aux différentes longueurs d’onde est mesurée et l’ensemble de ces mesures permet d’obtenir un spectre d’absorbance (pas 2 nm)
Cette information spectrale a été traitée par diverses méthodes d’analyse statistique afin d’évaluer certains paramètres notamment la vitesse de décomposition v, le rapport C:N et le facteur de qualité qo.
Les mesures de masses restantes ont été effectuées en corrigeant d’abord par rapport au temps de passage à l’étuve. Les masses initiales (MFo) sont en effet des masses fraîches alors que chaque échantillon de feuilles décomposées correspond à une masse sèche (MSt).
MSo=MFo*TMSr
où Mso est la masse sèche initiale et TMSr la teneur en matière sèche relative au temps de passage à l’étuve (Figure10). Elle a été calculée en utilisant deux échantillons de 3.5 grammes de feuilles
par couple clone/site (soit 18 répétitions) suivis pendant un mois et pesés aux dates 0, 2, 4, 7 et 30 jours.
La masse restante à t (MRt) est calculée comme suit : MRt=MSt/MSo
De plus une correction a du être faite en tenant compte des taux de cendre : en effet pendant le processus de décomposition, du sable se retrouve sur les litières décomposées et perturbe les mesures de masse restante. Pour s’affranchir de ce biais, 50 échantillons représentatifs des 6 semaines de décomposition ont été choisis à partir des données spectrales. Deux cent milligrammes de poudre ont été prélevés, pesés et brûlés à 650 degrés pendant 3 heures. Dans le résidu après brûlage, il reste les éléments minéraux des feuilles et le sable. Le rapport des masses avant et après brûlage donne un taux de cendre (TCt). La même manipulation a été effectuée avec des feuilles de litière initiale (non décomposée et non lessivée) afin d’obtenir un taux de cendre initial (TCo). L’information contenue dans les spectres a ensuite été utilisée afin d’évaluer les taux de cendre de chaque échantillon. Au pas 2 nm les valeurs des spectres entre deux longueurs d’onde successives sont très corrélées. Une méthode d’analyse statistique pour des valeurs de régresseurs fortement corrélées est la PLS (Partial Least Square Regression). Cette méthode consiste à faire une analyse en composante principale en tenant compte d’une variable supplémentaire comme variable indépendante. Comme on a cherché à décrire un phénomène de décomposition c’est la masse restante qui a été choisie. A partir des axes factoriels obtenus, une équation de calibration a été établie pour prédire les taux de cendres pour chacun des 162 échantillons. En corrigeant les masses mesurées avec ces taux de cendres on obtient les masses de matière organique restante.
La masse restante à t corrigée par les cendres vaut donc :
MRtcorr = MRt * (100-TCt)/(100-TCo)
C’est cette variable qui est utilisée par la suite dans les analyses statistiques pour évaluer la qualité initiale de la litière.
3.4.5 Analyses chimiques
Pour quantifier le carbone et l’azote contenus dans les feuilles, des mesures en micropesée ont été effectuées. Un broyage, supplémentaire à celui effectuée pour le NIRS, a été réalisé à l’aide d’un broyeur à bille. En effet, les analyses faites au CHN demandent d’avoir un matériel le plus fin et homogène possible en raison de la taille des capsules utilisées : le risque de tomber sur un échantillon de 2mg qui contienne une concentration différente de l’ensemble (2 g) est d’autant plus élevé que le broyage est hétérogène. Des standards spécifiques au laboratoire sont par ailleurs utilisés pour vérifier que les mesures ne dérivent pas dans le temps.
3.4.6 Analyses physiques
3.4.6.1 Epaisseur des feuilles (e)
Elle a été mesurée avec un appareil constitué d’un noyau de fer doux accroché à un ressort et placé dans le champs d’un bobine où circule du courant. Un millivoltmètre mesure la tension aux bornes de la bobine, proportionnelle au déplacement du noyau. Il existe une relation linéaire très forte entre l’épaisseur de la feuille (égale au déplacement du noyau) et la différence de tension. L’appareil a été recalibré toutes les 50 mesures environ. La sensibilité de l’appareil suppose que l’on utilise des parties végétales les plus plates possible. Le gondolement résultant du séchage des feuilles est en effet du même ordre de grandeur que l’épaisseur et si l’on ne s’assure pas que la portion de feuille est plate la mesure perd toute signification. Sur chaque feuille, un rectangle de 1cm sur 5 mm environ a été découpé sur le limbe et l’épaisseur mesurée avec trois répétitions sur cette portion de la feuille. Six feuilles ont été utilisées par couple clone/site (soit un total de 9*6*3=162 mesures)
3.4.6.2 Masse surfacique (LMA pour leaf mass area)
Elle est mesuré en faisant le rapport de la masse à la surface d’un échantillon (un dizaine de feuilles environ). Quatre répétitions par couple clone /site ont été effectuées (soit un total de 36 mesures).
3.4.6.3 Densité
Elle a été obtenue en faisant le rapport du LMA moyen à l’épaisseur moyenne : d=LMA/e.
4. RESULTATS
4.1MODELISATION DES CHUTES DE LITIERES
4.1.1 Analyse des chutes annuelles, effet de l’âge des arbres
La figure 11 montre l’effet de l’âge sur les quantités de litière qui tombent à l’hectare sur une année. Une observation globale permet de distinguer une première phase croissante suivie d’un plafonnement.
On a effectué une régression exponentielle de type (l’âge est donné en mois) :
Chute=A*(1-exp(-B*age²)) (eq1)
Le paramètre A correspond à un niveau de production moyen qui est constant à l’âge adulte (plafonnement) et le paramètre B à une performance de croissance (vitesse avec laquelle les chutes atteignent leur asymptote).
Pour un modèle où A et B sont communs à tous les clones (modèle à deux paramètres) on trouve un R² de 0.75. Pour un modèle où chaque paramètre est spécifique au clone (modèle à 4 paramètres, la donnée du 18-52 étant assimilée à du 18-50) on trouve un R² de 0.76 mais un test F réalisé sur la somme des carrés des écarts (Brown and Rothery 1994) montre que le gain en R² ne justifie pas l’ajout de deux paramètres. Ceci montre qu’il n’y a pas, pour ce modèle, de différences significatives entre les clones. Le modèle global a donc été choisi avec A=6.18 (t.ha-1.an-1) et B=0.0013 (mois-2)
Un modèle segmenté a ensuite été testé pour décrire l’âge critique de transition entre les deux phases de chutes.
Une régression linéaire bi modale de type :
Chute= B1*age pour age<AgeCritique (eq 2) Chute= AgeCritique(B1-B2)+ B2*age pour age>AgeCritique
donne un R² de 0.85, avec B1=0.15 t/ha/mois² et B2=0.025t/ha/mois².
Elle permet de montrer l’existence d’un seuil au niveau du fonctionnement des chutes : une première phase linéaire croissante jusqu’à un âge critique suivie d’un plafonnement (figure 12). Cet âge critique peut être relié à la fermeture du couvert à partir de laquelle le houppier cesse de grandir et les chutes potentielles de litière se stabilisent autour d’une moyenne (environ 6t/ha/an). L’observation des résidus en fonction de l’âge montre que la variabilité résiduelle s’exprime fortement pour les peuplements ayant atteint cet âge critique (environ 30 mois). Cependant les données disponibles étaient trop peu nombreuses aux jeunes âges pour vérifier cette hypothèse d’hétéroscédasticité.
4.1.2 Analyse des chutes mensuelles, effet du climat et de la croissance
La variabilité du modèle annuel reste forte (25% des chutes moyennes annuelles, soit 1.5 t.ha-1.an-1). Pour pouvoir l’expliquer et se conformer au pas de temps du modèle de croissance (mensuel), les chutes mensuelles ont été étudiées pour les trois sites ateliers décrits dans le matériel et méthode (H9807, R0013B, R9280, clone 1-41 uniquement).
4.1.2.1 Analyse générale
L’observation de la figure 13 permet de distinguer de façon grossière deux pics de chutes dans l’année, un au début de la saison humide et l’autre à la saison sèche.
La figure 14 donne les variations du taux de feuille dans la litière, en fonction de l’âge et différencié pour les deux saisons. Les feuilles représentent 98% de la litière jusqu’à deux ans. En saison sèche les feuilles constituent au minimum 75% de la litière. A partir de 24 mois les autres composantes (branches mortes, écorce, divers) peuvent tomber de façon prépondérante à la saison humide. Cela permet aussi de constater que les pics de chute en saison sèche et humide sont de natures différentes : il tombe essentiellement des feuilles en saison sèche car le niveau hydrique du sol baisse et la dessiccation du pétiole provoque la chute. En revanche il tombe beaucoup plus de composés divers en saison humide car le niveau d’agitation mécanique dû au vent et à la pluie est important. On a donc fait l’hypothèse dans le modèle mensuel qu’il existe une bi-modalité des chutes de litière; le premier pic correspond à un phénomène physiologique (dessiccation, baisse du régime hydrique) alors que le deuxième est plutôt dû à des phénomènes mécaniques.
4.1.2.2 Modèle pour les chutes de feuilles
Pour caractériser l’alternance des saisons, la dérivée de l’ETP a été examinée en faisant l’hypothèse que le début de la saison sèche correspond à une chute brutale de l’ETP tandis qu’une augmentation brutale correspond au début de la saison humide.
La figure 15 montre que chaque pic de chute de litière foliaire correspond à un extremum de la dérivée de l’ETP. Si c’est un maximum, il souligne l’augmentation du degré d’agitation extérieure (vents forts, pluies violentes) en correspondance avec le pic de saison humide. Si c’est un minimum il correspond à une chute du régime hydrique qui accompagne le début de la saison sèche et le pic de chute de litière foliaire associé.
La forme du modèle choisi s’appuie donc sur cette variable. On a effectué une régression linéaire des chutes de feuilles avec la dérivée de l’ETP de façon bi-modale : une pente positive pour les cas où la dérivée de l’ETP est positive, une pente négative sinon.
L’effet de l’âge a été pris en compte dans la pente du modèle. On obtient le résultat suivant :
Chute= a + b*d/dt(ETP) (eq3)
Où b=bo*(1-exp(-lambda*âge²)) bo>0 si d/dt(ETP)>0
bo<0 si d/dt(ETP)<0
On exprime ainsi 65% de la variabilité, avec une bonne prise en compte de l’effet âge (figure 16). En examinant les résidus on se rend compte qu’il existe des points qui sortent largement du modèle et qui correspondent à certains pics importants de chutes. En effet si la dérivée de l’ETP décrit bien le phénomène d’ensemble et la rythmicité des chutes, elle ne tient pas compte de la biomasse sur pied qui détermine indirectement la quantité de litière potentielle.
En ce qui concerne le pic de chute foliaire en saison sèche, les feuilles qui tombent ont été produites six mois avant pendant la saison humide (Laclau 2001). Or l’arrivée de ces feuilles permet d’élaborer des accroissements en surface terrière observés trois mois après (Fabre 2003). On a donc fait l’hypothèse qu’une chute au temps t était corrélée au ∆G à t-3 (figure 17).
De plus les feuilles qui tombent en début de saison humide ont été produites à la fin de la saison humide précédente. Ces feuilles permettent probablement à l’arbre de se maintenir pendant la saison sèche et si les conditions hydriques ne sont pas trop restrictives, de faire des réserves. A la saison humide suivante, ces feuilles tombent et les réserves de l’arbre accumulées permettent d’exprimer un nouveau potentiel de croissance en diamètre qui survient trois mois après le début de saison humide. On a donc fait l’hypothèse que le pic de début de saison humide était corrélé avec les accroissements de surface terrière qui surviennent trois mois après. (Figure 18)
Enfin les pics les plus forts et ceux qui sont le moins pris en compte par le modèle sont les pics qui surviennent en mai, qui marque chaque année, la transition entre les deux saisons. On a souligné cette particularité dans le modèle en affectant un coefficient particulier au mois qui permet le passage d’une saison à l’autre.
Le modèle final adopté est donc sous la forme :
Chute(t)= h*(a+b* d/dt(ETP)) (eq4)
Avec comme paramètres :
b=bo*(c-exp(-lambda*âge²)) avec bo>0 si d/dt(ETP)>0 bo<0 si d/dt(ETP)<0
h= 1 pour les jeunes âges et pour ∆G < seuilG r1*∆G(t+3) si d/dt(ETP)>0
r1*∆G(t-3) au mois de mai r2*∆G(t-3) si d/dt(ETP)<0
Environ 80% de la variabilité des chutes mensuelles est ainsi expliquée par le modèle. Le RMSE est de 88 kg soit 25% de la moyenne des chutes mensuelles (341 kg/ha).
4.1.2.3 Chutes mensuelles subéro-ligneuses
La chute de branches et d’écorces est caractérisée essentiellement par un pic en début de saison humide. L’hypothèse faite pour les chutes de feuilles en début de saison humide a été conservée ici puisque le ∆G à t+3 décrit le même phénomène mécanique.
Le modèle élaboré est de la forme :
Chute(t)= b* ∆G(t+3) (eq 5) Où
b= ETP*bo*(1-exp(-lambda*âge²)) si d/dt(ETP>0) et ∆G(t+3)>seuil b= bo*(1-exp(-lambda*âge²)) sinon
4.2QUALITE DES LITIERES EN FONCTION DE LA PROVENANCE ET LA FERTILITE
4.2.1 Analyse des propriétés des feuilles avant décomposition
Une ANOVA (analyse de variance) sur l’épaisseur et le LMA a été faite, en utilisant le critère de Scheffe comme discriminateur. Il ressort que le clone 1-41 a des feuilles plus épaisses et un LMA plus élevé que les deux clones plein frères. Il n’y a pas de différence significative entre le 18-50 et le 18-52, ni pour l’épaisseur ni pour le LMA.
Le LMA est par ailleurs significativement supérieur sur le plateau (station P). Les deux autres stations n’affichent pas de différences significatives.
Enfin les feuilles sont significativement moins épaisses sur la pente (station K) que sur les deux autres stations qui ne présentent pas de différences significatives.
Tableau 3 :Valeurs moyennes pour le LMA, l’épaisseur et la densité.
Modalité BF41 P41 BF50 K41 K50 P50 P52 BF52 K52 Densité 6.40 7.09 6.47 6.67 6.39 7.31 6.47 6.39 6.96 Epaisseur (mm) 0.1923 0.1936 0.1655 0.1702 0.1645 0.162 0.1751 0.1722 0.1617 LMA (g/m²) 122.99 137.28 107.04 113.49 105.12 118.45 113.26 110 112.59 4.2.2 Décomposition en microcosme 4.2.2.1 Analyse spectrophotométrique
Une analyse en composantes principales classique a été faite sur toutes les longueurs d’ondes comme variables indépendantes, et la provenance génétique, la station, la teneur en azote prédite et la masse restante prédite comme variables dépendantes. Cette analyse permet d’observer comment se distinguent ces paramètres à partir des seuls spectres.
Tableau 4 : Contribution des axes au nuage global (pourcentage cumulé)
Axes 1 2 3 4 5 6 7 8 9
% Variance 44 71.9 78.9 85.2 90.2 93 94.7 95.9 96.8
Les contributions des axes pour chaque variable sont les suivantes: Clones axes 4, 3 et 1 ; 94% de la variance expliquée. RM axes 1, 2 et 3 ; 79% de la variance expliquée. Azote axes 3, 1 et 17 ; 72% de la variance expliquée. Station axes 9, 2 et 3 ; 25% de la variance expliquée.
La paramètre le mieux expliqué est donc la provenance génétique. Le clone 1-41 se différencie en terme de composition chimique des deux autres qui sont plein frères.
Les teneur en azote sont elles aussi bien expliquées par les spectres puisque le modèle exprime 72% de la variance sur ces teneurs.
4.2.2.2 Profils de décomposition et modélisation
La figure 21 montre le profil de décomposition pour les neuf modalités étudiées. La perte moyenne de masse en six semaines est de 25%. La perte la plus importante se fait pendant la première semaine.
Il a été utilisé afin d’obtenir un classement des clones et des fertilités en fonction de la vitesse de décomposition. Il s’agit d’une régression exponentielle simple décroissante sur les masses restantes (MRtcorr) qui peut s’écrire sous la forme :
MRtcorr =Qolson*(exp(-v*t)) (eq5)
Où
Qolson représente la quantité de matière au stade initial, t le temps de décomposition,
v la vitesse de décomposition.
Ce modèle a été testé sous plusieurs modalités en faisant varier le nombre de paramètres (p) et en utilisant la procédure Nlin de SAS.
Option 1. Qolson et v communs pour l’ensemble des données (p=2 ) Option 2. Qolson et v spécifiques pour chaque clone (p=6)
Option 3. Qolson et v spécifiques pour chaque station (p=6)
Option 4. Qolson et v spécifiques pour chaque modalité (clone*station) (p=18) Les valeurs de v et Qolson sont données dans le tableau 5.
Tableau 5 : valeur des paramètres v et Qolson obtenus lors de l’ajustement du modèle d’Olson (1963)
vitesse v Option 1-41 18-50 18-52 1-41 18-50 18-52 1-41 18-50 18-52 1 2 0.020 0.016 0.026 0.020 0.016 0.026 0.020 0.016 0.026 3 4 0.016 0.012 0.014 0.034 0.015 0.037 0.011 0.020 0.027 Qolson Option 1-41 18-50 18-52 1-41 18-50 18-52 1-41 18-50 18-52 1 2 0.850 0.840 0.820 0.850 0.840 0.820 0.850 0.840 0.820 3 4 0.872 0.855 0.823 0.869 0.841 0.828 0.818 0.840 0.806 BF K P BF K P 0.020 0.019 0.029 0.014 0.840 0.850 0.850 0.820
On remarquera que le Qolson représente en fait le pourcentage de masse restante après lessivage : c’est en effet la quantité de matière initiale extrapolée à partir des différents stades de décomposition, donc la masse réelle à laquelle les décomposeurs se sont attaqués. On trouve par ailleurs des valeurs qui concordent avec celles trouvées dans la littérature (de 13 à 20% de perte de masse par lessivage)
Entre chaque option, la somme des carrés des écarts (SSE) a été testée en fonction du nombre de paramètres à l’aide d’un test-F (Brown and Rothery 1994). On a obtenu le classement suivant par ordre décroissant de significativité des modèles :
SSE RMSE Nbparam R²
modalité 0.146 0.029 18 0.709
clone 0.240 0.036 6 0.521
site 0.260 0.038 6 0.481
total 0.340 0.043 2 0.323
R²=1-SSE/(somme des carrés des écarts à la moyenne)
Le meilleur modèle est celui avec 18 paramètres et le moins bon est le modèle global à deux paramètres. Ceci indique que chaque modalité est significativement différente des autres. L’effet
clone et l’effet station sont eux aussi significatifs. De plus l’effet clone est légèrement plus fort que l’effet station puisqu’on obtient une SSE inférieure pour le même nombre de paramètres.
Compte tenu des valeurs observées des paramètres, nous avons testé l’hypothèse où Qolson et v sont communs pour les clones 1-41 et 18-50 et spécifiques pour le clone 18-52 (p=4). Le même test-F a été utilisé.
Le clone 18-52 décompose plus vite que les deux autres (le modèle global à 2 paramètres est moins bon que le modèle «clones groupés»). Le 18-50 et le 1-41 ne sont pas significativement différents (modèle avec clones groupés meilleur que modèle clone).
Une hypothèse similaire a été faite en observant l’effet de la station (Station à Kondi par rapport aux deux stations à Hinda).
SSE RMSE Nbparam R²
site 0.260 0.038 6 0.476
site groupés 0.313 0.041 4 0.370
total 0.340 0.043 2 0.315
La vitesse de décomposition est effectivement plus grande pour la station K que pour les deux autres (comparaison entre le modèle «sites groupés» et le modèle global) mais les deux stations à Hinda ne peuvent pas être regroupées (site/groupé). Les trois stations sont donc significativement différentes les unes des autres.
En considérant cette analyse réalisée avec le modèle d’Olson il y a donc bien un effet de la station et du clone sur la qualité de la litière foliaire : le clone 18-52 décompose mieux que les autres (tous sites confondus) et la décomposition est plus rapide sur le site de Kondi que pour les deux autres (tous clones confondus). La vitesse de décomposition la plus basse est obtenue dans le bas-fond mais l’effet n’est pas significatif.
Modèle d’Agren(1996)
Un autre critère permettant de décrire la qualité de la litière est le paramètre q0 dans l’équation de décomposition développé par Agren et Bossata (1996) qui s’appuie sur l’activité des décomposeurs dans le substrat.
L’équation s’écrit
MRtcorr = (1+fc*n11*beta*u0*(q0**(beta))*jours)**(-(1-e0)/(n11*beta*e0)) Où :
fc est la concentration de carbone dans la masse des décomposeurs
n11 décrit la distribution du carbone selon la qualité des molécules carbonées beta est un facteur de forme
uo est le taux de croissance des décomposeurs par unité de carbone
eo est le rapport du carbone produit au carbone assimilé par les décomposeurs jours est le temps de décomposition en microcosme
SSE RMSE Nbparam R²
clone 0.240 0.036 6 0.516
clones groupés 0.242 0.036 4 0.513
Ce modèle a été testé avec la procédure Nlin de SAS en laissant libre beta et q0 et en gardant des valeurs communes pour tous les autres paramètres. En effet ces paramètres sont tous liés au substrat de décomposition qui est identique pour tous les microcosmes et seul q0 est un facteur caractéristique de la litière en décomposition. Les valeurs de ces paramètres ont été définies dans Agren (1996), avec fc=0.5, eo=0.28, n11=0.36 et uo=0.0038.
Le classement des modalités selon q0 est donné par le tableau 6.
Une validation croisée a été testée pour ce modèle en fixant le qo à 1 pour une des modalités et en laissant libres les huit autres. On obtient le même classement dans tous les cas. La modalité bas-fond/1-41 (notée BF41) étant celle où qo est le plus bas les résultats sont présentés ici pour qo(BF41)=1
Tableau 6 : Valeur des paramètres q0 obtenus lors des ajustement du modèle d’Agren et Bossata (1996)
station clone modalité qo Std
bf 41 BF41 1 bf 50 BF50 1.0051 0.0182 k 50 k50 1.0295 0.0178 p 41 p41 1.0357 0.0178 bf 52 BF52 1.0414 0.0177 p 50 p50 1.0444 0.0177 k 41 k41 1.0574 0.0178 p 52 p52 1.0898 0.0175 k 52 k52 1.0942 0.0175
Ici encore le clone 18-52 se distingue des clones 1-41 et 18-50 par une qualité de litière supérieure. La station BF est significativement différente des deux autres avec un qo plus faible donc une vitesse de décomposition plus lente.
5 DISCUSSION
5.1MODELISATION
5.1.1 Modèle de chutes annuelles
Le modèle de chutes annuelles reflète, dans le cas exponentiel, comme dans le cas segmenté, une hétéroscédasticité des données : plus le peuplement est âgé et plus la variabilité résiduelle est forte. Cette tendance avait déjà été observée pour d’autres essences (pour le Pin Patula par exemple, Malhotra 1987). Pendant les premiers stades de croissance c’est uniquement la quantité de biomasse foliaire qui détermine la quantité de litière. Il n’y a en effet pas de branches mortes ni d’écorces susceptibles de tomber à ce stade. La litière est essentiellement composée de feuilles et l’effort mécanique nécessaire pour les faire tomber n’est pas très important (voir la méthode de récolte pour l’étude de la qualité des litières). La chute de branches et d’écorce intervient lors de la fermeture du couvert (absence de lumière dans le sous étage, élagage naturel). Cette chute est beaucoup plus sensible aux facteurs environnementaux car elle nécessite un effort mécanique beaucoup plus important. Ces deux facteurs (composition de la litière et sensibilité mécanique) permettent sans doute d’expliquer la variabilité des chutes autour de la valeur de plafonnement.
5.1.2 Modèle de chutes foliaires mensuelles
5.1.2.1 Choix des paramètres
Le modèle mensuel segmenté avec la dérivée de l’ETP (équation 3, sans prise en compte du ∆G) a été développé de façon discontinue pour le passage de la saison sèche à la saison humide. On a choisi de laisser libre les interceptes bien que le modèle avec un intercepte commun ait plus de signification (l’investissement d’un paramètre n’est pas justifié par le gain en SSE). La raison de ce choix tient au fait que ce n’est pas le même effet que décrit la dérivée de l’ETP selon qu’elle est croissante ou décroissante. Dans un cas c’est un effet mécanique qu’elle traduit, dans l’autre un effet biologique lié au stress. Ceci justifie que les interceptes aient été laissés libres.
Pour ce qui est de la fonction de correction par l’âge, on aurait pu choisir d’intégrer le modèle segmenté puisqu’il donne un R² meilleur que le modèle exponentiel. Pourtant, l’existence d’un seuil dépendant des données considérées (le seuil est un paramètre du modèle dans l’équation 3) fait diverger le modèle général si on l’intègre avec ce modèle segmenté. Il doit donc exister un effet de la station et du clone sur ce seuil d’âge critique (moment de la fermeture du couvert), mais les données de chutes annuelles étaient trop peu nombreuses pour vérifier cette hypothèse. On a donc choisi le modèle exponentiel pour décrire l’effet de l’âge, que ce soit dans la correction de la pente ou bien dans la correction par le ∆G (fonction b et h dans l’équation 4).
Le choix du paramètre de seuil pour le ∆G (seuilG de l’équation 4) est pour l’instant fixé de façon arbitraire dans le travail présenté. Cela reste cependant une variable accessible en prenant par exemple le minimum du ∆G pour tous les mois de saison sèche.
5.1.2.2 Analyse des résidus
Les résidus ne présentent pas de corrélation avec les variables explicatives (âge, ∆G, d/dt(ETP)) et leur espérance est nulle. Leur variabilité reste importante (88 kg soit 25% des moyennes des chutes mensuelles) et peut être due à différents facteurs :
1/ l’exposition n’a pas été prise en compte dans les conditions stationnelles ;
2/ les données d’ETP sont globales et l’acquisition de données locales devrait encore améliorer les performances du modèle.
Cependant, si l’amplitude des résidus est importante au pas de temps mensuel, elle l’est beaucoup moins au pas de temps annuel du fait de la compensation des écarts d’un mois sur l’autre : la somme des chutes prédites mensuelles sur une année donne une valeur très proche des chutes annuelles réelles. La régression entre les valeurs prédites sommées à l’année et les valeurs effectivement mesurées donne un R² de 0.98 (pente 1.06 et intercepte 0.19 t/ha/an).
5.1.2.3 Perspectives
Le modèle ainsi développé présente plusieurs avantages :
1/ il s’appuie sur des variables climatiques intégratrices simples à mesurer (il faut la température, le rayonnement et l’humidité relative pour obtenir l’ETP Turc) indépendantes des caractéristiques stationnelles et du clone (l’ETP à partir duquel les données ont été traitées concerne l’ensemble du massif) ;
2/ la fertilité de la station est prise en compte par l’intermédiaire du ∆G. Des essais ont été effectués avec l’accroissement en hauteur dominante (sur la parcelle H9807), mais celui-ci dépendant également de la densité pour l’eucalyptus au Congo, il n’apporte pas plus d’informations que ∆G.
Il utilise donc des variables globales facilement accessibles et il sera possible de l’enrichir progressivement avec de nouvelles données.
La limite majeure de ce modèle de chute de litière mensuel (pour une intégration dans le modèle de croissance Eucalypt-Dendro) est qu’il utilise un ∆G mesuré mensuel incluant l’effet des saisons. Or, le modèle de croissance ne tient pas compte du climat pour estimer les différents accroissements. Il simule un ∆G qui ne peut servir d’entrée au modèle de chute.
Deux options peuvent être envisagées pour les études futures :
1/ une intégration du climat dans le modèle de croissance, et les premiers essais sont concluants sur les accroissements en hauteur dominante et en surface terrière (Fabre, 2003) ;
2/ obtenir une nouvelle formulation du modèle annuel par intégration analytique du modèle de chute de litière mensuel (au moins dans la phase où il n’y pas plus de variations liées à l’âge). L’idée étant d’obtenir un modèle annuel qui utilise comme données d’entrée un accroissement en surface terrière calculé cette fois-ci sur une base annuelle, et des variables climatiques basées sur l’ETP et ses variations intra-annuelles.
Enfin, la transposition à d’autre clones semble possible. Compte tenu du fait que le modèle de chute annuelles était invariant en fonction du clone, il est probable que le modèle mensuel reste identique dans sa formulation. La différence entre clones s’exprimera plutôt via les paramètres de pente, ou de seuils (répartition des chutes différentes dans l’année) mais au final, la somme des chutes annuelles seront équivalentes entre clones. Cette analyse est à tempérer compte tenu du déséquilibre des données entre les trois clones étudiés.
5.2QUALITE DE LA LITIERE FOLIAIRE
5.2.1 Protocole d’expérience
La méthode de récolte des feuilles peut être contestée. L’action de secouer les arbres provoque une perturbation mécanique et l’on n’est pas sûr que le processus de translocation des éléments minéraux des feuilles vers les branches avant la chute soit complet. Dans la littérature, la plupart des mesures effectuées sur des litières proviennent de matériel ayant passé un certain temps dans les bacs à litière où les feuilles sont plus ou moins lessivées (Malhotra 1987, Guo 1998, Trofymow 1991). Comme c’est la matière organique qui est prioritairement lessivée, les teneurs en azote peuvent être artificiellement augmentées. Ceci expliquerait pourquoi on trouve des teneurs en azote légèrement plus faibles pour des conditions semblables (7 pour mille dans la présente contre 9 à 10 pour mille dans Laclau 2001, clone 1-41 peuplements de 5 ans).
Pour le modèle de Agren on trouve des valeurs de bêta très fortes par rapport aux valeurs données par Agren et Bosatta (1996). Ceci est du à la courte durée de l’expérience de décomposition. On ne dispose pas de points placés suffisamment loin dans le temps pour pouvoir obtenir un modèle prédictif. Le facteur de forme est donc sans signification. En revanche les six premières semaines suffisent à déterminer la pente initiale du modèle qui contient qo et qui ne dépend pas de bêta. Le modèle a donc été conservé bien que les valeurs trouvées pour certains paramètres ne correspondent pas à la réalité.
5.2.2 Méthodes statistiques
Les dérivées secondes sont utilisées comme variables indépendantes à la place des spectres bruts parce qu’elles permettent de s’affranchir d’une différence d’intensité du signal qui pourrait être uniquement liée à la granulométrie de l’échantillon (plus le grain du broyage est fin et plus la quantité d’énergie absorbée par les poudres est grande. Une hétérogénéité au niveau du broyage aura donc une répercussion sur la hauteur des spectres, à laquelle la dérivée seconde est insensible). De plus la dérivée seconde caractérise les changements de concavité d’une fonction ; elle repère donc naturellement les pics d’absorption sur le spectre. Ceci correspond bien à l’information sur la composition chimique que l’on recherche.
Le choix du nombre de facteurs PLS se fait en minimisant simultanément la SEC (erreur standard de calibration) et la SECV (erreur standard de validation croisée) afin d’éviter le phénomène d’overfitting. La SECV est la moyenne des écarts-types (SE) calculée par quatre validations successives, c’est-à-dire que le modèle est calculé à partir de trois quarts des échantillons et validé sur le quart restant, et ceci à quatre reprises. La SECV est la moyenne des quatre SE. La SECV présente un minimum car si le nombre de paramètres devient trop grand vis-à-vis du degré de liberté des données le modèle perd sa significativité (overfitting). On choisit donc le nombre de facteurs explicatifs en prenant celui qui minimise la SECV (la SEC est décroissante en fonction du nombre de facteurs).
Tableau 8: Résultat de la PLS avec 6 facteurs pour les taux de cendres. SEC est l’erreur standard de calibration ; SECV est l’erreur standard de validation croisée ; 1-VR est le taux de variance expliquée par le
modèle ; SD est la déviation standard d’une série de données
NB facteurs SEC R² F SECV 1-VR
1 0.047 0.743 641.21 0.048 0.741 2 0.034 0.869 213.4 0.034 0.866 3 0.029 0.901 70.78 0.03 0.894 4 0.026 0.925 72.08 0.027 0.915 5 0.024 0.936 35.9 0.026 0.922 6 0.023 0.939 13 0.026 0.926
5.2.3 Comparaison des trois critères de qualité
A partir des données prédites par spectroscopie, le taux de C/N a été calculé pour chacune des modalités au stade initial.
Tableau 7. Valeurs de C/N pour les neufs modalités
Modalité BF41 P41 BF50 K41 K50 P50 P52 BF52 K52
C/N 66.5 74.5 78.6 78.6 81.0 86.9 88.3 88.6 111.0
Pour les trois critères, le 18-52 se différencie des deux autres clones par une meilleure qualité. La figure 22 montre les corrélations qui existent de façon globale entre les trois paramètres. Elle est de 65% entre v et qo et de 62% entre C:N et qo.
Les expériences sur la décomposition menées par Berg et Ekbohm (1983) montrent que au-delà d’une certaine valeur de C:N la teneur en azote décroît en fonction de la perte de masse. Pour les valeurs inférieures au C/N critique la teneur en azote augmente avec la perte de masse.
Les résultats obtenus ici ne coïncident pas avec cette conclusion : pour le 18-52 qui possède un taux de C/N élevé, la corrélation avec la masse restante est négative, pour les autres elle est positive. Ceci est probablement dû au fait que notre expérience de décomposition a été menée en microcosme et s’étale sur six semaines alors que l’expérience de Berg a été faite en milieu naturel sur un an.
Par ailleurs, on obtient des conclusions différentes de Berg et McClaugherty (2003) puisque dans notre cas un taux de C/N bas, donc un taux élevé de N (on considère constante la teneur en carbone entre les modalités) correspond à une faible décomposabilité. Cependant, dans l’étude de Berg et McClaugherty (2003), les corrélations positives observées entre la perte de masse au bout d’un an et la teneur en azote de la litière initiale ne sont significatives que pour un nombre élevé d’espèces : lorsqu'une seule espèce est considérée, la relation n’est pas significative (Table 6.2, p114, Berg et McClaugherty 2003).
Il faudrait pour conclure sur ce point faire une série d’analyses complémentaires des teneur d’azote sur les solutions de lessiva, sur les feuilles après lessivage et enfin sur les cendres.