Publisher’s version / Version de l'éditeur:
Biochemical Journal, 38, 2, pp. 168-178, 1944
READ THESE TERMS AND CONDITIONS CAREFULLY BEFORE USING THIS WEBSITE.
https://nrc-publications.canada.ca/eng/copyright
Vous avez des questions? Nous pouvons vous aider. Pour communiquer directement avec un auteur, consultez la
première page de la revue dans laquelle son article a été publié afin de trouver ses coordonnées. Si vous n’arrivez pas à les repérer, communiquez avec nous à [email protected].
Questions? Contact the NRC Publications Archive team at
[email protected]. If you wish to email the authors directly, please see the first page of the publication for their contact information.
NRC Publications Archive
Archives des publications du CNRC
This publication could be one of several versions: author’s original, accepted manuscript or the publisher’s version. / La version de cette publication peut être l’une des suivantes : la version prépublication de l’auteur, la version acceptée du manuscrit ou la version de l’éditeur.
Access and use of this website and the material on it are subject to the Terms and Conditions set forth at
The nature of the Aeromonas fermentation
Stanier, R. Y.; Adams, G. A.
https://publications-cnrc.canada.ca/fra/droits
L’accès à ce site Web et l’utilisation de son contenu sont assujettis aux conditions présentées dans le site LISEZ CES CONDITIONS ATTENTIVEMENT AVANT D’UTILISER CE SITE WEB.
NRC Publications Record / Notice d'Archives des publications de CNRC:
https://nrc-publications.canada.ca/eng/view/object/?id=07d783b9-f2ee-415d-bcc7-91b2b7d0d23d https://publications-cnrc.canada.ca/fra/voir/objet/?id=07d783b9-f2ee-415d-bcc7-91b2b7d0d23d
I944
The
Nature of the Aeromonas Fermentation
BY R. Y. STANIER AND G. A. ADAMS, Divisionof AppliedBiology, National Re8earch Council, Ottawa,Canada
(Received 26 February 1944) The genus Aeromona8 was establishedby Kluyver
& van Niel (1936) for pseudomonads fermenting carbohydrateswith the production of carbon dioxide and hydrogen. Since von Wolzogen Kiihr (1932)
hadpreviously shown thataxnemberofthisgroup, Pseudomonas fermentan8, gave a positive Voges-Proskauer reaction, the existence of a butylene glycol fermentation in the genus was suspected, although the absence ofexactdataontheproducts offermentation left this point unsettled.
During the course ofan intensive investigation
on the bacteriological production of 2:3-butylene
glycol (Adams&Stanier, 1944),it seemed desirable
tomakeasomewhat closer study of this little-known Aeromona8 fermentatior.. The recent discovery of polar flagellation in P.ateu8 hydrophilus (Kulp & Borden, 1942; Guthrie& Hitchner, 1943), taken in
conjunction with its fermentative abilities, mark this organism as an Aeromonas species, and since cultures werereadily available it waschosen as a representative of thegroup.
MATERIALS AND METHODS
Through the kindness ofDr WV. L. Kulp and Dr E. R.
Hitchner we obtained a number ofstrains ofAeromo8as
hydro-phila. Preliminary analyses having shown that all
these organisms were capable of producing substantial
amountsof ethanol and 2:3-butylene glycol fromsugars, a
strain with particularly vigorous fermentative properties
wasselected formoredetailedquantitative investigations.
The basal medium employed consisted ofyeast extract
1-25g.,CaCO,1-0g.,
MgSO4
0-025g., -OM-phosphate buffer(pH 7.4) 0-65ml., tap water 250ml. The carbohydrates used were sterilized separately in concentrated aqueous
solutionand added tothe basalmedium just before
inocu-lation in amounts givinga final concentration of 1-2 %.
Fermentationswere carriedout at30° and allowedtorun
for3-4days. During this periodaslowstreamof oxygen-freenitrogenwaspassed through the flasks. At the end of
thefermentation, an excess of HCIwasaddedtodissolve
the residual
CaCO3,
and aeration with nitrogen wascon-tinued for several hoursto sweepoutall theCO..
Carbon dioxidewasdeterminedgravimetrically. Hydro-gen was continuously oxidized by passage over heated
copperoxide anddeterminedaswater. Aftercompletion of
the fermentation, the fermented solution wasneutralized
and made up to500 ml. A measured fraction was made
alkalineand extracted continuously withetherin orderto
removebutyleneglycol and acetoin.The ether extractwas
evaporated, andtheresidue takenupin water. Butylene
glycol wasdeterminedby anunpublished method developed in the Department of Biochemistry at the University of Wisconsin. The butylene glycol is oxidized with periodic
acid, and the excess periodatedeterminediodometrically in neutral solution. Acetoin wasestimated by the method of Langlykke & Peterson (1937). After extraction of the alkaline solution with ether, it was made acid and re-extracted with ether to remove the organic acids, which were taken up in water again afterremoval ofthe ether. The acid extract was steamdistilledandthevolatileportion estimated as acetic acid by titration. The non-volatile fraction contained lactic and succinic acids. The formerwas
determined by the method of Boyland (1928), the latterby the formation of the silver salt. Pyruvicacid,when present, was withheld from going into the ether acid extract by saturating the solution with sodium bisulphite. Ethanol was determined by dichromate oxidation, following frac-tional distillationof a measured fraction ofthe fermentation solution.
Pyruvic acid was determined manometrically by ceric sulphate oxidation(Krebs & Johnson, 1937). Xylome esti-mations were carried out by the copper reduction method ofSchoorl (1929). Glucose was determined by the Hanes (1929) modification of theHagedorn-Jensen procedure.
RESULTS
Carbon balances were obtained for the fermenta-tions of glucose, xylose and pyruvic acid. An attempt was also made to study the fermentation of mannitol but, largely owing to the slow and
incompleteattack on thiscompound,asatisfactory
balance was not obtained.
Fermentaion ofglucose. Theproductsofglucose fermentationbyA.hydroph.ilaare shown in Table1. The mainorganic ones arealways butylene glycol,
ethanol and lacticacid;inaddition,smaller amounts ofacetoin, acetic acid and succinic acid are found. Fluctuations in the quantitative relationships be-tween the various products may occur from one
experiment to the next; thus, C02 production can vary from 140 to 170 mol. per 100 mol. ofglucose fermented,with concomitant differences in the rela-tive amounts of
C.
andC3 compounds.The ratio ofbutylene glycol to ethanol can also fluctuate over
quite a wide range, although it is usually about 1: 1 onamolecular basis.
In order toestablishbeyond doubtthenatureof the glycol produced in this fermentation, a small samplewasisolatedandcharacterized. About151. ofafermented yeast extract-glucose mediumwere
AEROMONAS FERMENTATION
Table 1. Fermentationofglucose by Aeromonas hydrophila Glucose fermented: 5.490 g.
Number of mol./100 mol. of glucose fermented
Products 2:3-Butylene glycol Acetoin Ethanol Acetic acid Lacticacid Succinic acid CO2
Hs
Total Weight (g.) 1-502 0-047 0-730 0-085 0-639 0-130 2-230 0-035 Product 54-7 1-7 52-0 4-6 23-3 3-6 166-257.5
Carbon 218-8 6-8 104-0 92 69-9 14-4 166-2 589-3 CalculatedCo2
109-4 3.4 52-04-6
-3-6166-8'
Calculated H2 54.7 3.4 9-2 -3-6 63-7 Oxidation values +_ 164-1 - 3-4 - 104-0 3-6 332-4 336-057.7
329-2 Carbonrecovery ... ... ...CO2
calculated/CO2
observed ...98-2% 0-997
evaporated to a volume of 41. in an open steam
kettle, saturated with anhydrous
K2CO3,
andex-tractedwith twice the volume ofbutanol. Most of the butanol was removedby distillation througha shortVigreux column,theresidualextract was dis-tilled under reduced pressure in a Stedmancolumn
and the pure glycol fraction recovered. Oxidation
of a small sample ofthis glycolwith bromine and
FeCl3
gave a good yield of diacetyl, which wascharacterizedbythe
melting-point
of itsbis-phenylhydrazone. Theproductionofdiacetylonoxidation
provedconclusivelythat theglycolwas2:3-butylene
glycol. Judgingfrom its opticalrotation (-
0.970),
it consists of a mixture of the 1- and meso-forms with a
prepo'
lerance of the latter. Thus it differsbothfrom tho 2:3-butylene glycol producedin the
Aerobacter fermentation,whichis a d-mesomixture,
and fromthatproduced in theAerobacillus fermen-tation, which is the pure1-isomer.
Fermentation of xylose. Table 2 presents the
carbon balance for a xylose fermentation by A. hydrophila. In this particular experiment the
butylene glycol-ethanol ratio was somewhat less
H2
calculated/H2
observed ... 1-108O/R index ... ... ... 1-021
than 1:1, but this is not characteristic; ratios of 1:1 or over have been obtained in other similar
experiments. Thus there are no significant differ-ences, either qualitative or quantitative, between theend-products ofpentose and hexose fermenta-tionbyA.hydrophila. This indicates very strongly that there must be a resynthesis from the xylose (or from the C2 fragments of the xylose) of a
C,
compoundwhich is subsequentlydecomposed in a manner similar to that operative in glucose
dis-similation. Unless one is willing to assume the production ofCO2from a
C2
compound-r-a reaction unknowninfermentative processes-a simplecalcu-lationshows the necessity for this assumption. Were the end-products ofpentose fermentation derived
directlyfrom the fragmentsofa
C3-C2
split,theCO2
production could not exceed 100
mol./100
mol. of pentose fermented, since no CO2 couldbe derived fromtheC.
fraction. Furthermore, the productionof
C3
compoundswill reduce thispossiblemaximum ofCO2
productionby an amountequivalenttothe number of mols ofC3
compound formed. In theexperiment shown in Table 2, the observed
C02
Table2. Fermentation ofxylo8ebyAeromonashydrophila
Xylose fermented: 4-010 g.
Numberofmol./100 mol. of xylose fermented
Products 2:3-Butylene glycol Acetoin Ethanol Acetic acid Lacticacid Succinicacid CO2
H2
Total Weight(g.)
0-939 0-061 0-600 0-150 0-491 0-036 1-584 0-029 Product 39-0 2-6 48-9 9-3 20-4 1-1 134-7 53-9 96-6% 1-041 Carbon 156-0 10-4 97-8 18-6 61-2 4-4 134-7 Calculated CO2 78-0 5-2 48-9 9-3 -1-1 - 483-1 140-3 Calculated H2 39-0 5-2 18-6 -1-1 61-7 Oxidationvalues r 'A +_ 117-0 5-2 - 97-8 1.1 269-4 270-5 Carbonrecovery ...H,c ... ... C02calculated/CO,"observed ... 53-9 273-9 1-1260-988
Vol.
38
169
H2
calculated,/H2
obsorved ...O/R
index ... ... ...R.
Y. STANIER AND G. A. ADAMS
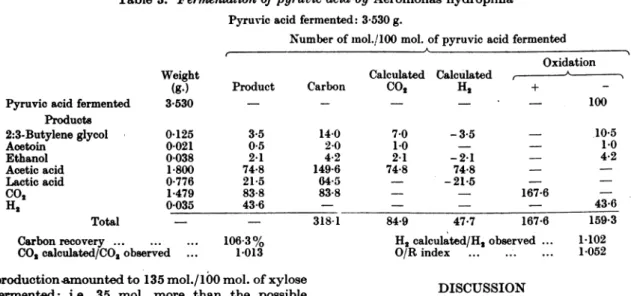
Table 3.
Fermentation
of pyruvic acid by Aeromonas hydrophilaPyruvicacid fermented:3-530g.
Numberof mol./100 mol. of pyruvic acid fermented
Pyruvicacidfermented Products 2:3-Butyleneglycol Acetoin Ethanol Aceticacid Lacticacid CO2 H2 Total Weight (g.) 3 530 0-125 0-021 0-038 1-800 0-776 1-479 0035 Carbonrecovery ... ...
CO2 calculated/CO2observed
Calculated Calculated Product Carbon CO H2 3-5 0-5 2-1 74-8 21-5 83-8 43-6 14-0 2-0 4-2 149-6 04-5 83-8 318-1 .. . 106-3% .... 1013 7-0 1-0 2-1 74-8 84*9 -3.5 -2-1 74-8 -
21*5
47.7 Oxidation + A 100 167-6 167-6 H2calculated/H2observed ... O/R index ... ... .production-amountedto135 mol./100 mol. of xylose fermented: i.e. 35 mol. more than the possible maximumfroma
C3-C2
split and58mol.morethan theamounttobeexpected after taking intoaccount the lactic and succinic acid formed. It would be interesting to determine the effect of A.hydrophila on glycol aldehyde, the C2 compound which one mightexpect to result from the primary fission of the pentose molecule.Fermentation ofpyruvic acid. Aeromona8
hydro-phila causes a vigorous fermentation of pyruvic
acid. The balance sheet for this fermentation is shown in Table 3. Surprisingly enough, the pro-duction of butylene glycol, acetoin and ethanol is largely suppressed, and the chief organic end-productsareacetic and lactic acids. Approximately
three molecules of pyruvic acid are oxidized to acetic acid for every one reduced to lactic acid, andconsequentlyaconsiderable volume of hydrogen
isproduced. The three main reactionsappeartobe:
3CH3.CO.COOH+3H20 3CH COOH+3CO2+6H,
CH3.CO COOH+2H CH,. CHOH COOH,
4H 2H2.
Thus thedismutation of pyruvic acid by A. hy-drophila belongs to the type demonstrated for heterofermentative lactic acid bacteria (Nelson &
Werkman, 1936) and for staphylococci(Krebs,1937),
with thedifferencethatin the latterfermentations there is an equimolecular production of the two
acids and consequentlynohydrogen formation. Thedissimilation of pyruvic acid by A.hydrophila differs notably from that whichoccursin theother two bacterial groups where one finds a butylene glycol fermentation. When growing anaerobically on pyruvic acid, AerobaciUus polymyxa produces principally acetoin and acetic acid(Adams &Stanier, 1944), while under similar conditions Aerobacter
aerogene8 gives rise to butylene glyocl and acetic
aoid(Mickelson, Reynolds & Werkman, 1936).
DISCUSSION
Theseinvestigationshavedefinitely established the existence of a butylene glycol fermentation in a
bacterialgroupwhere itwaspreviouslynot known. Thus there are at present three morphologically
widely separated bacterial families-the Entero-bacteriaceae (Aerobacter andSerratia), the
Bacilla-ceae (Aerobacillue) and the Pseudomonadaceae
(Aeromona8) in whichabutyleneglycolfermentation
occurs. In each family there are also organisms
which carry out entirely different carbohydrate fermentations; in the Pseudomonadaceae, the alco-holic fermentation of Zymomona lindneri: in the Enterobacteriaceae, the mixed acid fermentationof Escherichia coli and other species: and in the
Bacillaceaeawidevariety of dissimilatoryprocesses
(acetone-butanol, acetone-ethanol and other fer-mentations)performed by various organisms.
This absence of correlation between morpho-logical and biochemical characters isof considerable interest from the standpoint of comparative bio-chemistry. Itsuggestsverystronglythat thepresent fermentative mechanisms for the dissimilation of
carbohydrates have developed independently along similar lines in different branches ofthe bacteria.
Thishypothesis is supported by the slightbut signi-ficant differences which characterize a particular fermentationascarriedoutbyorganismsbelonging todifferent morphologicalgroups. Agood example is the differencesinoptical rotationofthebutylene
glycols produced by Aerobacter, Aeromonas and
Aerobacillus.
SUMMARY
1. Aeromona hydrophila,atypicalrepresentative
of the genus Aeromona. Kluyver and van Niel, causes a butylene glycol ferinentation of carbo-hydrates. The end-products from glucose are
2:3-10.5 1-0 4-2 43-6 159-3 1-102 1-052
1944
Vol. 38
AEROMONAS FERMENTATION
171
butylene glycol, ethanol, acetic and lactic acids,
CO2
and H,, with traces of acetoin and succinic acid. The 2:3-butylene glycol is a mixture of the 1- and meso-forms.2. The products of xylosefermentation arevery similar, both quantitatively and qualitatively, to those of glucose fermentation. This is regarded as
evidenceagainsta simple
C3-C2
splitof the pentosemolecule.
3. The main products of the fermentation of
pyruvic acidare lacticandacetic acids, CO2and
H2.
Approximatelythree molecules ofpyruvicacid are oxidized to acetic acid for every one reduced to lactic acid.
REFERENCES Adams, G. A. & Stanier, R. Y. (1944). Canad. J. Res.
(inthePress).
Boyland, E. (1928). Biochem. J. 22, 236.
Guthrie, R.& Hitchner, E.R.(1943). J.Bact.46,52. Hanes, C. S.(1929). Biochem. J.28,99.
Kluyver,A. J. & van Niel,C. B. (1936). Zbl. Bakt. ia,94,369. Krebs, H. A. (1937). Biochem. J. 31, 661.
-& Johnson,W. A. (1937). Biochem. J. 31, 645. Kulp, W. L. & Borden, D. G.(1942). J.Bact.44, 673.
Langlykke, A. & Peterson, W. H. (1937). Industr. Engng Chem.(Anal. ed.), 9, 163.
Mickelson, M., Reynolds, H. & Werkman, C. H. (1936). Proc.Soc. exp. Biol., N. Y., 34, 748.
Nelson, M. E. & Werkman, C. H. (1936). Iowa St. Coll. J. Sci. 10, 141.
Schoorl,N. (1929). Chem. Weekbl.26,130.
von Wolzogen Kuhr, C. A. H. (1932). Zbl. Bakt. II, 85, 223.
Experiments on the Methylation and Acetylation of
Wool,
Silk
Fibroin,
Collagen
and
Gelatin
BY S. BLACKBURN AND H. PHILLIPS, The Wool
Indu8trie
ResearchA88ociation, Torridon, Leedi (Received 3 March 1944)In apreviouspaper (Blackburn, Carter& Phillips, 1941) it was shown that woolandsilkfibroincould be O-methylated either by contact with methyl
bromide or iodide at room temperatures, or with methyl sulphate in buffers within the pH range
2-5-8-5. With both proteins the number of
CH3
groups introduced by repeated treatment with
methylsulphatewas greater than couldbe accom-modated on the free carboxyl groups of glutamic
andaspartic acids, andit wassuggestedthatcertain 'activated' peptide linkages methylatedas well as thefreecarboxylgroups of thesalt-linkages. CH3I and
CH3Br
introducedfewerCOH groupsinto wool and silk fibroin thandidmethyl sulphate.Thepresent paperdescribes the extension of this
work to themethylationof collagen and gelatinby similar methods,and themethylation with methyl sulphate andCH8Brof woolwhich has bieen chemi-cally modified by alkali, acid,
nitrous
acid andformaldehyde. In addition, the
siaultaneous
ace-tylation and methylation of wool, silk fibroin and collagen by the combined action ofmethanol and
acetic anhydride has been investigated.
METHODS
Methylation of collagenandgelatin with methyl
suphate
andmethyl halidesThecollagenused in theseinvestigations was a limed hide which had been delimed with acetic acid and then de-hydratedinacetone. The gelatin wasCoignet'sGoldLabel.
The methylation procedure wassimilarto thatdescribed for wool and silk (Blackburn et al. 1941), with certain modifications to suit the propertiesof collagen and gelatin. Both proteins were cut intosmallpiecesand acetate buffer was used for the methylations with methyl sulphate. At theend ofeachperiod ofmethylationabout1 g. ofcollagen was removed, washed in three changes of water and then shaken mechanically with water(25ml.) for 1 hr. It was thendehydrated with acetone,air-driedand conditioned.* The moisture content of the collagen, the physical pro-perties ofwhich appeared to beunaltered,wasdetermined by Barritt & King's method (1926). During methylation, the gelatin absorbed water and swelled greatly. The re-sulting gel and solution was dialyzed incellophan against waterfor 3 days,thewaterbeingchangedfrequently.The resultinggelatinousmasswasdried in vacuobelow 60°.
Before methylation with either CH3Br or CH3I, the collagen was allowed to reach equilibrium with a borate buffer of pH 8, and then dehydrated with acetone and conditioned. The gelatin was treated dry asbought. After methylation, both proteinswere rinsedinseveralchanges of benzene and then conditioned. When proteins are methylated byeither
CHsBr
orCH3I,
onehalogenanion is liberated for each CH3 group introduced. To provide an additional measure of their degree of methylation, the halogen contentof some of themethylated collagens and gelatins was therefore determined by the open Carius method(Peters&VanSlyke, 1932). The methylated gelatin* Allproteins described in this paper as conditioned had
beenexposed to a controlled atmosphere of 210 and 70% R.H. until they attained a constant weight. Although all the analyses were made on the conditioned proteins, the results are recorded as percentages of the anhydrous material.